
In Vitro Fermentation Characteristics and Fiber-Degrading Enzyme Kinetics of Cellulose, Arabinoxylan, β-Glucan and Glucomannan by Pig Fecal Microbiota

 , ,
, ,  ,
,  ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Substrates
2.2. Preparation of Inocula
2.3. Cumulative Gas Production Trial
2.4. In Vitro Fermentation Trial
2.5. Samples Detection
2.5.1. Monosaccharides Composition
2.5.2. SCFA and Lactate Detection
2.5.3. Bacterial Community
2.5.4. Determination of Fiber-Degrading Enzymes Activity
2.6. Statistical Analysis
3. Results
3.1. Monosaccharides Composition of Cellulose, Arabinoxylan, β-Glucan and Glucomannan
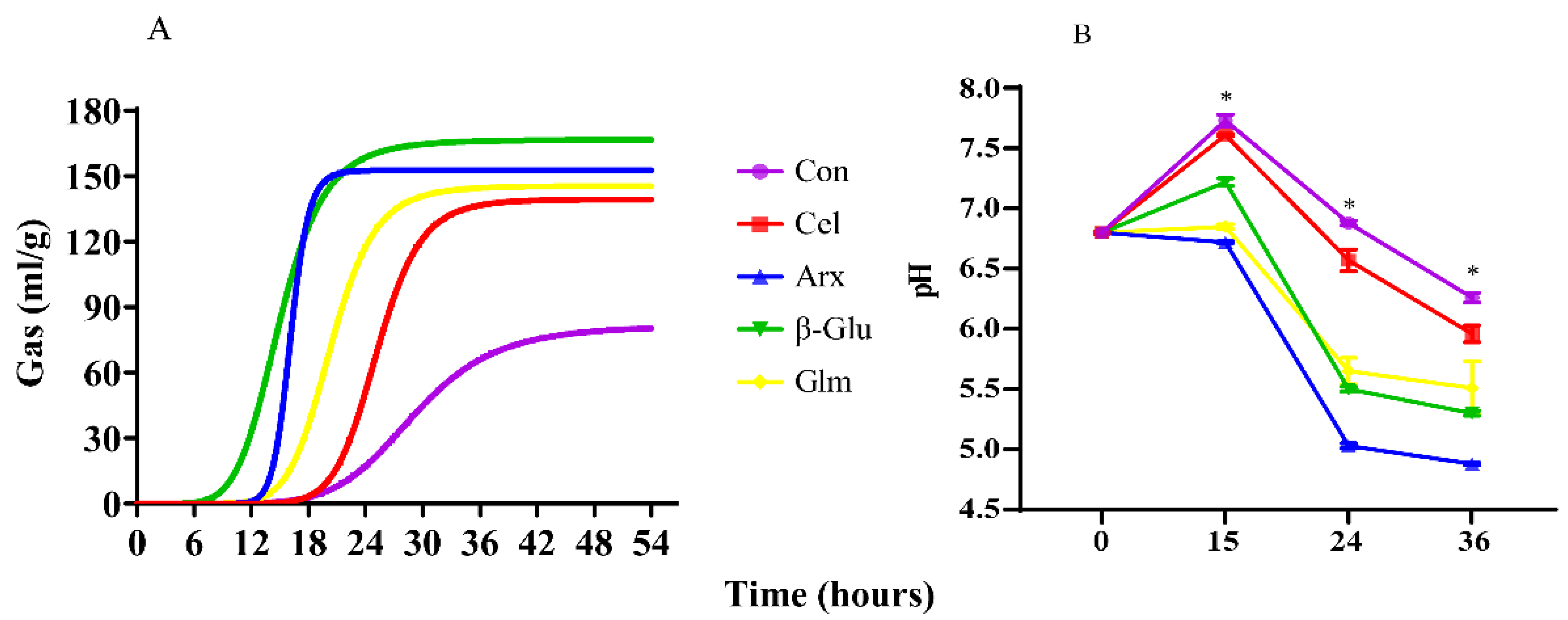
3.2. Gas Production and pH Shifts during Cellulose, Arabinoxylan, β-Glucan and Glucomannan Fermentation
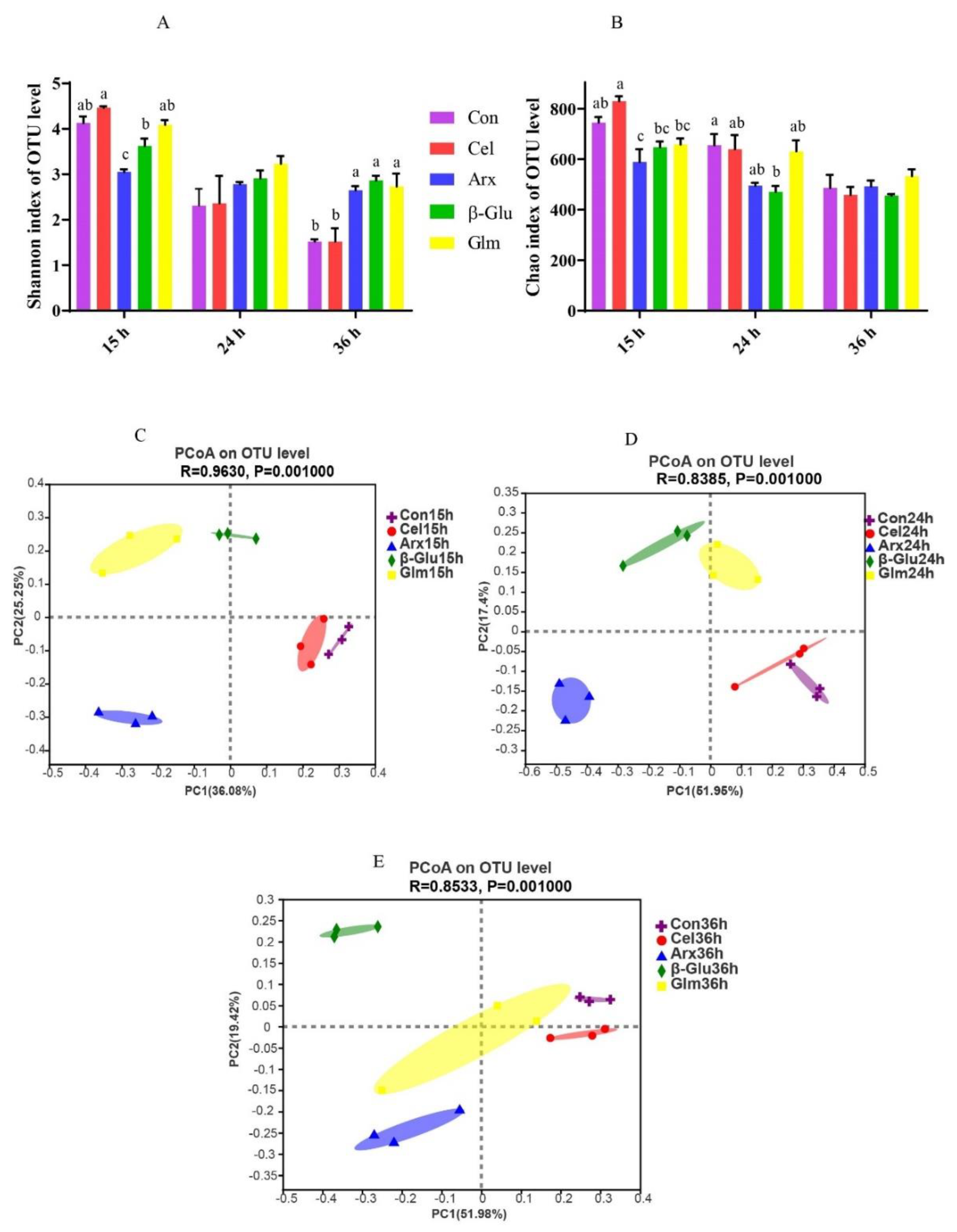
3.3. The Changes in Microbial Diversity during Cellulose, Arabinoxylan, β-Glucan and Glucomannan Fermentation
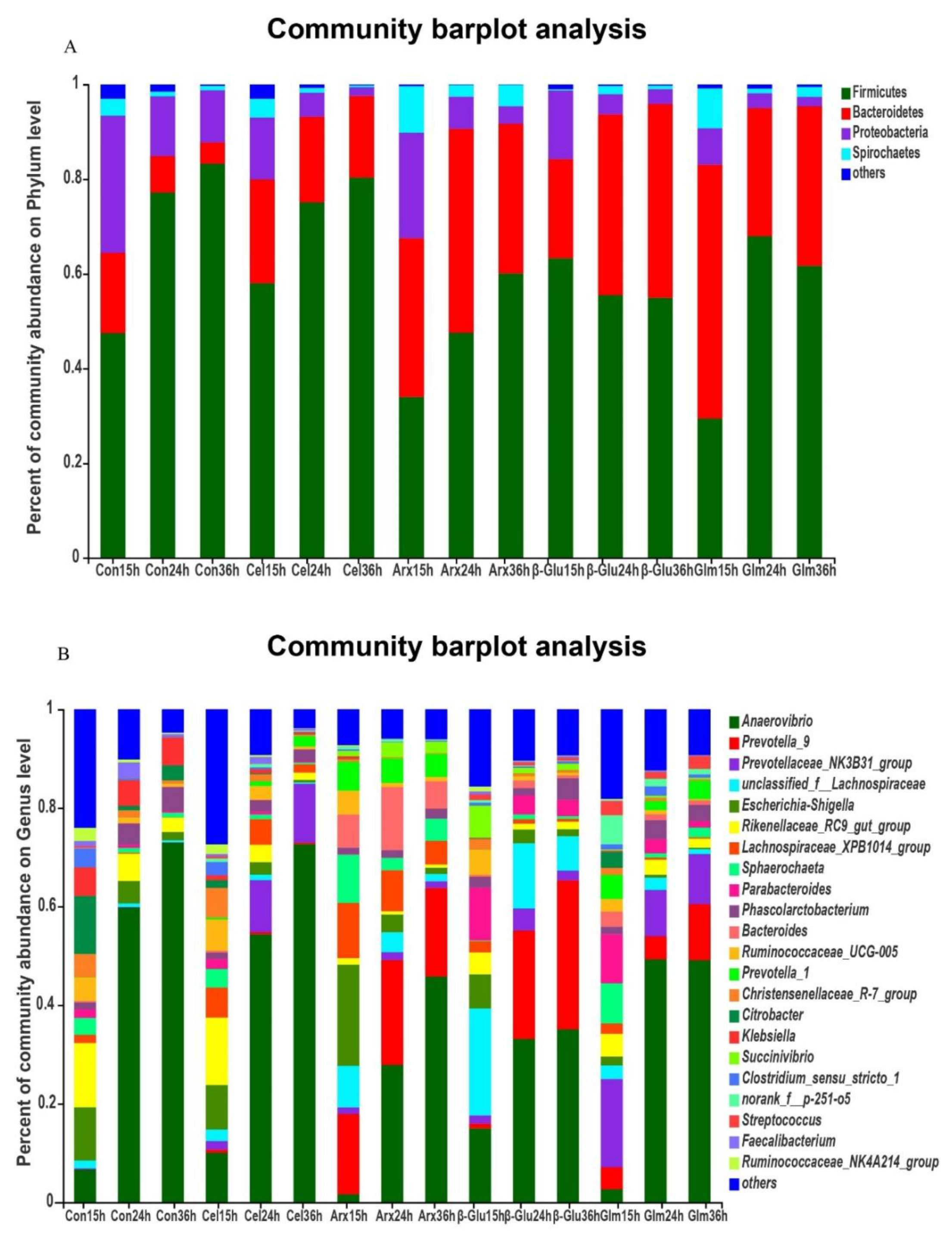
3.4. The Changes in the Microbial Composition during Cellulose, Arabinoxylan, β-Glucan and Glucomannan Fermentation
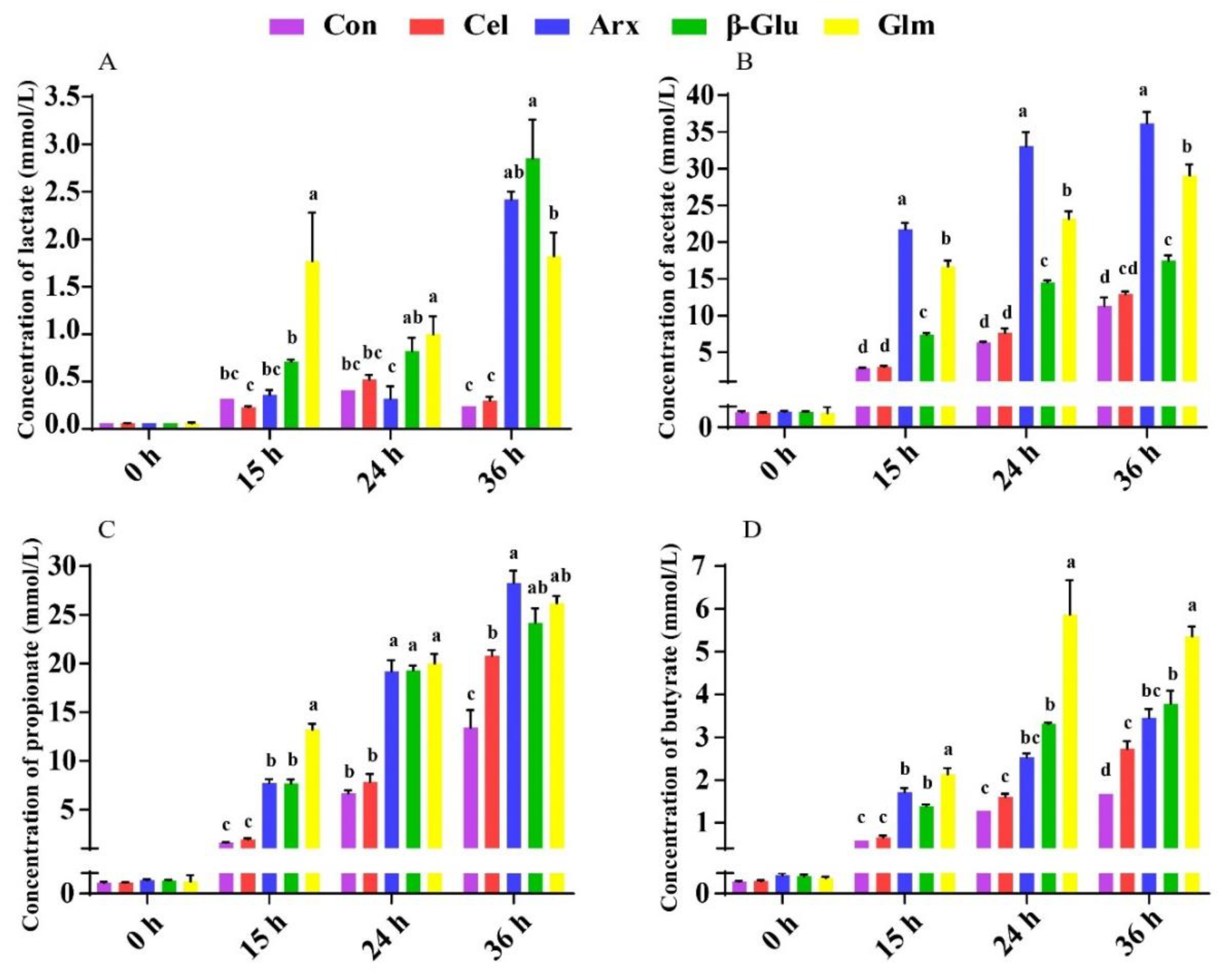
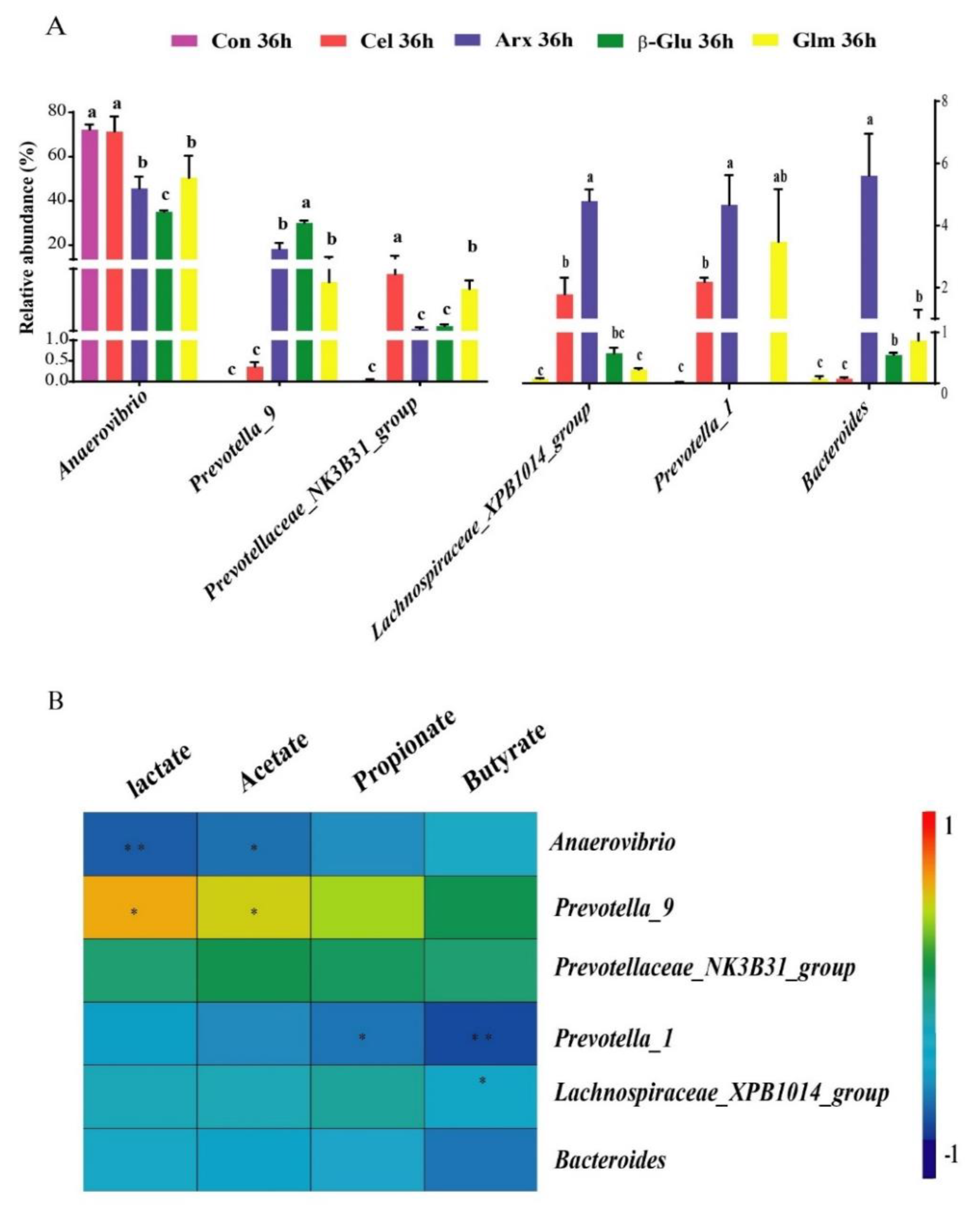
3.5. The Production of Lactate, SCFA and Their Relationship with the First Six Differential Bacteria
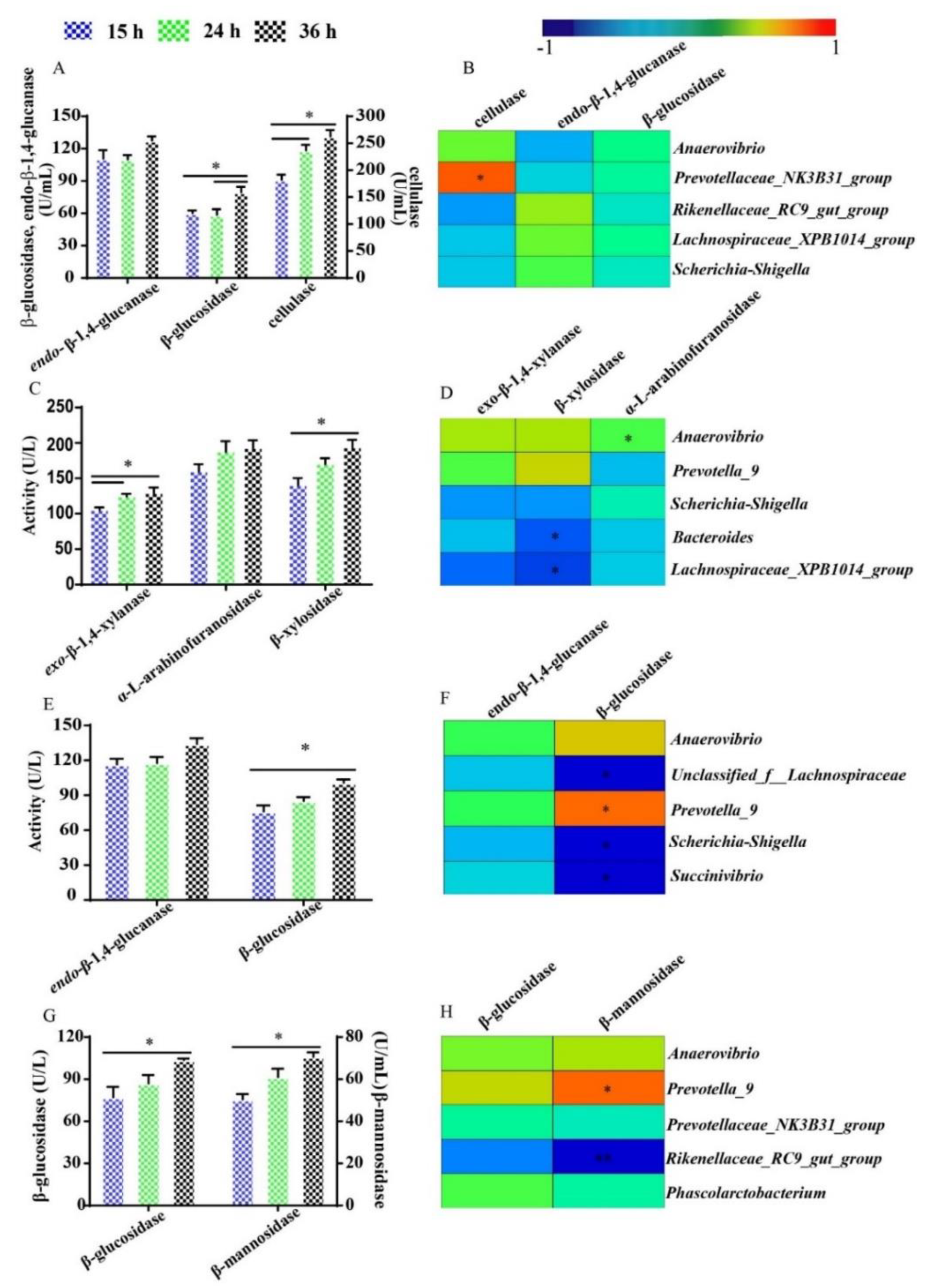
3.6. The Activity of Fiber-Degrading Enzymes and Their Relationship with Dominant Bacteria during Fermentation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Seo, Y.S.; Lee, H.B.; Kim, Y.; Park, H.Y. Dietary Carbohydrate Constituents Related to Gut Dysbiosis and Health. Microorganisms 2020, 8, 427. [Google Scholar]
- Jonathan, R.B.; David, A. Commensal Bacteria at the Interface of Host Metabolism and the Immune System. Nat. Immunol. 2013, 14, 676–684. [Google Scholar]
- Zhang, H.; Wielen, N.V.; Hee, B.V.; Wang, J.; Hendriks, W.; Gilbert, M. Impact of Fermentable Protein, by Feeding High Protein Diets, on Microbial Composition, Microbial Catabolic Activity, Gut Health and beyond in Pigs. Microorganisms 2020, 11, 1735. [Google Scholar] [CrossRef]
- Sokol, H.; Lay, C.; Seksik, P.; Tannock, G.W. Analysis of Bacterial Bowel Communities of IBD Patients: What has it Revealed? Inflamm. Bowel Dis. 2008, 14, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Nagy-Szakal, D.; Hollister, E.B.; Luna, R.A.; Szigeti, R.; Tatevian, N.; Smith, C.W.; Versalovic, J.; Kellermayer, R. Cellulose Supplementation Early in Life Ameliorates Colitis in Adult Mice. PLoS ONE 2013, 8, e56685. [Google Scholar]
- Kim, Y.; Hwang, S.W.; Kim, S.; Lee, Y.S.; Kim, T.Y.; Lee, S.H.; Kim, S.J.; Yoo, H.J.; Kim, E.N.; Kweon, M. Dietary Cellulose Prevents Gut Inflammation by Modulating Lipid Metabolism and Gut Microbiota. Gut Microbes. 2020, 11, 944–961. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wang, W.; Degroote, J.; Possemiers, S.; Chen, D.; De Smet, S.; Michiels, J. Arabinoxylan in Wheat is More Responsible than Cellulose for Promoting Intestinal Barrier Function in Weaned Male Piglets. J Nutr. 2015, 145, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Yu, Y.; Lin, D.; Zheng, P.; Zhang, P.; Hu, M.; Wang, Q.; Pan, W.; Yang, X.; Hu, T.; et al. β-Glucan Attenuates Cognitive Impairment via the Gut-Brain Axis in Diet-induced Obese Mice. Microbiome 2020, 8, 143. [Google Scholar] [CrossRef]
- Miyamoto, J.; Watanabe, K.; Taira, S.; Kasubuchi, M.; Li, X.; Irie, J.; Itoh, H.; Kimura, I. Barley β-Glucan Improves Metabolic Condition via Short-chain Fatty Acids Produced by Gut Microbial Fermentation in High Fat Diet Fed Mice. PLoS ONE 2018, 13, e0196579. [Google Scholar]
- Tanabe, K.; Nakamura, S.; Moriyama-Hashiguchi, M.; Kitajima, M.; Ejima, H.; Imori, C.; Oku, T. Dietary Fructooligosaccharide and Glucomannan Alter Gut Microbiota and Improve Bone Metabolism in Senescence-Accelerated Mouse. J. Agric. Food Chem. 2019, 67, 867–874. [Google Scholar] [CrossRef]
- Jonathan, M.C.; van den Borne, J.J.G.C.; Wiechen, P.V.; Silva, C.S.; da Scholsa, H.A. In Vitro Fermentation of 12 Dietary Fibers by Faecal Inoculum from Pigs and Humans. Food Chem. 2012, 133, 889–897. [Google Scholar] [CrossRef]
- Mikkelsen, D.; Gidley, M.J.; Williams, B.A. In Vitro Fermentation of Bacterial Cellulose Composites as Model Dietary Fibers. J. Agric. Food Chem. 2011, 59, 4025–4032. [Google Scholar] [CrossRef]
- Long, C.; Rösch, C.; de Vries, S.; Schols, H.; Venema, K. Cellulase and Alkaline Treatment Improve Intestinal Microbial Degradation of Recalcitrant Fibers of Rapeseed Meal in Pigs. J. Agric. Food Chem. 2020, 68, 11011–11025. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Flanagan, B.M.; Williams, B.A.; Mikkelsen, D.; Yu, W.; Gidley, M.J. Extracellular Depolymerisation Triggers Fermentation of Tamarind Xyloglucan and Wheat Arabinoxylan by a Porcine Faecal Inoculum. Carbohyd. Polym. 2018, 201, 575–582. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Zhao, J.B.; Tao, S.Y.; Zhou, X.J.; Pi, Y.; Gerrits, W.J.; Johnston, L.J.; Zhang, S.; Yang, H.; Liu, L.; et al. Effect of Dietary Fiber Fermentation on Short-chain fatty Acid Production and Microbial Composition In Vitro. J. Sci. Food Agric. 2020, 100, 4282–4291. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zuo, B.; Huang, S.; Zeng, B.; Han, D.; Li, T.; Liu, T.; Wu, Z.; Wei, H.; Zhao, J.; et al. Spatial Heterogeneity of Bacterial Colonization across Different Gut Segments following Inter-species Microbiota Transplantation. Microbiome 2020, 8, 161. [Google Scholar] [CrossRef]
- Menke, K.H. Estimation of the Energetic Feed Value Obtained from Chemical Analysis and In Vitro Gas Production using Rumen Fluid. Anim. Res. Dev. 1988, 28, 7–55. [Google Scholar]
- Zhang, D.F.; Yang, H.J. In Vitro Ruminal Methanogenesis of a Hay-rich Substrate in Response to Different Combination Supplements of Nitrocompounds; Pyromellitic Diimide and 2-bromoethanesulphonate. Anim. Feed Sci. Technol. 2011, 163, 20–32. [Google Scholar] [CrossRef]
- Groot, J.C.; Cone, J.W.; Williams, B.A.; Debersaques, F.M.; ALantinga, E.A. Multiphasic Analysis of Gas Production Kinetics for In Vitro Fermentation of Ruminant Feeds. Anim. Feed Sci. Technol. 1996, 64, 77–89. [Google Scholar] [CrossRef]
- Ibáñez, A.B.; Bauer, S. Downscaled Method using Glass Microfiber Filters for the Determination of Klason Lignin and Structural Carbohydrates. Biomass Bioenergy 2014, 68, 75–81. [Google Scholar] [CrossRef]
- Dunkley, K.D.; Dunkley, C.S.; Njongmeta, N.L.; Callaway, T.R.; Hume, M.E.; Kubena, L.F.; Nisbet, D.J.; Ricke, S.C. Comparison of In Vitro Fermentation and Molecular Microbial Profiles of High-fiber Feed Substrates Incubated with Chicken Cecal Inocula. Poult. Sci. 2007, 86, 801–810. [Google Scholar] [CrossRef]
- Li, N.; Huang, S.; Jiang, L.; Dai, Z.; Li, T.; Han, D.; Wang, J. Characterization of the Early Life Microbiota Development and Predominant Lactobacillus Species at Distinct Gut Segments of Low- and Normal-Birth-Weight Piglets. Front. Microbiol. 2019, 10, 797. [Google Scholar] [CrossRef] [PubMed]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end Assembler for Illumina Sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-throughput Community Sequencing Data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef] [PubMed]
- Bacci, G.; Bani, A.; Bazzicalupo, M.; Ceccherini, M.T.; Galardini, M.; Nannipieri, P.; Pietramellara, G.; Mengoni, A. Evaluation of the Performances of Ribosomal Database Project (RDP) Classifier for Taxonomic Assignment of 16S rRNA Metabarcoding Sequences Generated from Illumina-Solexa NGS. J. Genom. 2015, 3, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A Comprehensive Online Resource for Quality Checked and Aligned Ribosomal RNA Sequence Data Compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-source, Platform-independent, Community-supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Tiwari, U.P.; Singh, A.K.; Jha, R. Fermentation Characteristics of Resistant Starch, Arabinoxylan, and β-Glucan and their Effects on the Gut Microbial Ecology of Pigs: A Review. Anim. Nutr. 2019, 5, 217–226. [Google Scholar] [CrossRef]
- Verma, N.; Kumar, V. Impact of Process Parameters and Plant Polysaccharide Hydrolysates in Cellulase Production by Trichoderma Reesei and Neurospora Crassa under Wheat Bran based Solid State Fermentation. Biotechnol. Rep. 2020, 25, e00416. [Google Scholar] [CrossRef]
- Zhang, S.; Li, W.; Smith, C.J.; Musa, H. Cereal-derived Arabinoxylans as Biological Response Modifiers: Extraction, Molecular Features, and Immune-stimulating Properties. Crit. Rev. Food Sci. Nutr. 2015, 55, 1035–1052. [Google Scholar] [CrossRef]
- Carlson, J.L.; Erickson, J.M.; Hess, J.M.; Gould, T.J.; Slavin, J.L. Prebiotic Dietary Fiber and Gut Health: Comparing the in Vitro Fermentations of Beta-Glucan, Inulin and Xylooligosaccharide. Nutrients 2017, 9, 1361. [Google Scholar] [CrossRef] [PubMed]
- Kilua, A.; Nomata, R.; Nagata, R.; Fukuma, N.; Shimada, K.; Han, K.-H.; Fukushima, M. Purple Sweet Potato Polyphenols Differentially Influence the Microbial Composition Depending on the Fermentability of Dietary Fiber in a Mixed Culture of Swine Fecal Bacteria. Nutrients 2019, 11, 1495. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pang, B.; Yan, X.; Shang, X.; Hu, X.; Shi, J. Prebiotic Properties of Different Polysaccharide Fractions from Artemisia Sphaerocephala Krasch Seeds Evaluated by Simulated Digestion and In Vitro Fermentation by Human Fecal Microbiota. Int. J. Biol. Macromol. 2020, 162, 414–424. [Google Scholar] [CrossRef] [PubMed]
- Bergamaschi, M.; Tiezzi, F.; Howard, J.; Huang, Y.J.; Gray, K.A.; Schillebeeckx, C.; McNulty, N.P.; Maltecca, C. Gut Microbiome Composition Differences among Breeds Impact Feed Efficiency in Swine. Microbiome 2020, 8, 110. [Google Scholar] [CrossRef] [PubMed]
- Mannelli, F.; Cappucci, A.; Pini, F.; Pastorelli, R.; Decorosi, F.; Giovannetti, L.; Mele, M.; Minieri, S.; Conte, G.; Pauselli, M.; et al. Effect of Different Types of Olive Oil Pomace Dietary Supplementation on the Rumen Microbial Community Profile in Comisana ewes. Sci. Rep. 2018, 8, 8455. [Google Scholar] [CrossRef]
- Luo, J.; Chen, D.; Mao, X.; He, J.; Zeng, D. Purified β-glucans of Different Molecular Weights Enhance Growth Performance of LPS-challenged Piglets via Improved Gut Barrier Function and Microbiota. Animals 2019, 9, 602. [Google Scholar] [CrossRef]
- Shi, Y.; Liu, J.; Yan, Q.; You, X.; Yang, S.; Jiang, Z. In Vitro Digestibility and Prebiotic Potential of Curdlan (1→3)-β-d-glucan Oligosaccharides in Lactobacillus species. Carbohydr. Polym. 2018, 188, 17–26. [Google Scholar] [CrossRef]
- Lattimer, J.M.; Haub, M.D. Effects of Dietary Fiber and its Components on Metabolic Health. Nutrients 2010, 2, 1266–1289. [Google Scholar] [CrossRef]
- Oh, S.; Koike, S.; Kobayashi, Y. Effect of Ginkgo Extract Supplementation on In Vitro Rumen Fermentation and Bacterial Profiles under Different Dietary Conditions. Anim. Sci. J. 2017, 88, 1737–1743. [Google Scholar] [CrossRef]
- Pi, Y.; Hu, J.; Bai, Y.; Wang, Z.; Wu, Y.; Ye, H.; Zhang, S.; Tao, S.; Xiao, Y.; Han, D.; et al. Effects of Dietary Fibers with Different Physicochemical Properties on Fermentation Kinetics and Microbial Composition by Fecal Inoculum from Lactating Sows In Vitro. J. Sci. Food Agric. 2021, 101, 907–917. [Google Scholar] [CrossRef]
- Prins, R.A.; Lankhorst, A.; van der Meer, P.; Van Nevel, C.J. Some Characteristics of Anaerovibrio lipolytica a Rumen Lipolytic Organism. Antonie Leeuwenhoek 1975, 41, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Gong, J.; Liu, X.; Chen, C.; Sun, X.; Li, H.; Zhou, Y.; Cui, C.; Wang, Y.; Yang, Y.; et al. Gut Dysbiosis and Lack of Short Chain Fatty Acids in a Chinese Cohort of Patients with Multiple Sclerosis. Neurochem. Int. 2019, 129, 104468. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Dong, D.; Wang, C.; Cui, Y.; Jiang, C.; Ni, Q.; Su, T.; Wang, G.; Mao, E.; Peng, Y. Risk Factors and Intestinal Microbiota: Clostridioides Difficile Infection in Patients Receiving Enteral Nutrition at Intensive Care Units. Crit. Care 2020, 24, 426. [Google Scholar] [CrossRef] [PubMed]
- Schrempf, W.; Ziemssen, T. Glatiramer Acetate: Mechanisms of Action in Multiple Sclerosis. Autoimmun. Rev. 2007, 6, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Ariestanti, C.A.; Seechamnanturakit, V.; Harmayani, E.; Wichienchot, S. Optimization on Production of Konjac Oligo-glucomannan and their Effect on the Gut Microbiota. Food Sci. Nutr. 2019, 7, 788–796. [Google Scholar] [CrossRef] [PubMed]
- Bang, S.-J.; Kim, G.; Lim, M.Y.; Song, E.-J.; Jung, D.-H.; Kum, J.-S.; Nam, Y.-D.; Park, C.-S.; Seo, D.-H. The Influence of In Vitro Pectin Fermentation on the Human Fecal Microbiome. AMB Express 2018, 8, 98. [Google Scholar] [CrossRef] [PubMed]
- Heinritz, S.N.; Weiss, E.; Eklund, M.; Aumiller, T.; Heyer, C.M.; Messner, S.; Rings, A.; Louis, S.; Bischoff, S.C.; Mosenthin, R. Impact of a High-fat or High-fiber Diet on Intestinal Microbiota and Metabolic Markers in a Pig Model. Nutrients 2016, 8, 317. [Google Scholar] [CrossRef]
- Bommarius, A.S.; Katona, A.; Cheben, S.E.; Patel, A.S.; Ragauskas, A.J.; Knudson, K.; Pu, Y. Cellulase Kinetics as a Function of Cellulose Pretreatment. Metab. Eng. 2008, 10, 370–381. [Google Scholar] [CrossRef]
- Rong, W.; Han, K.; Zhao, Z.; An, J.; Li, Q.; Bi, K. The Protective Effect of Xanthoceras Sorbifolia Bunge Husks on Cognitive Disorder based on Metabolomics and Gut Microbiota Analysis. J. Ethnopharmacol. 2020, 113094. [Google Scholar] [CrossRef]
- Liu, B.; Wang, W.; Zhu, X.; Sun, X.; Xiao, J.; Li, D.; Cui, Y.; Wang, C.; Shi, Y. Response of Gut Microbiota to Dietary Fiber and Metabolic Interaction with SCFAs in Piglets. Front. Microbiol. 2018, 9, 2344. [Google Scholar] [CrossRef]
- Williams, B.A.; Mikkelsen, D.; Paih, L.L.; Gidley, M.J. In Vitro Fermentation Kinetics and End-products of Cereal Arabinoxylans and (1,3;1,4)-β-glucans by Porcine Faeces. J. Cereal Sci. 2011, 53, 53–58. [Google Scholar] [CrossRef]
- Ivarsson, E.; Roos, S.; Liu, Y.; Lindberg, E.J. Fermentable Non-starch Polysaccharides Increases the Abundance of Bacteroides-Prevotella-Porphyromonas in Ileal Microbial Community of Growing Pigs. Animal 2014, 8, 1777–1787. [Google Scholar] [CrossRef]
- Petersen, L.M.; Bautista, E.J.; Nguyen, H.; Hanson, B.M.; Chen, L.; Lek, S.H.; Sodergren, E.; Weinstock, G.M. Community Characteristics of the Gut Microbiomes of Competitive Cyclists. Microbiome 2017, 5, 98. [Google Scholar] [CrossRef]
- Hu, C.; Li, F.; Duan, Y.; Yin, Y.; Kong, X. Glutamic Acid Supplementation reduces Body Fat Weight in Finishing Pigs When Provided Solely or in Combination With Arginine and it is Associated with Colonic Propionate and Butyrate Concentrations. Food Funct. 2019, 10, 4693–4704. [Google Scholar] [CrossRef] [PubMed]
- Kovatcheva-Datchary, P.; Nilsson, A.; Akrami, R.; Lee, Y.S.; De Vadder, F.; Arora, T.; Hallen, A.; Martens, E.; Björck, I.; Bäckhed, F. Dietary Fiber-induced Improvement in Glucose Metabolism is Associated with Increased Abundance of Prevotella. Cell Metab. 2015, 22, 971–982. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Guo, C.; Sun, D.; Seddik, H.E.; Mao, S. The Ruminal Microbiome and Metabolome Alterations associated with Diet-induced Milk Fat Depression in Dairy Cows. Metabolites 2019, 9, 154. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Li, F.; Zhuang, Y.; Xu, J.; Wang, J.; Mao, X.; Zhang, Y.; Liu, X. Alteration in Gut Microbiota associated with Hepatitis B and Non-hepatitis Virus related Hepatocellular Carcinoma. Gut Pathog. 2019, 11, 1. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Yang, D.; Song, F.; Liu, M.; Dai, Y.; Zheng, F.; Yue, H. In Vitro Effects of Ginseng and the Seed of Zizyphus Jujuba Var. Spinosa on Gut Microbiota of Rats with Spleen Deficiency. Chem. Biodivers. 2020, 17, e2000199. [Google Scholar]
- Metzler-Zebeli, B.U.; Newman, M.A.; Grüll, D.; Zebeli, Q. Functional Adaptations in the Cecal and Colonic Metagenomes Associated with the Consumption of Transglycosylated Starch in a Pig Model. BMC Microbiol. 2019, 19, 87. [Google Scholar] [CrossRef]
- Sun, L.; Jia, H.; Li, J.; Yu, M.; Yang, Y.; Tian, D.; Zhang, H.; Zou, Z. Cecal Gut Microbiota and Metabolites might Contribute to the Severity of Acute Myocardial Ischemia by Impacting the Intestinal Permeability, Oxidative Stress, and Energy Metabolism. Front. Microbiol. 2019, 10, 1745. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polysaccharides | Glucose | Galactose | Mannose | Arabinose | Xylose |
|---|---|---|---|---|---|
| cellulose | 96.62 | 0.00 | 0.00 | 0.00 | 3.29 |
| arabinoxylan | 2.02 | 0.00 | 0.00 | 33.42 | 65.38 |
| β-glucan | 92.07 | 0.00 | 0.00 | 0.00 | 0.00 |
| glucomannan | 58.29 | 6.34 | 28.82 | 0.00 | 2.59 |
| Items | Con | Cel | Arx | β-Glu | Glm | p-Value |
|---|---|---|---|---|---|---|
| A, mL/g | 81.55 ± 8.95 c | 139.37 ± 3.74 b | 152.69 ± 3.89 a | 166.66 ± 3.71 a | 145.52 ± 3.74 a | <0.01 |
| B | 6.80 ± 1.09 d | 10.96 ± 0.26 b | 16.84 ± 0.78 a | 6.31 ± 0.19 d | 8.85 ± 0.56b c | <0.01 |
| C, h | 29.11 ± 0.36 a | 25.19 ± 0.46 b | 16.12 ± 0.15 d | 14.97 ± 0.27 d | 20.39 ± 0.38 c | <0.01 |
| RmaxG, mL/h | 27.65 ± 0.26 a | 24.77 ± 0.44 b | 16.01 ± 0.14 d | 14.22 ± 0.26 e | 19.86 ± 0.32 c | <0.01 |
| TRmaxG, h | 4.78 ± 0.61 c | 15.32 ± 0.71 b | 39.95 ± 1.79 a | 17.99 ± 0.24 b | 16.03 ± 1.29 b | <0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, Y.; Zhou, X.; Li, N.; Zhao, J.; Ye, H.; Zhang, S.; Yang, H.; Pi, Y.; Tao, S.; Han, D.; et al. In Vitro Fermentation Characteristics and Fiber-Degrading Enzyme Kinetics of Cellulose, Arabinoxylan, β-Glucan and Glucomannan by Pig Fecal Microbiota. Microorganisms 2021, 9, 1071. https://doi.org/10.3390/microorganisms9051071
Bai Y, Zhou X, Li N, Zhao J, Ye H, Zhang S, Yang H, Pi Y, Tao S, Han D, et al. In Vitro Fermentation Characteristics and Fiber-Degrading Enzyme Kinetics of Cellulose, Arabinoxylan, β-Glucan and Glucomannan by Pig Fecal Microbiota. Microorganisms. 2021; 9(5):1071. https://doi.org/10.3390/microorganisms9051071
Chicago/Turabian StyleBai, Yu, Xingjian Zhou, Na Li, Jinbiao Zhao, Hao Ye, Shiyi Zhang, Hongjian Yang, Yu Pi, Shiyu Tao, Dandan Han, and et al. 2021. "In Vitro Fermentation Characteristics and Fiber-Degrading Enzyme Kinetics of Cellulose, Arabinoxylan, β-Glucan and Glucomannan by Pig Fecal Microbiota" Microorganisms 9, no. 5: 1071. https://doi.org/10.3390/microorganisms9051071
APA StyleBai, Y., Zhou, X., Li, N., Zhao, J., Ye, H., Zhang, S., Yang, H., Pi, Y., Tao, S., Han, D., Zhang, S., & Wang, J. (2021). In Vitro Fermentation Characteristics and Fiber-Degrading Enzyme Kinetics of Cellulose, Arabinoxylan, β-Glucan and Glucomannan by Pig Fecal Microbiota. Microorganisms, 9(5), 1071. https://doi.org/10.3390/microorganisms9051071