
Capripoxvirus Infections in Ruminants: A Review

 ,
,
Abstract
1. Introduction
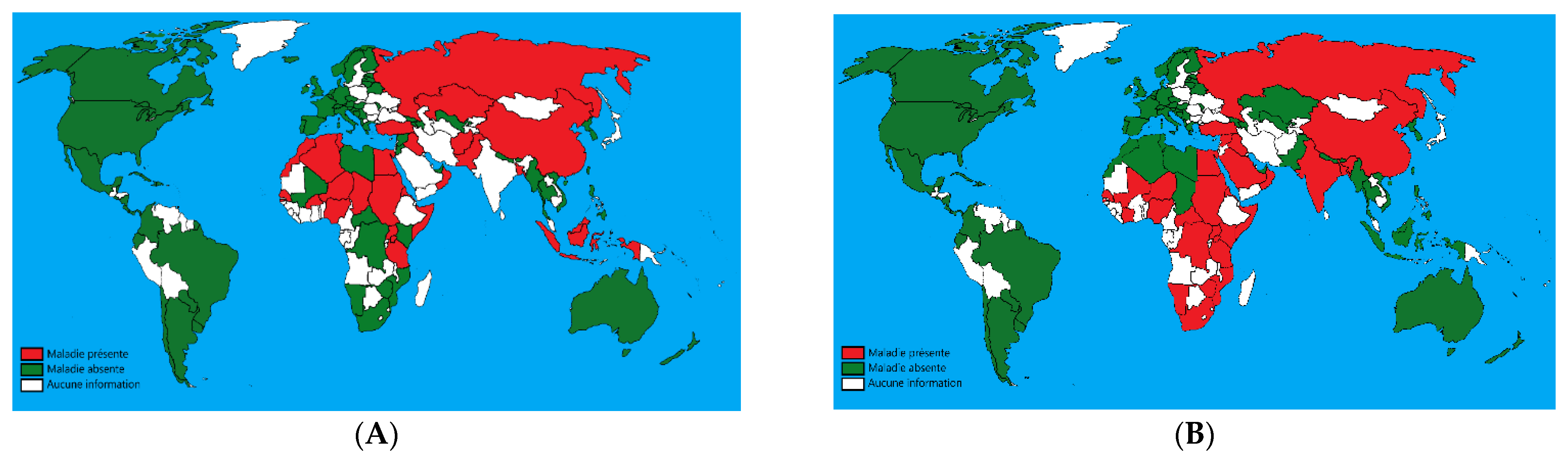
2. Importance
3. Aetiology
4. Clinical Signs
5. Immunity
6. Vaccination against Capripoxviruses
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- OIE. Sheep Pox and Goat Pox. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals. Chapter 3.7.12. 2018. Available online: https://www.oie.int/fileadmin/Home/eng/Animal_Health_in_the_World/docs/pdf/Disease_cards/SHEEP_GOAT_POX.pdf (accessed on 10 February 2021).
- Babiuk, S.; Bowden, T.R.; Boyle, D.B.; Wallace, D.B.; Kitching, R.P. Capripoxviruses: An Emerging Worldwide Threat to Sheep, Goats and Cattle. Transbound Emerg. Dis. 2008, 55, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.S.M.; Venter, E.H.; Shisler, J.L.; Gari, G.; Mekonnen, G.A.; Juleff, N.; Lyons, N.A.; De Clercq, K.; Upton, C.; Bowden, T.R.; et al. Review: Capripoxvirus Diseases: Current Status and Opportunities for Control. Trans. Emerg. Dis. 2017, 64, 729–745. [Google Scholar] [CrossRef] [PubMed]
- World Animal Health Information Database (WAHIS). 2020. Available online: https://www.oie.int/wahis_2/public/wahid.php/Diseaseinformation/Diseasetimelines (accessed on 26 October 2020).
- Sprygin, A.; Pestova, Y.; Wallace, D.B.; Tuppurainen, E.; Kononov, A.V. Transmission of Lumpy Skin Disease virus: A short review. Virus Res. 2019, 269, 197637. [Google Scholar] [CrossRef] [PubMed]
- Buller, R.M.; Arif, B.M.; Black, D.N.; Dumbell, K.R.; Esposito, J.J.; Lefkowitz, E.J.; McFad–den, G.; Moss, B.; Mercer, A.A.; Moyer, R.W.; et al. Family Poxviridae. In Virus Taxonomy: Classification and Nomenclature of Viruses. Eighth Report of the International Committee on Taxonomy of Viruses; Fauquet, C.M., Mayo, M.A., Maniloff, J., Desselberger, U., Ball, L.A., Eds.; Elsevier: Amsterdam, The Netherlands, 2005; pp. 117–133. [Google Scholar]
- Kitching, R.P.; Mellor, P.S. Insect transmission of capripox virus. Res. Vet. Sci. 1988, 40, 255–258. [Google Scholar] [CrossRef]
- Bowden, T.R.; Babiuk, S.L.; Parkyn, G.R.; Copps, J.S.; Boyle, D.B. Capripoxvirus tissue tropism and shedding: A quantitative study in experimentally infected sheep and goats. Virology 2008, 371, 380–393. [Google Scholar] [CrossRef] [PubMed]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Mechanical transmission of Lumpy Skin Disease virus by Aedes aegypti (Diptera: Culicidae). Epidemiol. Infect. 2001, 126, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Chihota, C.M.; Rennie, L.F.; Kitching, R.P.; Mellor, P.S. Attempted mechanical transmission of Lumpy Skin Disease virus by biting insects. Med. Vet. Entomol. 2003, 17, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.S.M.; Stoltsz, W.H.; Troskie, M.; Wallace, D.B.; Oura, C.A.L.; Mellor, P.S.; Coetzer, J.A.W.; Venter, E.H. A potential role for ixodid (hard) tick vectors in the transmission of Lumpy Skin Disease virus in cattle. Transbound. Emerg. Dis. 2011, 58, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.S.; Lubinga, J.C.; Wilhelm, J.C.; Stoltsz, H.; Troskie, M.; Carpenter, S.T.; Coetzer, J.A.W.; Venter, E.H.; Oura, A.L. Mechanical transmission of Lumpy Skin Disease virus by Rhipicephalus appendiculatus male ticks. Epidemiol. Infect. 2013, 141, 425–430. [Google Scholar] [CrossRef]
- Lubinga, J.C.; Tuppurainen, E.S.; Coetzer, J.A.; Stoltsz, W.H.; Venter, E.H. Transovarial passage and transmission of LSDV by Amblyomma hebraeum, Rhipicephalus appendiculatus and Rhipicephalus decoloratus. Exp. Appl. Acarol. 2014, 62, 67–75. [Google Scholar] [CrossRef]
- Sohier, C.; Haegeman, A.; Mostin, L.; De Leeuw, I.; Van Campe, W.; De Vleeschauwer, A.; Tuppurainen, E.; van den Berg, T.; De Regge, N.; De Clercq, K. Experimental evidence of mechanical Lumpy Skin Disease virus transmission by Stomoxys calcitrans biting flies and Haematopota spp. horseflies. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Babiuk, S.; Bowden, T.R.; Parkyn, G.; Dalman, B.; Hoa, D.M.; Long, N.T.; Vu, P.P.; Bieu, D.X.; Copps, J.; Boyle, D.B. Yemen and Vietnam capripoxviruses demonstrate a distinct host preference for goats compared with sheep. J. Gen. Virol. 2009, 90, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Kreindel, S.; Masiulis, M.; Skrypnyk, A.; Zdravkova, A.; Escher, M.; Raizman, E. Emergence of lumpy skin disease in Asia and Europe. Empres Anim. Health 2016, 46, 360. [Google Scholar]
- Hurisa, T.T.; Jing, Z.; Jia, H.; Chen, G.; He, X.B. A Review on Sheeppox and Goatpox: Insight of Epidemiology, Diagnosis, Treatment and Control Measures in Ethiopia. J. Infect. Dis. Epidemiol. 2018, 4, 057. [Google Scholar] [CrossRef]
- Tuppurainen, E.S.; Galon, N. Lumpy Skin Disease: Current situation in Europe and neighbouring regions and necessary control measures to halt the spread in South-East Europe. Oie Reg. Comm. 2016. [Google Scholar]
- Yeruham, I.; Nir, O.; Braverman, Y.; Davidson, M.; Grinstein, H.; Haymovitch, M.; Zamir, O. Spread of Lumpy Skin Disease in Israeli dairy herds. Vet. Rec 1995, 137, 91–93. [Google Scholar] [CrossRef] [PubMed]
- Ayelet, G.; Abate, Y.; Sisay, T.; Nigussie, H.; Gelaye, E.; Jemberie, S.; Asmare, K. Lumpy Skin Disease: Preliminary vaccine efficacy assessment and overview on outbreak impact in dairy cattle at debre zeit, central Ethiopia. Antivir. Res. 2013, 98, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Ben-Gera, J.; Klement, E.; Khinich, E.; Stram, Y.; Shpigel, N.Y. Comparison of the efficacy of Neethling Lumpy Skin Disease virus and x10RM65 sheep-pox live attenuated vaccines for the prevention of Lumpy Skin Disease–The results of a randomized controlled field study. Vaccine 2015, 33, 4837–4842. [Google Scholar] [CrossRef] [PubMed]
- Boumart, Z.; Daouam, S.; Belkourati, I.; Rafi, L.; Tuppurainen, E.; Tadlaoui, K.O.; El Harrak, M. Comparative innocuity and efficacy of live and inactivated sheeppox vaccines. BMC Vet. Res. 2016, 12, 133. [Google Scholar] [CrossRef]
- Hamdi, J.; Boumart, Z.; Daouam, S.; El Arkam, A.; Bamouh, Z.; Jazouli, M.; Tadlaoui, O.K.; Fassi Fihri, O.; Gavrilov, B.; El Harrak, M. Development and Evaluation of an Inactivated Lumpy Skin Disease Vaccine for Cattle. Vet. Microbiol 2020, 245, 108689. [Google Scholar] [CrossRef] [PubMed]
- Wolff, J.; Moritz, T.; Schlottau, K.; Hoffmann, D.; Beer, M.; Hoffmann, B. Development of a Safe and Highly Efficient Inactivated Vaccine Candidate against Lumpy Skin Disease Virus. Vaccines 2021, 9, 4. [Google Scholar] [CrossRef]
- Yeruham, I.; Yadin, H.; Van Ham, M.; Bumbarov, V.; Soham, A.; Perl, S. Economic and epidemiological aspects of an outbreak of sheeppox in a dairy sheep flock. Vet. Rec 2007, 160, 236–237. [Google Scholar] [CrossRef] [PubMed]
- Garner, M.G.; Sawarkar, S.D.; Brett, E.K.; Edwards, J.R.; Kulkarni, V.B.; Boyle, D.B.; Singh, S.N. The Extent and Impact of Sheep Pox and Goat Pox in the State of Maharashtra, India. Trop. Anim. Health Prod. 2000, 32, 205–223. [Google Scholar] [CrossRef] [PubMed]
- Tuppurainen, E.S.M.; Oura, C.A.L. Review: Lumpy Skin Disease: An Emerging Threat to Europe, the Middle East and Asia. Trans. Emerg. Dis. 2012, 9, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Gari, J.G. Epidemiological Study of Lumpy Skin Disease and Its Economic Impact in Ethiopia. Ph.D. Thesis, Institut National Polytechnique de Toulouse, Toulouse, France, 2011. [Google Scholar]
- Coetzer, J.A.W. Lumpy Skin Disease. In Infectious Diseases of Livestock; Coetzer, J.A.W., Tustin, R.C., Eds.; Oxford University Press: Oxford, UK, 2004; pp. 1268–1276. [Google Scholar]
- Peck, D.; Bruce, M. The economic efficiency and equity of government policies on brucellosis: Comparative insights from Albania and the United States of America. Rev. Sci. Tech. Off. Int. Epiz 2017, 36, 291–302. [Google Scholar] [CrossRef]
- Tuppurainen, E.; Babiuk, S.; Klement, E. Lumpy Skin Disease; Springer International Publishing AG: Berlin/Heidelberg, Germany, 2018. [Google Scholar]
- Alemayehu, G.; Zewde, G.; Admassu, B. Risk assessments of Lumpy Skin Disease in Borena bull market chain and its implication for livelihoods and international trade. Trop. Anim. Health Prod. 2013, 45, 1153–1159. [Google Scholar] [CrossRef] [PubMed]
- Hailu, B.; Alemayehu, G. Epidemiology, Economic Importance and Control Techniques of Lumpy Skin Disease: A review. InJAR 2015, 3, 197–205. [Google Scholar]
- Joshi, R.K.; Chandra, R.; Rao, V.D.P.; Garg, S.K. Goatpox: A Review. J. Appl. Anim. Res. 1992, 2, 57–72. [Google Scholar] [CrossRef][Green Version]
- Biswas, S.; Noyce, R.S.; Babiuk, L.A.; Lung, O.; Bulach, D.M.; Bowden, T.R.; Boyle, D.B.; Babiuk, B.; Evans, D.H. Extended sequencing of vaccine and wild-type capripoxvirus isolates provides insights into genes modulating virulence and host range. Transbound. Emerg. Dis. 2019, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Van Regenmortel, M.H.V.; Fauquet, C.M.; Bishop, D.H.L. Virus Taxonomy: Seventh Report of the International Committee on Taxonomy of Viruses; Academic Press: Jerusalem, Israel, 1996. [Google Scholar]
- Nagington, J. Electron Microscopy in Differential Diagnosis of Poxvirus Infections. Brit. Med. J. 1964, 2, 1499–1500. [Google Scholar] [CrossRef][Green Version]
- Vilagines, P.; Vilagines, R. Les poxviridés: Caractères généraux. In Virologie Médicale; Maurin, J., Ed.; Presses Universitaires Lyon: Lyon, France, 1985; pp. 310–319. [Google Scholar]
- Moss, B. Recombinant Poxviruses; CRC Press: Boca Raton, FL, USA, 1992; pp. 45–80. [Google Scholar]
- McLysaght, A.; Baldi, P.F.; Gaut, B.S. Extensive gene gain associated with adaptive evolution of poxviruses. Proc. Natl. Acad. Sci. USA 2003, 100, 15655–15660. [Google Scholar] [CrossRef]
- Hughes, A.L.; Friedman, R. Poxvirus genome evolution by gene gain and loss. Mol. Phylogenetics Evol. 2005, 35, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Bratke, K.A.; McLysaght, A. Identification of multiple independent horizontal gene transfers into poxviruses using a comparative genomics approach. BMC Evol. Biol. 2008, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Tarbouriech, O.; Flusin, C.; Sele, F.; Iseni, F. La synthèse du génome des poxvirus. Virologie 2012, 16, 210–224. [Google Scholar]
- Moss, B. Poxviridae: The viruses and their replication. In Fields Virology; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 2905–2946. [Google Scholar]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Kutish, G.F.; Rock, D.L. Genome of Lumpy Skin Disease Virus. J. Virol. 2001, 75, 7122–7130. [Google Scholar] [CrossRef]
- Tulman, E.R.; Afonso, C.L.; Lu, Z.; Zsak, L.; Sur, J.H.; Sandybaev, N.T.; Kerembekova, U.Z.; Zaitsev, V.L.; Kutish, G.F.; Rock, D.L. The genomes of sheepox and goatpox viruses. J. Virol. 2002, 76, 6054–6061. [Google Scholar] [CrossRef]
- Rouby, S.R. RPO30 Gene based PCR for Detection and Differentiation of Lumpy Skin Disease Virus and Sheep Poxvirus Field and Vaccinal Strains. Vet. Sci. Res. Rev. 2018, 4, 1–8. [Google Scholar] [CrossRef]
- Chibssa, T.R.; Settypalli, T.B.K.; Berguido, F.J.; Grabherr, R.; Loitsch, A.; Tuppurainen, E.; Nwankpa, N.; Tounkara, K.; Madani, H.; Omani, A.; et al. An HRM Assay to Differentiate Sheepox Virus Vaccine Strains from Sheepox Virus Field Isolates and other Capripoxvirus Species. Sci. Rep. 2019, 9, 6646. [Google Scholar] [CrossRef]
- Lamien, C.E.; Lelenta, M.; Goger, W.; Silber, R.; Tuppurainen, E.; Matijevic, M.; Luckins, A.G.; Diallo, A. Real time PCR method for simultaneous detection, quantitation and differentiation of Capripoxviruses. J. Virol. Methods 2011, 171, 134–140. [Google Scholar] [CrossRef]
- Le Goff, C.; Lamien, C.E.; Fakhfakh, E.; Chadeyras, A.; Aba-Adulugba, E.; Libeau, G.; Tuppurainen, E.; Wallace, D.B.; Adam, T.; Silber, R.; et al. Capripoxvirus G-protein-coupled chemokine receptor: A host-range gene suitable for virus animal origin discrimination. J. Gen. Virol. 2009, 90, 1967–1977. [Google Scholar] [CrossRef]
- Diallo, A.; Viljoen, G.J. Genus Capripoxvirus. In Poxviruses; Mercer, A.A., Schmidt, A., Weber, O., Eds.; Birkhäuser: Basel, Switzerland, 2007; pp. 167–181. [Google Scholar]
- Hamdi, J.; Bamouh, Z.; Jazouli, M.; Alhyane, M.; Safini, N.; Tadlaoui, K.O.; Fassi Fihri, O.; El Harrak, M. Experimental infection of indigenous North African goats with goatpox virus. Acta Vet. Scand 2021, 63, 9. [Google Scholar] [CrossRef] [PubMed]
- CFSPH, Iowa State University. Sheep & Goat Pox. 2008. Available online: http://www.cfsph.iastate.edu/Factsheets/pdfs/sheep_and_goat_pox.pdf (accessed on 19 February 2021).
- Barnard, B.J.; Munz, E.; Dumbell, K.; Prozesky, L. Lumpy Skin Disease. In Infectious Diseases of Livestock with Special Reference to Southern Africa; Coetzer, J.A.W., Thomson, G.R., Tustin, R.C., Eds.; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Wolff, J.; Abd El Rahman, S.; King, J.; El-Beskawy, M.; Pohlmann, M.; Beer, M.; Hoffmann, B. Establishment of a Challenge Model for Sheeppox Virus Infection. Microorganisms 2020, 8, 2001. [Google Scholar] [CrossRef] [PubMed]
- Gari, G.; Abie, G.; Gizaw, D.; Wubete, A.; Kidane, M.; Asgedom, H.; Tuppurainen, E.S.M. Evaluation of the safety, immunoge- nicity and efficacy of three capripoxvirus vaccine strains against Lumpy Skin Disease virus. Vaccine 2015, 33, 3256–3261. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, J.; Bamouh, Z.; Jazouli, M.; Boumart, Z.; Omari Tadlaoui, K.; Fassi Fihri, O.; El Harrak, M. Experimental evaluation of the cross-protection between Sheepox and bovine Lumpy skin vaccines. Sci. Rep. 2020, 10, 8888. [Google Scholar] [CrossRef]
- Arsevska, E.; Bronner, A.; Calavas, D.; Cauchard, J.; Caufour, P.; Falala, S.; Hamon, M.; Hendrikx, P.l.; Lancelot, R.; Mercier, A.; et al. Dermatose nodulaire contagieuse des bovins: État des connaissances et situation épidémiologique dans les Balkans au 31 juillet 2016. Bull. Epid St. Anim Alim 2016, 75, 20–24. [Google Scholar]
- Omar, R. Comparison of the Two Lumpy Skin Disease Virus Vaccines, Neethling and Herbivac, and Construction of a Recombinant Herbivac-Rift Valley Fever Virus Vaccine. Master’s Thesis, University of Cape Town, Cape Town, South Africa, 2015. [Google Scholar]
- Smith, S.A.; Kotwal, G.J. Immune Response to Poxvirus Infections in Various Animals. Crit. Rev. Microbiol. 2002, 28, 149–185. [Google Scholar] [CrossRef]
- Aristizábal, B.; González, A. Innate immune system. In Autoimmunity: From Bench to Bedside; Anaya, J.M., Shoenfeld, Y., Rojas-Villarraga, A., Eds.; El Rosario University Press: Bogota, Colombia, 2013. [Google Scholar]
- Moutaftsi, M.; Tscharke, D.C.; Vaughan, K.; Koelle, D.M.; Stern, L.; Calvo-Calle, M.; Ennis, F.; Terajima, M.; Sutter, G.; Crotty, S.; et al. Uncovering the interplay between CD8, CD4 and antibody responses to complex pathogens. Future Microbiol. 2010, 5, 221–239. [Google Scholar] [CrossRef]
- Bonilla, F.A.; Oettgen, H.C. Adaptative immunity. J. Allergy Clin. Immunol. 2010, 125, 33–40. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.S.; Warrington, R.; Watson, W.; Kim, H.L. An introduction to immunology and immunopathology. Allergy Asthma Clin. Immunol. 2018, 14, 49. [Google Scholar] [CrossRef]
- Milovanović, M.; Dietze, K.; Milićević, V.; Radojičić, S.; Valčić, M.; Moritz, T.; Hoffman, B. Humoral immune response to repeated lumpy skin disease virus vaccination and performance of serological tests. BMC Vet. Res. 2019, 15, 80. [Google Scholar] [CrossRef]
- Samojlović, M.; Polacek, V.; Gurjanov, V.; Lupulovic, D.; Lazic, G.; Petrovic, T.; Lazic, S. Detection of antibodies against Lumpy skin disease virus by Virus neutraliza–tion test and ELISA methods. Acta Vet. Beogr. 2019, 69, 47–60. [Google Scholar] [CrossRef]
- Kitching, R.P.; Bhat, P.P.; Black, D.N. The characterization of African strains of capripoxvirus. Epidemiol. Infect. 1989, 102, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Carn, V.M. Control of capripoxvirus infections. Vaccine 1993, 11, 1275–1279. [Google Scholar] [CrossRef]
- Seet, B.T.; Johnston, J.B.; Brunetti, C.R.; Barrett, J.W.; Everett, H.; Cameron, C.; Sypula, J.; Nazarian, S.H.; Lucas, A.; McFadden, G. Poxviruses and immune evasion. Annu. Rev. Immunol. 2003, 21, 377–423. [Google Scholar] [CrossRef]
- Norian, R.; Ahangaran, N.A.; Varshovi, H.R.; Azadmehr, A. Evaluation of humoral and cell-mediated immunity of two capripoxvirus vaccine strains against Lumpy Skin Disease virus. Iran. J. Virol. 2016, 10, 1–11. [Google Scholar] [CrossRef]
- Varshovi, H.R.; Norian, R.; Azadmehr, A.; Afzal Ahangaran, N. Immune response characteristics of Capri pox virus vaccines following emergency vaccination of cattle against Lumpy Skin Disease virus. IJVST 2017, 9, 33–40. [Google Scholar]
- Alberts, B. T Cells and MHC Proteins. In Molecular Biology of the Cell, 4th ed.; Garland Science: New York, NY, USA, 2002. [Google Scholar]
- Bray, M.; Wright, M.E. Progressive vaccinia. Clin Infect. Dis. 2003, 36, 766–774. [Google Scholar] [CrossRef]
- Belyakov, I.M.; Earl, P. Shared modes of protection against poxvirus infection by attenuated and conventional smallpox vaccine viruses. Proc. Natl. Acad. Sci. USA 2003, 100, 9458–9463. [Google Scholar] [CrossRef]
- Xu, R.; Johnson, A. Cellular and humoral immunity against vaccinia virus infection of mice. J. Immunol. 2004, 172, 6265. [Google Scholar] [CrossRef]
- Fang, M.; Sigal, L. Antibodies and CD8+ T cells are complementary and essential for natural resistance to a highly lethal cytopathic virus. J. Immunol. 2005, 175, 6829. [Google Scholar] [CrossRef]
- Edghill-Smith, Y.; Bray, M. Smallpox vaccine does not protect macaques with AIDS from a lethal monkeypox virus challenge. J. Infect. Dis. 2005, 191, 372–381. [Google Scholar] [CrossRef]
- Fenner, F.; Anderson, D.A.; Arita, I.; Jezek, Z.; Ladnyi, I.D. Smallpox and Its Eradication; World Health Organization: Geneva, Switzerland, 1988. [Google Scholar]
- Redfield, R.R.; Wright, D.C. Disseminated vaccinia in a military recruit with human immunodeficiency virus (HIV) disease. N Engl. J. Med. 1987, 316, 673–676. [Google Scholar] [CrossRef] [PubMed]
- Panchanathan, V.; Chaudhri, G.; Karupiah, G. Correlates of protective immunity in poxvirus infection: Where does antibody stand? Immunol. Cell Biol. 2008, 86, 80–86. [Google Scholar] [CrossRef]
- Kennedy, J.S.; Frey, S.E.; Yan, L.; Rothman, A.L.; Cruz, J.; Newman, F.K.; Orphin, L.; Belshe, R.B.; Ennis, F.A. Induction of Human T Cell–Mediated Immune Responses after Primary and Secondary Smallpox Vaccination. JID 2004, 190, 1286–1294. [Google Scholar] [CrossRef]
- Hammarlund, E.; Lewis, M.W.; Hansen, S.G. Duration of antiviral immunity after smallpox vaccination. Nat. Med. 2003, 9, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, S.M.; Pan, S.C.; Chen, S.Y.; Huang, P.F.; Chang, S.C. Age distribution for T cell reactivity to vaccinia virus in a healthy population. Clin. Infect. Dis. 2004, 38, 86–89. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S.; Felgner, P.; Davies, H.; Glidewell, J.; Villarreal, L.; Ahmed, R. Cutting edge: Long-term B cell memory in humans after smallpox vaccination. J. Immunol. 2003, 171, 4969–4973. [Google Scholar] [CrossRef] [PubMed]
- Putz, M.M.; Alberini, I.; Midgley, C.M.; Manini, I.; Montomoli, E.; Smith, G.L. Prevalence of antibodies to Vaccinia virus after smallpox vaccination in Italy. J. Gen. Virol. 2006, 86, 2955–2960. [Google Scholar] [CrossRef] [PubMed]
- Agianniotaki, E.I.; Babiuk, S.; Katsoulos, P.D.; Chaintoutis, S.C.; Praxitelous, A.; Quizon, K.; Boscos, C.; Polizopoulou, Z.S.; Chondrokouki, E.D.; Dovas, C.I. Colostrum transfer of neutralizing antibodies against lumpy skin disease virus from vaccinated cows to their calves. Transbound. Emerg. Dis. 2018, 65, 2043–2048. [Google Scholar] [CrossRef] [PubMed]
- Weiss, K.E. Lumpy Skin Disease. Virol. Monogr. 1968, 3, 111–131. [Google Scholar]
- Precausta, P.; Kota, F.; Vellut, G. A new freeze-dried living virus vaccine against sheep pox. Comp. Immunol. Microbiol. Infect. Dis. 1979, 1, 305–319. [Google Scholar] [CrossRef]
- Gitao, C.G.; Mbindyo, C.; Omani, R.; Chemweno, V. Review of Sheep Pox Disease in Sheep. J. Vet. Med. Res. 2017, 4, 1068. [Google Scholar]
- Gulyaz, V. Investigation of immunity in lambs born to ewes vaccinated with sheep and goat pox vaccine. Pendik Veteriner Mikrobiyoloji Dergisi 1999, 30, 55–62. [Google Scholar]
- Rodriguez, J.F.; Janeczko, R.; Esteban, M. Isolation and characterization of neutralizing monoclonal antibodies to vaccinia virus. J. Virol. 1985, 56, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Gordon, J.; Mohandas, A.; Wilton, S.; Dales, S. A prominent antigenic surface polypeptide involved in the biogenesis and function of the vaccinia virus envelope. Virology 1991, 181, 671–686. [Google Scholar] [CrossRef]
- Hsiao, J.C.; Chung, C.S.; Chang, W. Vaccinia virus envelope D8L protein binds to cell surface chondroitin sulfate and mediates the adsorption of intracellular mature virions to cells. J. Virol 1999, 73, 8750–8761. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Animal Health and Welfare (AHAW). Scientific Opinion on sheep and goat pox. EFSA J. 2014, 12, 3885. [Google Scholar] [CrossRef]
- EFSA Panel on Animal Health and Welfare (AHAW). Scientific Opinion on Lumpy Skin Disease. EFSA J. 2015, 13, 3986. [Google Scholar]
- Munyanduki, H.M. Improvement of the Capripoxvirus, lumpy Skin Disease Virus for Use as a Vaccine Vector. Ph.D. Thesis, Faculty of Health Sciences Department of Pathology, University of Cape Town, Cape Town, South Africa, 2018. Available online: http://hdl.handle.net/11427/30034 (accessed on 19 February 2021).
- Kalra, S.K.; Sharma, V.K. Adaptation of Jaipur strain of sheeppox virus in primary lamb testicular cell culture. Indian J. Exp. Biol. 1981, 19, 165–169. [Google Scholar]
- Das, S.K.; Pandey, A.K.; Mallick, B.B. A nore on rhe nat–ural goat pox outbreak in Garwal Hills of Uttar Pradesh. Indian Vet. J. 1978, 55, 671–673. [Google Scholar]
- Van Rooyen, P.J.; Munz, E.K.; Weiss, K.E. The optimal conditions for the multi–plication of Neethling-type Lumpy Skin Disease virus in embryonated eggs. Onderstepoort J. Vet. Res. 1969, 36, 165–174. [Google Scholar]
- Bhanuprakash, V.; Hosamani, M.; Juneja, S.; Kumar, N.; Singh, R.K. Detection of goat pox antibodies, comparative efficacy of indirect ELISA and ounterimmunoelectrophoresis. J. Appl. Anim. Res. 2006, 30, 177–180. [Google Scholar] [CrossRef]
- Ramyar, H.; Hessami, M. Development of an attenuated live vaccine against sheep-pox. Arch. Inst. Razi 1968, 20, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.B.; Ergm, H.; Koylu, A. Tests on sheep of attenuated sheep pox vaccines. Res. Vet. Sci. 1973, 14, 53. [Google Scholar] [CrossRef]
- Penkova, V.M.; Jasslm, F.A.; Thompson, J.R.; AI-Doon, T.M. The propagation of an attenuated sheep pox virus and its use as a vaccine. Bull. Off. Int. Eptzoot. 1974, 81, 329–339. [Google Scholar]
- Mateva, V.; Stoichev, S. Adaptation and cultivation of sheep-pox virus (strain Perego) in tissue cultures and evaluation of its immunogenic properties. Vet. Nauk. 1975, 12, 18–23. [Google Scholar]
- Anandan, R.; Sundara Rajan, S.; Kannamani, G.; Jayaraman, M.S. Studies on live and inactivated sheep pox vaccines. Cherion 1972, 1, 42–55. [Google Scholar]
- Abbas, F.; Khan, F.A.; Hussain, A.; Ahmed, F.; Ahmed, M.; Ahmed, S.; Ahmed, M.Z.; Attique, M.A.; Wadood, A.; Taj, M.K. Production of goatpox virus vaccine from a live attenuated goatpox virus strain. J. Anim. Plant Sci. 2010, 20, 315–317. [Google Scholar]
- Hosamani, M.; Nandi, S.; Mondal, B.; Singh, R.K.; Rasool, T.J.; Bandyopadhyay, S.K. A vero cell-attenuated goat pox virus provides protection against virulent virus challenge. Acta Virol. 2004, 48, 15–21. [Google Scholar] [PubMed]
- Dubey, S.C.; Sawhney, A.M. Live and reactivated tissue culture vaccines against goat pox. Indian Vet. J. 1978, 55, 925. [Google Scholar]
- Capstick, P.B.; Prydle, J.; Coackley, W.; Burdm, M.L. Protection of cattle against the Neethhng type virus of lumpy skin disease. Vet. Rec. 1959, 71, 422–423. [Google Scholar]
- Davies, F.G.; Mbugwa, G. The alterations in pathogenicity and immunogenicity of a Kenya sheep and goat pox virus on serial passage in bovine foetal cell cultures. J. Comp. Pathol. 1985, 95, 565–572. [Google Scholar] [CrossRef]
- Davies, F.G. Characteristics of a virus causing a pox disease in sheep and goats on Kenya, with observations on the epidemiology and control. J. Hyg. (Camb.) 1976, 76, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Kitching, R.P.; Smale, C. Comparison of the external dimensions of capripoxvirus isolates. Res. Vet. Sci. 1986, 41, 425–427. [Google Scholar] [CrossRef]
- Chaudhary, S.S.; Pandey, K.D.; Singh, R.P.; Verma, P.C.; Gupta, P.K. A Vero cell derived combined vaccine against sheep pox and Peste des petits ruminants for sheep. Vaccine 2009, 27, 2548–2553. [Google Scholar] [CrossRef] [PubMed]
- Yogisharadhya, R.; Bhanuprakash, V.; Hosamani, M.; Venkatesan, G.; Balamurugan, V.; Bora, D.P.; Bhanot, V.; Prabhu, M.; Singh, R.K. Comparative efficacy of live replicating sheepox vaccine strains in Ovines. Biologicals 2011, 39, 417–423. [Google Scholar] [CrossRef]
- Sevik, M.; Dogan, M. Epidemiological and Molecular Studies on Lumpy Skin Disease Outbreaks in Turkey during 2014-2015. Transbound. Emerg. Dis. 2016, 64, 1268–1279. [Google Scholar] [CrossRef]
- Kitching, R.P. Vaccines for Lumpy Skin Disease, Sheep Pox and Goat Pox. Vaccines for OIE List A and Emerging Animal Diseases. In Proceedings of the Symposium, Ames, IA, USA, 16–18 September 2003; pp. 161–167. [Google Scholar]
- Le Goff, C.; Fakhfakh, E.; Chadeyras, A.; Aba-Adulugba, E.; Libeau, G.; Hammami, S.; Diallo, A.; Albina, E. Host-range phylogenetic grouping of capripoxviruses: Genetic typing of CaPVs. In Applications of Gene-Based Technologies for Improving Animal Production and Health in Developing Countries; Makkar, H.P.S., Viljoen, G.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 727–733. [Google Scholar]
- Tuppurainen, E.S.M.; Pearson, C.R.; Bachanek-Bankowska, K.; Knowles, N.J.; Amareen, S.; Frost, L.; Henstock, M.R.; Lamien, C.E.; Diallo, A.; Mertens, P.P.C. Characterization of sheep pox virus vaccine for cattle against Lumpy Skin Disease virus. Antivir. Res. 2014, 109, 1–6. [Google Scholar] [CrossRef]
- Kara, P.D.; Afonso, C.L.; Wallace, D.B.; Kutish, G.F.; Abolnik, C.; Lu, Z.; Vreede, F.T.; Taljaard, L.C.F.; Zsak, A.; Viljoen, G.J.; et al. Comparative sequence analysis of the South African vaccine strain and two virulent field isolates of Lumpy Skin Disease virus. Arch. Virol. 2003, 148, 1335–1356. [Google Scholar] [CrossRef]
- Mathijs, E.; Vandenbussche, F.; Haegeman, A.; King, A.; Nthangeni, B.; Potgieter, C.; Maartens, L.; Van Born, S.; De Clercq, K. Complete genome sequences of the Neethling-Like Lumpy Skin Disease virus strains obtained directly from three com–mercial live attenuated vaccines. Genome Announc. 2016, 4, e01255-16. [Google Scholar] [CrossRef]
- Katsoulos, P.D.; Chaintoutis, S.C.; Dovas, C.I.; Polizopoulou, Z.S.; Brellou, G.D.; Agianniotaki, E.I.; Tasioudi, K.E.; Chondrokouki, E.; Papadopoulos, O.; Karatzias, H.; et al. Investigation on the incidence of adverse reactions, viraemia and haematological changes following field immunization of cattle using a live attenuated vaccine against Lumpy Skin Disease. Transbound. Emerg. Dis. 2018, 65, 174–185. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Lumpy Skin Disease: I. Data collection and analysis. EFSA J. 2017, 15, 4773. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); Calistri, P.; DeClercq, K.; Gubbins, S.; Klement, E.; Stegeman, A.; Cortinas Abrahantes, J.; Antoniou, S.E.; Broglia, A.; Gogin, A. Scientific report on Lumpy Skin Disease: III. Data collection and analysis. EFSA J. 2019, 17, 5638. [Google Scholar] [CrossRef]
- Agianniotaki, E.I.; Chaintoutis, S.C.; Haegeman, A.; Tasioudi, K.E.; De Leeuw, I.; Katsoulos, P.D.; Sachpatzidis, A.; De Clercq, K.; Alexandropoulos, T.; Polizopoulou, Z.S.; et al. Development and validation of a TaqMan probe-based real-time PCR method for the differentiation of wild type Lumpy Skin Disease virus from vaccine virus strains. J. Virol. Methods 2017, 249, 48–57. [Google Scholar] [CrossRef]
- Bedekovic, T.; Simic, I.; Kresic, N.; Lojkic, I. Detection of Lumpy Skin Disease virus in skin lesions, blood, nasal swabs and milk following preventive vaccination. Transbound. Emerg. Dis. 2017, 65, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Yeruham, I.; Perl, S.; Nyska, A.; Abraham, A.; Davidson, M.; Haymovitch, M.; Zamir, O.; Grinstein, H. Adverse reactions in cattle to a capripox vaccine. Vet. Rec. 1994, 135, 330–332. [Google Scholar] [CrossRef] [PubMed]
- Salib, F.A.; Osman, A.H. Incidence of Lumpy Skin Disease among Egyptian cattle in Giza Governorate, Egypt. Vet. World 2011, 4, 162–167. [Google Scholar]
- Kitching, R.P.; Hammond, J.M.; Taylor, W.P. A single vaccine for the control of capripox infection in sheep and goats. Res. Vet. Sci. 1987, 42, 53–60. [Google Scholar] [CrossRef]
- Achour, H.A.; Bouguedour, R.; Bouhbal, A.; Guechtouli, A.; Aouissat, M. Étude comparative du pouvoir immunisant de quelques souches atténuées de virus de la clavelée et d’un vaccin sensibiliser. Rev. Sci. Tech. Oie 2000, 19, 773–783. [Google Scholar] [CrossRef]
- Bhanuprakash, V.; Hosamani, M.; Singh, R.K. Prospects of control and eradication of capripox from the Indian subcontinent: A perspective. Antivir. Res. 2011, 91, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Awad, M.; Michael, A.; Soliman, S.M.; Samir, S.S.; Daoud, A.M. Trials for preparation of inactivated sheep pox vaccine using binary ethyleneimine. Egypt. J. Immunol. 2003, 10, 67–72. [Google Scholar]
- Kadymov, R.A. Combined immunization of sheep against anthrax, sheep pox and clostridial infections. Vet. Mosc. 1975, 2, 50–52. [Google Scholar]
- Selyaninov, Y.O.; Balyshev, V.M.; Kosyachenko, N.S.; Egorova, I.Y. Experimental assessment of the efficacy of associated vaccine against anthrax and sheep pox. Russ. Agric. Sci. 2003, 43–45. [Google Scholar]
- Hosamani, M.; Singh, S.K.; Mondal, B.; Sen, A.; Bhanuprakash, V.; Bandyopadhyay, S.K.; Yadav, M.P.; Singh, R.K. A bivalent vaccine against goat pox and peste des petits ruminants induces protective immune response in goats. Vaccine 2006, 24, 6058–6064. [Google Scholar] [CrossRef]
- Fakri, F.; Ghzal, F.; Daouam, S.; Elarkam, A.; Douieb, L.; Zouheir, Y.; Tadlaoui, K.; Fassi-Fihri, O. Development and field application of a new combined vaccine against Peste des Petits Ruminants and Sheep Pox. Trials Vaccinol. 2015, 4, 33–37. [Google Scholar] [CrossRef]
- Fakri, F.Z.; Embarki, T.; Baha, W.; Tadlaoui, K.O.; Fihri, O.F. Large mass vaccination of small ruminants against Peste des Petits Ruminants and Sheeppox using a combined live attenuated vaccine. J. Vet. Med. Res. 2020, 7, 1200. [Google Scholar]
- Wallace, D.; Ellis, C.; Espach, A.; Smith, S.; Greyling, R.; Viljoen, G. Protective immune responses induced by different recombinant vaccine regimes to Rift Valley Fever. Vaccine 2006, 24, 7181–7189. [Google Scholar] [CrossRef]
- Wallace, D.B.; Mather, A.; Kara, P.D.; Naicker, L.; Mokoena, N.B.; Pretorius, A.; Nefefe, T.; Thema, N.; Babiuk, S. Protection of Cattle Elicited Using a Bivalent Lumpy Skin Disease Virus-Vectored Recombinant Rift Valley Fever Vaccine. Front. Vet. Sci. 2020, 7, 256. [Google Scholar] [CrossRef]
- Diallo, A.; Minet, C.; Berhe, G.; Le Goff, C.; Black, D.N.; Fleming, M.; Barrett, C.; Libeau, G. Goat immune response to Capripox vaccine expressing the hemagglutinin protein of Pest des Petits Ruminants. Ann. N. Y. Acad. Sci. 2002, 969, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Hu, S.; Qu, L.; Hu, Q.; Zhang, Q.; Zhi, H.; Huang, K.; Bu, Z. A goat poxvirus-vectored peste-des-petits-ruminants vaccine induces long-lasting neutralization antibody to high levels in goats and sheep. Vaccine 2010, 28, 4742–4750. [Google Scholar] [CrossRef]
- Fakri, F.; Bamouh, Z.; Ghzal, F.; Baha, W.; Tadlaoui, K.; Fihri, O.F.; Elharrak, M. Comparative evaluation of three capripoxvirus- vectored peste des petits ruminants vaccines. Virology 2018, 514, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Sun, Y.; Chen, W.; Bu, Z. The 135 gene of goatpox virus encodes an inhibitor of NF-kappaB and apoptosis and may serve as an improved insertion site to generate vectored live vaccine. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Romero, C.H.; Barrett, T.; Evans, S.A.; Kitching, R.P.; Gershon, P.D.; Bostock, C.; Black, D.N. Single Capripoxvirus recombinant vaccine for the protection of cattle against Rinderpest and Lumpy Skin Disease. Vaccine 1993, 11, 737–742. [Google Scholar] [CrossRef]
- Wade-Evans, A.M.; Romero, C.H.; Mellor, P.; Takamatsu, H.; Anderson, J.; Thevasagayam, J.; Fleming, M.J.; Mertens, P.P.; Black, D.N. Expression of the major core structural protein (VP7) of bluetongue virus, by a recombinant capripox virus, provides partial protection of sheep against a virulent heterotypic bluetongue virus challenge. Virology 1996, 220, 227–231. [Google Scholar] [CrossRef]
- Perrin, A.; Albina, E.; Bréard, E.; Sailleau, C.; Promé, S.; Grillet, C.; Kwiatek, O.; Russo, P.; Thiéry, R.; Zientara, S.; et al. Recombinant capripoxviruses expressing proteins of bluetongue virus: Evaluation of immune responses and protection in small ruminants. Vaccine 2007, 25, 6774–6783. [Google Scholar] [CrossRef]
- Ma, W.; Wei, J.; Wei, Y.; Guo, H.; Jin, Y.; Xue, Y.; Wang, Y.; Yi, Z.; Liu, L.; Huang, J.; et al. Immunogenicity of the capsid precursor and a nine-amino-acid site-directed mutant of the 3C protease of foot-and-mouth disease virus coexpressed by a recombinant goatpox virus. Arch. Virol. 2014, 159, 1715–1722. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Fan, X.; Li, L.; Ren, W.; Han, X.; Wu, X.; Wang, Z. Development of recombinant goatpox virus expressing Echinococcus granulosus EG95 vaccine antigen. J. Virol. Methods 2018, 261, 28–33. [Google Scholar] [CrossRef]
- Sun, Z.; Liu, L.; Zhang, H.; Li, Y.; Wei, F.; Li, Z.; Wang, P.; Fu, Q.; Ren, Y.; Zhang, Y.; et al. Expression and functional analysis of Brucella outer membrane protein 25 in recombinant goat pox virus. Mol. Med. Rep. 2019, 19, 2323–2329. [Google Scholar] [CrossRef]
- Boshra, H.; Truong, T.; Nfon, C.; Bowden, T.R.; Gerdts, V.; Tikoo, S.; Babiuk, L.A.; Kara, P.; Mather, A.; Wallace, D.B.; et al. A lumpy skin disease virus deficient of an IL-10 gene homologue provides protective immunity against virulent capri–poxvirus challenge in sheep and goats. Antivir. Res. 2015, 123, 39–49. [Google Scholar] [CrossRef]
- Molla, W.; Frankena, K.; Gari, G.; Kidane, M.; Shegu, D.; de Jong, M.C.M. Seroprevalence and risk factors of lumpy skin disease in Ethiopia. Prev. Vet. Med. 2018, 160, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Haegeman, A.; De Leeuw, I.; Mostin, L.; Van Campe, W.; Aerts, L.; Vastag, M.; De Clercq, K. An Immunoperoxidase Monolayer Assay (IPMA) for the detection of lumpy skin disease antibodies. J. Virol. Methods 2019, 113800. [Google Scholar] [CrossRef]
- Haegeman, A.; De Vleeschauwer, A.; De Leeuw, I.; Vidanović, D.; Šekler, M.; Petrović, T.; Demarez, C.; Lefebvre, D.; De Clercq, K. Overview of diagnostic tools for Capripox virus infections. Prev. Vet. Med. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kitching, R.P. Progress towards sheep and goat pox vaccines. Vaccine 1983, 1, 4–9. [Google Scholar] [CrossRef]
- Haqiqi, S.; Hedayati, Z.; Varshuei, H.R.; Aqa Ebrahimiyan, M.; Khedmati, K.A.D. Study of Safety and Immunogenicity of Goat Pox Vaccine against Sheep Pox in Susceptible Sheep; Agricultural Scientific Information and Documentation Centre, Agricultural Research and Education Organization: Tehran, Iran, 2008. [Google Scholar]
- Capstick, P.B.; Cocackley, W. Protection of Cattle against Lumpy Skin Disease. I. Trials with a vaccine against Neethling type infection. Cab Rev. Perspect. Agric. Vet. Sci. Nutr. Nat. Resour. 1961, 2, 362–368. [Google Scholar]
- Brenner, J.; Bellaiche, M.; Gross, E.; Elad, D.; Oved, Z.; Haimo–vitz, M.; Wasserman, A.; Friedgut, O.; Stram, Y.; Bumbarov, V.; et al. Appearance of skin lesions in cattle populations vaccinated against Lumpy Skin Disease: Statutory challenge. Vaccine 2009, 27, 1500–1503. [Google Scholar] [CrossRef]
- Solyom, F.; Perenlei, L.; Roith, J. A live a~enuated virus vaccine against sheep pox. Acta Vet. Acad. Scient Hung. 1980, 28, 389. [Google Scholar]
- Abd-Elfatah, E.B.; El-Mekkawi, M.F.; Aboul-Soud, E.A.; Fawzi, E.M.; El-Soally, S.A. Immunological response of a new trivalent capripoxvirus vaccine in pregnant ewes and does. Slov. Vet. Res. 2019, 56 (Suppl. 22), 445–455. [Google Scholar] [CrossRef]
- Prasad, I.J.; Datt, N.S. Observation on the use of live and inactivated vaccines ag inst goat pox. Indian Vet. J. 1973, 50, 1–10. [Google Scholar]
- Abu-Elzein, E. Observations on natural and experimental infection of sheep and goats with a virulent field Capripoxvirus with high affinity to goats. Vet. Arshiv 2003, 73, 119–131. [Google Scholar]
- Abutarbush, S.M.; Hananeh, W.M.; Ramadan, W.; Al Sheyab, O.M.; Alnajjar, A.R.; Al Zoubi, I.G.; Knowles, N.J.; Bachanek-Bankowsa, K.; Tuppurainen, E.S.M. Adverse reactions to field vaccination against Lumpy Skin Disease in Jordan. Transbound. Emerg. Dis. 2016, 63, 213–219. [Google Scholar] [CrossRef]
- Ali, A.A.; Esmat, M.; Attia, H.; Selim, A.; Abdel-Hamid, Y.M. Clinical and patho–logical studies on Lumpy Skin Disease in Egypt. Vet. Rec. 1990, 127, 549–550. [Google Scholar]
- Abdallah, F.M.; El Damaty, H.M.; Kotb, G.F. Sporadic cases of Lumpy Skin Disease among cattle in Sharkia province, Egypt: Genetic characterization of Lumpy Skin Disease virus isolates and pathological findings. Vet. World 2018, 11, 1150–1158. [Google Scholar] [CrossRef] [PubMed]
- Zeedan, G.S.G.; Mahmoud, A.H.; Abdalhamed, A.B.; Abd El-Razik, K.A.E.; Khafagi, M.H.; Abou Zeina, H.A.A. Detection of Lumpy Skin Disease virus in cattle using real-time polymerase chain reaction and serological diagnostic assays in different governorates in Egypt in 2017. Vet. World 2019, 12, 1093–1100. [Google Scholar] [CrossRef]
- Norian, R.; Afzal Ahagran, N.; Varshovi, H.R.; Azadmehr, A. Comparative efficacy of two heterologous capripox vaccines to control Lumpy Skin Disease in cattle. Bulg. J. Vet. Med. 2017, 22, 171–179. [Google Scholar] [CrossRef]
- Mikhael, C.A.; Ibrahim, M.M.; Saad, M.A. Efficacy of Alternative Vaccination with Attenuated Sheep Pox and Inactivated Lumpy Skin Disease Vaccines against Lumpy Skin Disease. SCVMJ 2016, 21, 125–142. [Google Scholar] [CrossRef][Green Version]
- Mikhael, C.A.; Nakhla, O.E.; Mohamed, N.A. Study on the capability of a dual capripox vaccine in protection of cattle against LSD infection. J. Vet. Med. Res. 2017, 24, 224–233. [Google Scholar] [CrossRef]
- Aboul-Soud, E.A.; Kafafy, M.H.; El-Suall, S.A.; Zaghloul, M.A.; Mikhael, C.A. Preparation of trivalent vaccine against Lumpy Skin Disease using different capripox viral strain. Int. J. Vet. Sci. Med. 2018, in press. [Google Scholar]
- Zhugunissov, K.; Bulatov, Y.; Orynbayev, M.; Kutumbetov, L.; Abduraimov, Y.; Shayakhmetov, Y.; Taranov, D.; Amanova, Z.; Mambetaliyev, M.; Absatova, Z.; et al. Goatpox virus (G20-LKV) vaccine strain elicits a protective response in cattle against Lumpy Skin Disease at challenge with Lumpy Skin Disease virulent field strain in a comparative study. Vet. Microbiol. 2020, 245, 108695. [Google Scholar] [CrossRef]


{kind=link}
| Isolate/Strain | Origin | Cells | Passages for Attenuation | Reference |
|---|---|---|---|---|
| SPV RM65 | Sheep | Sheep kidney cells | 30 | [101] |
| SPV Romania | Sheep | Lamb kidney cells | 40 | [88] |
| SPV Bakirkoy | Sheep | Calf kidney cells | 32 | [102] |
| SPV Rumania Fanar | Sheep | Lamb testis | 26 | [103] |
| SPV Perego | Sheep | Lamb testis and calf kidney | 11 times in lamb testis and 10 times in calf kidney | [104] |
| SPV Ranipet | Sheep | Sheep thyroid cells | 35 | [105] |
| GPV Gorgan | Goat | Kid kidney cortex cells | - | [106] |
| GPV Uttarkashi | Goat | Primary lamb testis cells and Vero cells | 34 on primary cells and 26 on Vero | [107] |
| GPV Mysore | Goats | Primary lamb testis cells | 25 | [108] |
| GPV Kedong | Sheep | Lamb testis cells | - | [109] |
| GPV Isiolo | Sheep | Lamb testis cells | - | [109] |
| LSD KSGP O 180 | Sheep | Bovine foetalmuscle cells | 18 | [110] |
| LSD KSGP O 240 | Sheep | Lamb testis cells | 13–27 | [111] |
| LSD Neethling | Cattle | Lamb kidney cells and chorioallantoic membranes (CAM) | 61 on primary cells and 20 on CAM | [99] |
| Vaccine/Strain | Safety and Protection | Reference | ||
|---|---|---|---|---|
| Sheep | Goat | Cattle | ||
| GPV Gorgan | Safe, partially protective | Safe and protective | Safe and protective | [56,106,154] |
| GPV Mysore | - | Safe and protective | - | [108] |
| GPV Uttarkashi | - | Safe and protective | - | [107] |
| GPV Kedong and isiolo | - | - | Safe and protective | [155] |
| SPV RM65 | Safe and protective | - | Partially protective | [21,156] |
| SPV Perego | Safe and protective | - | - | [103,157] |
| SPV Rumania Fanar | Safe and protective | - | - | |
| SPV Romania | Safe and protective | Safe and protective | Partially protective | [57,158] |
| SPV Bakirkoy | Safe and protective | - | Partially protective | [102,115] |
| LSD Neethling | Partially protective | - | Causes Neethling disease, protective | [23,57] |
| LSD KSGP 0180 | Safe and protective | Safe and protective | Safe and protective | [110] |
| LSD KSGP 0240 | Safe and protective | Safe and protective | Residual virulence, partial protection | [20,112,126,127] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hamdi, J.; Munyanduki, H.; Omari Tadlaoui, K.; El Harrak, M.; Fassi Fihri, O. Capripoxvirus Infections in Ruminants: A Review. Microorganisms 2021, 9, 902. https://doi.org/10.3390/microorganisms9050902
Hamdi J, Munyanduki H, Omari Tadlaoui K, El Harrak M, Fassi Fihri O. Capripoxvirus Infections in Ruminants: A Review. Microorganisms. 2021; 9(5):902. https://doi.org/10.3390/microorganisms9050902
Chicago/Turabian StyleHamdi, Jihane, Henry Munyanduki, Khalid Omari Tadlaoui, Mehdi El Harrak, and Ouafaa Fassi Fihri. 2021. "Capripoxvirus Infections in Ruminants: A Review" Microorganisms 9, no. 5: 902. https://doi.org/10.3390/microorganisms9050902
APA StyleHamdi, J., Munyanduki, H., Omari Tadlaoui, K., El Harrak, M., & Fassi Fihri, O. (2021). Capripoxvirus Infections in Ruminants: A Review. Microorganisms, 9(5), 902. https://doi.org/10.3390/microorganisms9050902