
Cold Atmospheric-Pressure Plasma Caused Protein Damage in Methicillin-Resistant Staphylococcus aureus Cells in Biofilms



Abstract
1. Introduction
2. Materials and Methods
2.1. Biofilm Assay
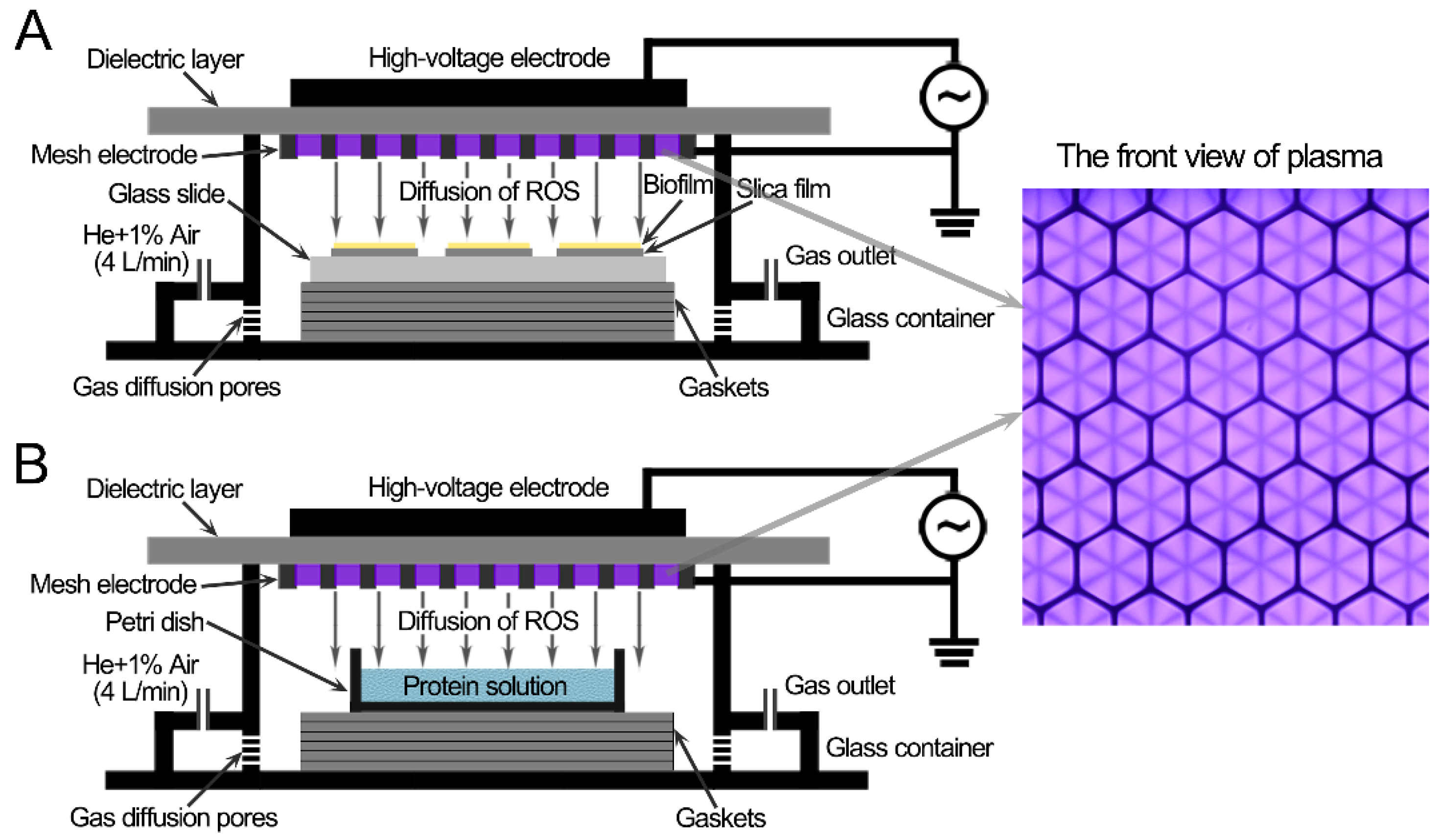
2.2. CAP Device and CAP Treatments
2.3. Protein Extraction and Detection
2.4. SaFtsZ and SaClpP Protein Expression and Purification
2.5. Immunoblotting
2.6. Blue Native Polyacrylamide Gel Electrophoresis
2.7. In Vitro Assembly Assay of SaFtsZ
2.8. Enzymatic Activity Assay of SaClpP
2.9. Mass Spectrometry of SaClpP
3. Results
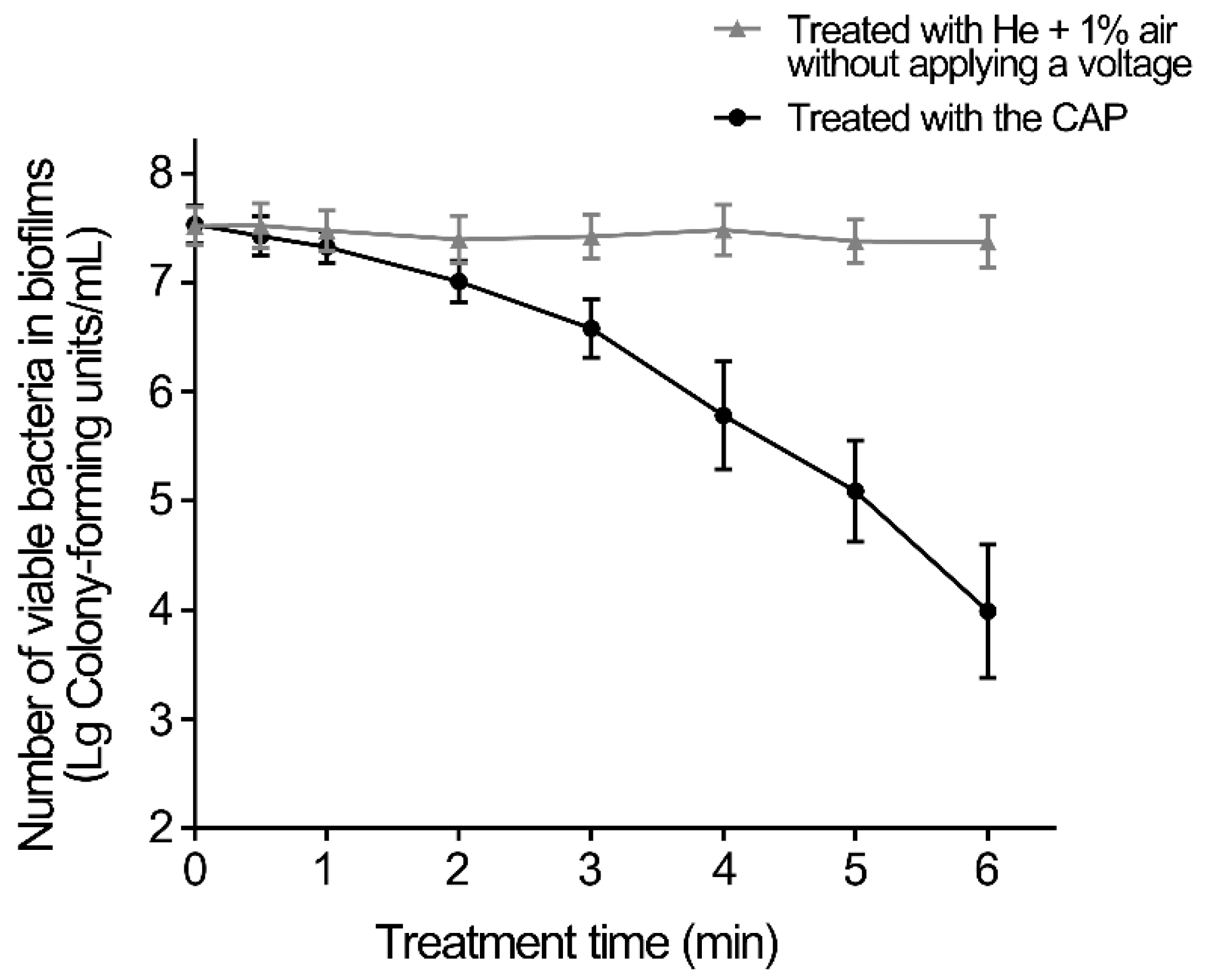
3.1. Inactivation of CAP Treatment on Biofilms
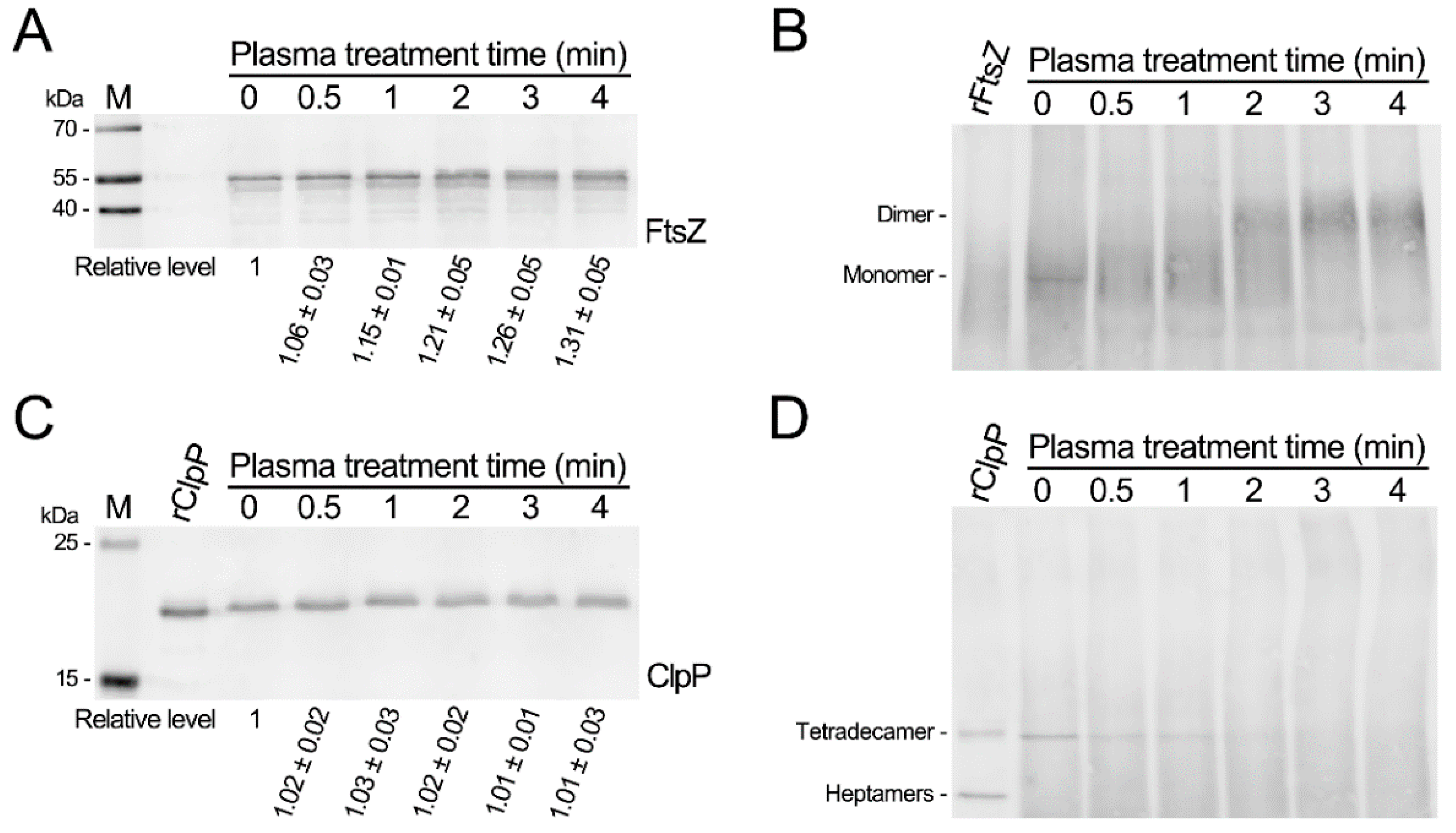
3.2. CAP Treatment Causes Polymerization of the SaFtsZ and SaClpP and Induces Oxidative Modification of Proteins
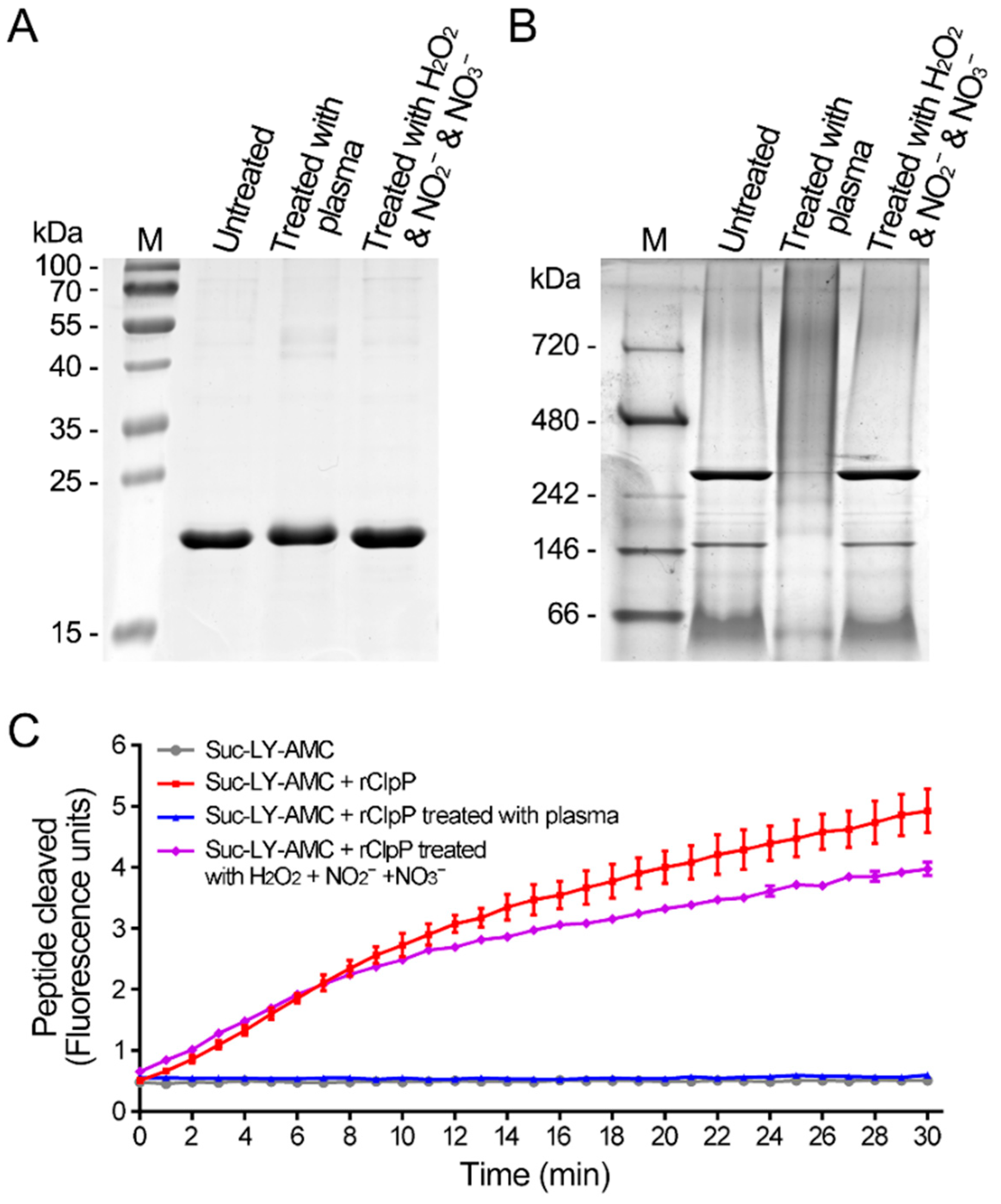
3.3. CAP Treatment Causes Inactivation of the SaFtsZ and SaClpP Proteins
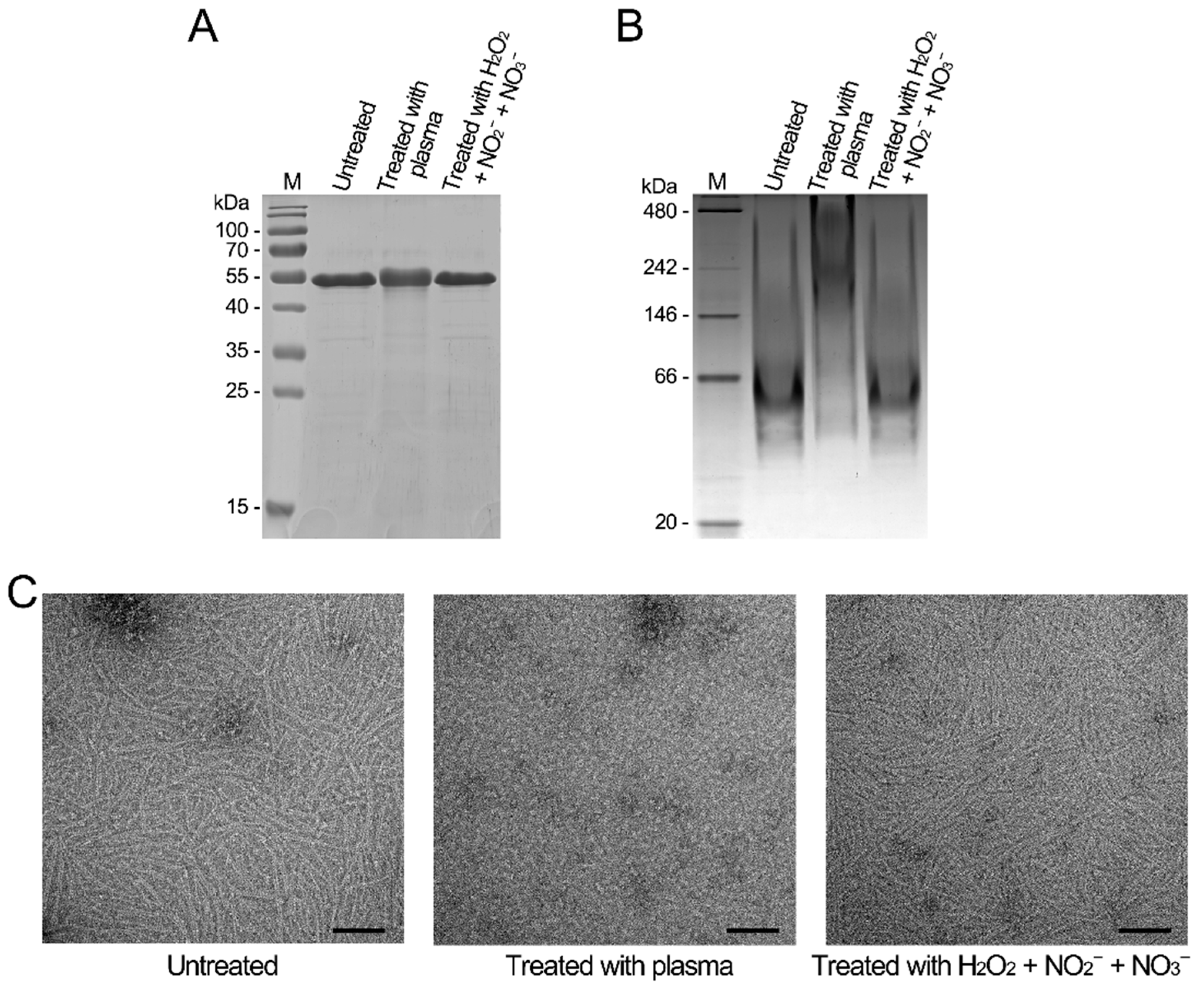
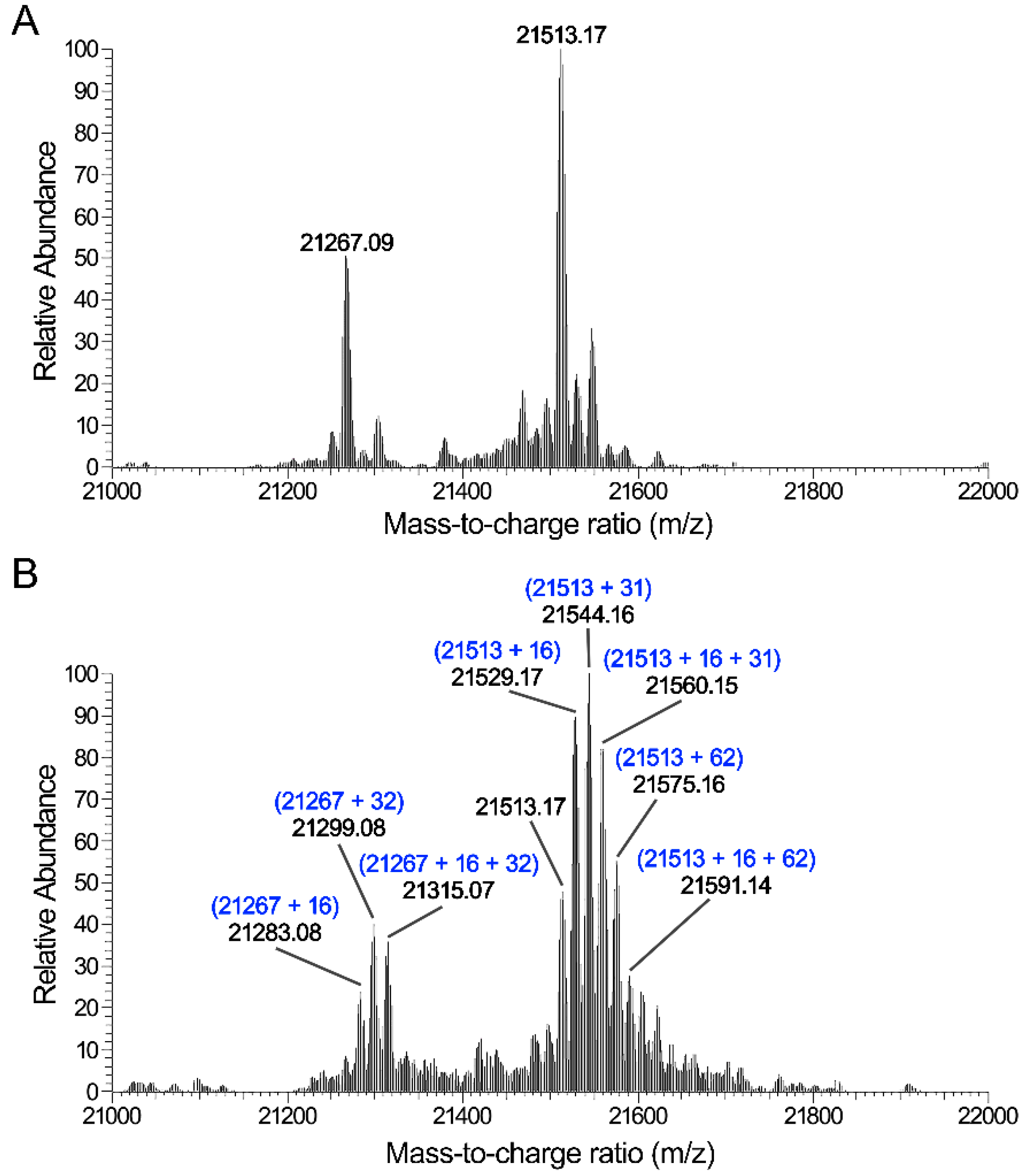
3.4. CAP Induces Oxidative Modification in Proteins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jain, S.; Sengupta, M.; Sarkar, S.; Ghosh, S.; Mitra, A.N.; Sinha, A.; Chakravorty, S. Can EDTA Change MRSA into MSSA? A Future Prospective! J. Clin. Diagn. Res. 2016, 10, DC22–DC25. [Google Scholar] [CrossRef] [PubMed]
- Zetola, N.; Francis, J.S.; Nuermberger, E.L.; Bishai, W.R. Community-acquired meticillin-resistant Staphylococcus aureus: An emerging threat. Lancet Infect. Dis. 2005, 5, 275–286. [Google Scholar] [CrossRef]
- Guillamet, C.V.; Kollef, M.H. How to stratify patients at risk for resistant bugs in skin and soft tissue infections? Curr. Opin. Infect. Dis. 2016, 29, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Bahamondez-Canas, T.F.; Heersema, L.A.; Smyth, H.D.C. Current status of in vitro models and assays for susceptibility testing for wound biofilm infections. Biomedicines 2019, 7, 34. [Google Scholar] [CrossRef]
- Akiyama, H.; Huh, W.K.; Yamasaki, O.; Oono, T.; Iwatsuki, K. Confocal laser scanning microscopic observation of glycocalyx production by Staphylococcus aureus in mouse skin: Does S. aureus generally produce a biofilm on damaged skin? Br. J. Dermatol. 2002, 147, 879–885. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Okuda, K.; Zendo, T.; Sugimoto, S.; Iwase, T.; Tajima, A.; Yamada, S.; Sonomoto, K.; Mizunoe, Y. Effects of bacteriocins on methicillin-resistant Staphylococcus aureus biofilm. Antimicrob. Agents Chemother. 2013, 57, 5572–5579. [Google Scholar] [CrossRef]
- Hoiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef]
- Mah, T.F.C.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Gao, P.; Ho, P.L.; Yan, B.; Sze, K.H.; Davies, J.; Kao, R.Y.T. Suppression of Staphylococcus aureus virulence by a small-molecule compound. Proc. Natl. Acad. Sci. USA 2018, 115, 8003–8008. [Google Scholar] [CrossRef]
- Del Pozo, J.L. Biofilm-related disease. Expert Rev. Anti Infect. Ther. 2018, 16, 51–65. [Google Scholar] [CrossRef]
- Fridman, G.; Friedman, G.; Gutsol, A.; Shekhter, A.B.; Vasilets, V.N.; Fridman, A. Applied Plasma Medicine. Plasma Process. Polym. 2008, 5, 503–533. [Google Scholar] [CrossRef]
- Kang, M.H.; Hong, Y.J.; Attri, P.; Sim, G.B.; Lee, G.J.; Panngom, K.; Kwon, G.C.; Choi, E.H.; Uhm, H.S.; Park, G. Analysis of the antimicrobial effects of nonthermal plasma on fungal spores in ionic solutions. Free Radic. Biol. Med. 2014, 72, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Kong, M.G.; Kroesen, G.; Morfill, G.; Nosenko, T.; Shimizu, T.; Van Dijk, J.; Zimmermann, J.L. Plasma medicine: An introductory review. New J. Phys. 2009, 11, 115012. [Google Scholar] [CrossRef]
- Moreau, M.; Orange, N.; Feuilloley, M.G. Non-thermal plasma technologies: New tools for bio-decontamination. Biotechnol. Adv. 2008, 26, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Bekeschus, S.; Clemen, R.; Niessner, F.; Sagwal, S.K.; Freund, E.; Schmidt, A. Medical gas plasma jet technology targets murine melanoma in an immunogenic fashion. Adv. Sci. 2020, 7, 1903438. [Google Scholar] [CrossRef] [PubMed]
- Lin, A.; Gorbanev, Y.; De Backer, J.; Van Loenhout, J.; Van Boxem, W.; Lemiere, F.; Cos, P.; Dewilde, S.; Smits, E.; Bogaerts, A. Non-thermal plasma as a unique delivery system of short-lived reactive oxygen and nitrogen species for immunogenic cell death in melanoma cells. Adv. Sci. 2019, 6, 1802062. [Google Scholar] [CrossRef]
- Guo, L.; Xu, R.B.; Zhao, Y.M.; Liu, D.X.; Liu, Z.J.; Wang, X.H.; Chen, H.L.; Kong, M.G. Gas Plasma Pre-treatment Increases Antibiotic Sensitivity and Persister Eradication in Methicillin-Resistant Staphylococcus aureus. Front. Microbiol. 2018, 9, 537. [Google Scholar] [CrossRef]
- Wang, J.; Yu, Z.; Xu, Z.; Hu, S.; Li, Y.; Xue, X.; Cai, Q.; Zhou, X.; Shen, J.; Lan, Y.; et al. Antimicrobial mechanism and the effect of atmospheric pressure N2 plasma jet on the regeneration capacity of Staphylococcus aureus biofilm. Biofouling 2018, 34, 935–949. [Google Scholar] [CrossRef]
- Xu, Z.; Shen, J.; Cheng, C.; Hu, S.; Lan, Y.; Chu, P.K. In Vitro antimicrobial effects and mechanism of atmospheric-pressure He/O2 plasma jet on Staphylococcus aureus biofilm. J. Phys. D Appl. Phys. 2017, 50, 105201. [Google Scholar] [CrossRef]
- Czapka, T.; Maliszewska, I.; Olesiak-Banska, J. Influence of atmospheric pressure non-thermal plasma on inactivation of biofilm cells. Plasma Chem. Plasma Process. 2018, 38, 1181–1197. [Google Scholar] [CrossRef]
- Milam, S.L.; Osawa, M.; Erickson, H.P. Negative-stain electron microscopy of inside-out FtsZ rings reconstituted on artificial membrane tubules show ribbons of protofilaments. Biophys. J. 2012, 103, 59–68. [Google Scholar] [CrossRef][Green Version]
- Zheng, Y.; Du, R.; Cai, S.; Liu, Z.; Fang, Z.; Liu, T.; So, L.; Lu, Y.; Sun, N.; Wong, K. Study of benzofuroquinolinium derivatives as a new class of potent antibacterial agent and the mode of inhibition targeting FtsZ. Front. Microbiol. 2018, 9, 1937. [Google Scholar] [CrossRef]
- Haydon, D.J.; Stokes, N.R.; Ure, R.; Galbraith, G.; Bennett, J.M.; Brown, D.R.; Baker, P.J.; Barynin, V.V.; Rice, D.W.; Sedelnikova, S.E.; et al. An inhibitor of FtsZ with potent and selective anti-staphylococcal activity. Science 2008, 321, 1673–1675. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Stewart, P.S.; Greenberg, E.P. Bacterial biofilms: A common cause of persistent infections. Science 1999, 284, 1318–1322. [Google Scholar] [CrossRef] [PubMed]
- Culp, E.; Wright, G.D. Bacterial proteases, untapped antimicrobial drug targets. J. Antibiot. 2017, 70, 366–377. [Google Scholar] [CrossRef]
- Ma, X.; Ehrhardt, D.W.; Margolin, W. Colocalization of cell division proteins FtsZ and FtsA to cytoskeletal structures in living Escherichia coli cells by using green fluorescent protein. Proc. Natl. Acad. Sci. USA 1996, 93, 12998–13003. [Google Scholar] [CrossRef]
- Romberg, L.; Simon, M.; Erickson, H.P. Polymerization of Ftsz, a bacterial homolog of tubulin: Is assembly cooperative? J. Biol. Chem. 2001, 276, 11743–11753. [Google Scholar] [CrossRef] [PubMed]
- Wagstaff, J.M.; Tsim, M.; Oliva, M.A.; Garcia-Sanchez, A.; Kureisaite-Ciziene, D.; Andreu, J.M.; Lowe, J. A polymerization-associated structural switch in FtsZ that enables treadmilling of model filaments. mBio 2017, 8, e002541-17. [Google Scholar] [CrossRef]
- Ortiz, C.; Natale, P.; Cueto, L.; Vicente, M. The keepers of the ring: Regulators of FtsZ assembly. FEMS Microbiol. Rev. 2016, 40, 57–67. [Google Scholar] [CrossRef]
- McCool, J.D.; Long, E.; Petrosino, J.F.; Sandler, H.A.; Rosenberg, S.M.; Sandler, S.J. Measurement of SOS expression in individual Escherichia coli K-12 cells using fluorescence microscopy. Mol. Microbiol. 2004, 53, 1343–1357. [Google Scholar] [CrossRef]
- Hu, J.; Zhang, X.; Liu, X.; Chen, C.; Sun, B. Mechanism of reduced vancomycin susceptibility conferred by walK mutation in community-acquired methicillin-resistant Staphylococcus aureus strain MW2. Antimicrob. Agents Chemother. 2015, 59, 1352–1355. [Google Scholar] [CrossRef] [PubMed]
- Llama-Palacios, A.; Potupa, O.; Sanchez, M.C.; Figuero, E.; Herrera, D.; Sanz, M. Aggregatibacter actinomycetemcomitans growth in biofilm versus planktonic state: Differential expression of proteins. J. Proteome Res. 2017, 16, 3158–3167. [Google Scholar] [CrossRef]
- Feng, J.; Michalik, S.; Varming, A.N.; Andersen, J.H.; Albrecht, D.; Jelsbak, L.; Krieger, S.; Ohlsen, K.; Hecker, M.; Gerth, U.; et al. Trapping and proteomic identification of cellular substrates of the ClpP protease in Staphylococcus aureus. J. Proteome Res. 2013, 12, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Conlon, B.P.; Nakayasu, E.S.; Fleck, L.E.; LaFleur, M.D.; Isabella, V.M.; Coleman, K.; Leonard, S.N.; Smith, R.D.; Adkins, J.N.; Lewis, K. Activated ClpP kills persisters and eradicates a chronic biofilm infection. Nature 2013, 503, 365–370. [Google Scholar] [CrossRef]
- Frees, D.; Gerth, U.; Ingmer, H. Clp chaperones and proteases are central in stress survival, virulence and antibiotic resistance of Staphylococcus aureus. Int. J. Med. Microbiol. 2014, 304, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Gersch, M.; Kolb, R.; Alte, F.; Groll, M.; Sieber, S.A. Disruption of oligomerization and dehydroalanine formation as mechanisms for ClpP protease inhibition. J. Am. Chem. Soc. 2014, 136, 1360–1366. [Google Scholar] [CrossRef] [PubMed]
- McGillivray, S.M.; Tran, D.N.; Ramadoss, N.S.; Alumasa, J.N.; Okumura, C.Y.; Sakoulas, G.; Vaughn, M.M.; Zhang, D.X.; Keiler, K.C.; Nizet, V. Pharmacological inhibition of the ClpXP protease increases bacterial susceptibility to host cathelicidin antimicrobial peptides and cell envelope-active antibiotics. Antimicrob. Agents Chemother. 2012, 56, 1854–1861. [Google Scholar] [CrossRef]
- Springer, M.T.; Singh, V.K.; Cheung, A.L.; Donegan, N.P.; Chamberlain, N.R. Effect of clpP and clpC deletion on persister cell number in Staphylococcus aureus. J. Med. Microbiol. 2016, 65, 848–857. [Google Scholar] [CrossRef]
- Michel, A.; Agerer, F.; Hauck, C.R.; Herrmann, M.; Ullrich, J.; Hacker, J.; Ohlsen, K. Global regulatory impact of ClpP protease of Staphylococcus aureus on regulons involved in virulence, oxidative stress response, autolysis, and DNA repair. J. Bacteriol. 2006, 188, 5783–5796. [Google Scholar] [CrossRef]
- Zhang, J.; Ye, F.; Lan, L.; Jiang, H.; Luo, C.; Yang, C.G. Structural switching of Staphylococcus aureus Clp protease: A key to understanding protease dynamics. J. Biol. Chem. 2011, 286, 37590–37601. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Hartling, J.A.; Flanagan, J.M. The structure of ClpP at 2.3 A resolution suggests a model for ATP-dependent proteolysis. Cell 1997, 91, 447–456. [Google Scholar] [CrossRef]
- Ye, F.; Li, J.H.; Yang, C.G. The development of small-molecule modulators for ClpP protease activity. Mol. Biosyst. 2017, 13, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Zhao, Y.; Liu, D.; Liu, Z.; Chen, C.; Xu, R.; Tian, M.; Wang, X.; Chen, H.; Kong, M.G. Cold atmospheric-pressure plasma induces DNA-protein crosslinks through protein oxidation. Free Radic. Res. 2018, 52, 783–798. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Xu, R.B.; Liu, D.X.; Qi, Y.; Guo, Y.H.; Wang, W.T.; Zhang, J.; Liu, Z.J.; Kong, M.G. Eradication of methicillin-resistant Staphylococcus aureus biofilms by surface discharge plasmas with various working gases. J. Phys. D Appl. Phys. 2019, 52, 425202. [Google Scholar] [CrossRef]
- Wittig, I.; Braun, H.P.; Schagger, H. Blue native PAGE. Nat. Protoc. 2006, 1, 418–428. [Google Scholar] [CrossRef]
- Thompson, M.W.; Singh, S.K.; Maurizi, M.R. Processive degradation of proteins by the ATP-dependent Clp protease from Escherichia coli. Requirement for the multiple array of active sites in ClpP but not ATP hydrolysis. J. Biol. Chem. 1994, 269, 18209–18215. [Google Scholar] [CrossRef]
- Li, D.H.; Chung, Y.S.; Gloyd, M.; Joseph, E.; Ghirlando, R.; Wright, G.D.; Cheng, Y.Q.; Maurizi, M.R.; Guarne, A.; Ortega, J. Acyldepsipeptide antibiotics induce the formation of a structured axial channel in ClpP: A model for the ClpX/ClpA-bound state of ClpP. Chem. Biol. 2010, 17, 959–969. [Google Scholar] [CrossRef]
- Beranova-Giorgianni, S.; Desiderio, D.M. Mass spectrometry of the human pituitary proteome: Identification of selected proteins. Rapid Commun. Mass Spectrom. 2000, 14, 161–167. [Google Scholar] [CrossRef]
- Itooka, K.; Takahashi, K.; Kimata, Y.; Izawa, S. Cold atmospheric pressure plasma causes protein denaturation and endoplasmic reticulum stress in Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2018, 102, 2279–2288. [Google Scholar] [CrossRef]
- Simpson, J.A.; Narita, S.; Gieseg, S.; Gebicki, S.; Gebicki, J.M.; Dean, R.T. Long-lived reactive species on free-radical-damaged proteins. Biochem. J. 1992, 282, 621–624. [Google Scholar] [CrossRef]
- Davies, M.J. Protein oxidation and peroxidation. Biochem. J. 2016, 473, 805–825. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. Reactive species formed on proteins exposed to singlet oxygen. Photochem. Photobiol. Sci. 2004, 3, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Davies, M.J. Singlet oxygen-mediated damage to proteins and its consequences. Biochem. Biophy. Res. Commun. 2003, 305, 761–770. [Google Scholar] [CrossRef]
- Huang, M.; Zhuang, H.; Zhao, J.; Wang, J.; Yan, W.; Zhang, J. Differences in cellular damage induced by dielectric barrier discharge plasma between Salmonella Typhimurium and Staphylococcus aureus. Bioelectrochemistry 2020, 132, 107445. [Google Scholar] [CrossRef] [PubMed]
- Kawano, A.; Yamasaki, R.; Sakakura, T.; Takatsuji, Y.; Haruyama, T.; Yoshioka, Y.; Ariyoshi, W. Reactive oxygen species penetrate persister cell membranes of Escherichia coli for effective cell killing. Front. Cell Infect. Microbiol. 2020, 10, 496. [Google Scholar] [CrossRef] [PubMed]
- Vatansever, F.; de Melo, W.C.; Avci, P.; Vecchio, D.; Sadasivam, M.; Gupta, A.; Chandran, R.; Karimi, M.; Parizotto, N.A.; Yin, R.; et al. Antimicrobial strategies centered around reactive oxygen species--bactericidal antibiotics, photodynamic therapy, and beyond. FEMS Microbiol. Rev. 2013, 37, 955–989. [Google Scholar] [CrossRef]
- Arjunan, K.P.; Sharma, V.K.; Ptasinska, S. Effects of atmospheric pressure plasmas on isolated and cellular DNA—A review. Int. J. Mol. Sci. 2015, 16, 2971–3016. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucleic Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Designation | Sequence (5′-3′) |
|---|---|
| ftsZ up | ATCATGCCATGGTAGAATTTGAACAAGGATTTA |
| ftsZ His-tag down | ATACCGCTCGAGACGTCTTGTTCTTCTTGAACGTC |
| clpP up | ATGGGAATTCCATATGAATTTAATTCCTACAGTTATTGAAAC |
| clpP His-tag down | ATACCGCTCGAGTTTTGTTTCAGGTACCATCACTTC |
| clpP down | ATACCGCTCGAGTTATTTTGTTTCAGGTACCATCACTTC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, L.; Yang, L.; Qi, Y.; Niyazi, G.; Huang, L.; Gou, L.; Wang, Z.; Zhang, L.; Liu, D.; Wang, X.; et al. Cold Atmospheric-Pressure Plasma Caused Protein Damage in Methicillin-Resistant Staphylococcus aureus Cells in Biofilms. Microorganisms 2021, 9, 1072. https://doi.org/10.3390/microorganisms9051072
Guo L, Yang L, Qi Y, Niyazi G, Huang L, Gou L, Wang Z, Zhang L, Liu D, Wang X, et al. Cold Atmospheric-Pressure Plasma Caused Protein Damage in Methicillin-Resistant Staphylococcus aureus Cells in Biofilms. Microorganisms. 2021; 9(5):1072. https://doi.org/10.3390/microorganisms9051072
Chicago/Turabian StyleGuo, Li, Lu Yang, Yu Qi, Gulimire Niyazi, Lingling Huang, Lu Gou, Zifeng Wang, Lei Zhang, Dingxin Liu, Xiaohua Wang, and et al. 2021. "Cold Atmospheric-Pressure Plasma Caused Protein Damage in Methicillin-Resistant Staphylococcus aureus Cells in Biofilms" Microorganisms 9, no. 5: 1072. https://doi.org/10.3390/microorganisms9051072
APA StyleGuo, L., Yang, L., Qi, Y., Niyazi, G., Huang, L., Gou, L., Wang, Z., Zhang, L., Liu, D., Wang, X., Chen, H., & Kong, M. G. (2021). Cold Atmospheric-Pressure Plasma Caused Protein Damage in Methicillin-Resistant Staphylococcus aureus Cells in Biofilms. Microorganisms, 9(5), 1072. https://doi.org/10.3390/microorganisms9051072