
Genomic and Phenotypic Analysis of Linezolid-Resistant Staphylococcus epidermidis in a Tertiary Hospital in Innsbruck, Austria

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Isolates, Identification, and Antimicrobial Susceptibility Testing
2.2. DNA Extraction, Library Preparation, and Whole Genome Sequencing of Isolates
2.3. NGS Data Processing
2.4. Detection of Linezolid Resistance-Associated Genes and Mutations
2.5. Phylogenetic Computations/Multilocus Sequence Typing
2.6. Data Collection
2.7. Statistical Analysis
3. Results
3.1. Patients Characteristics
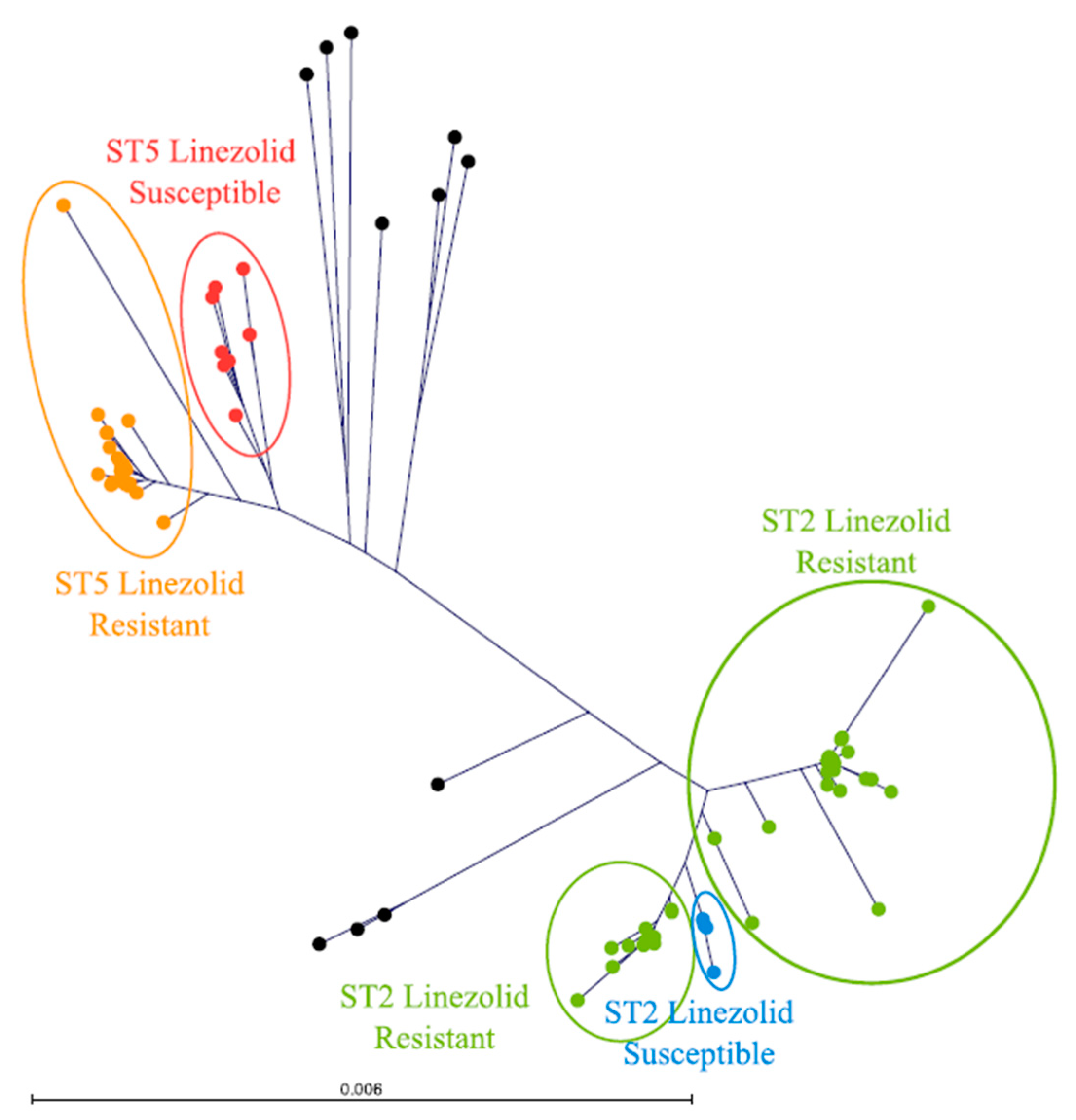
3.2. Mechanisms of Resistance to Linezolid Mainly Based on Mutations of the 23S rRNA in ST2 Isolates
3.3. Novel Variants in the 23S rRNA Linked to Linezolid Resistance
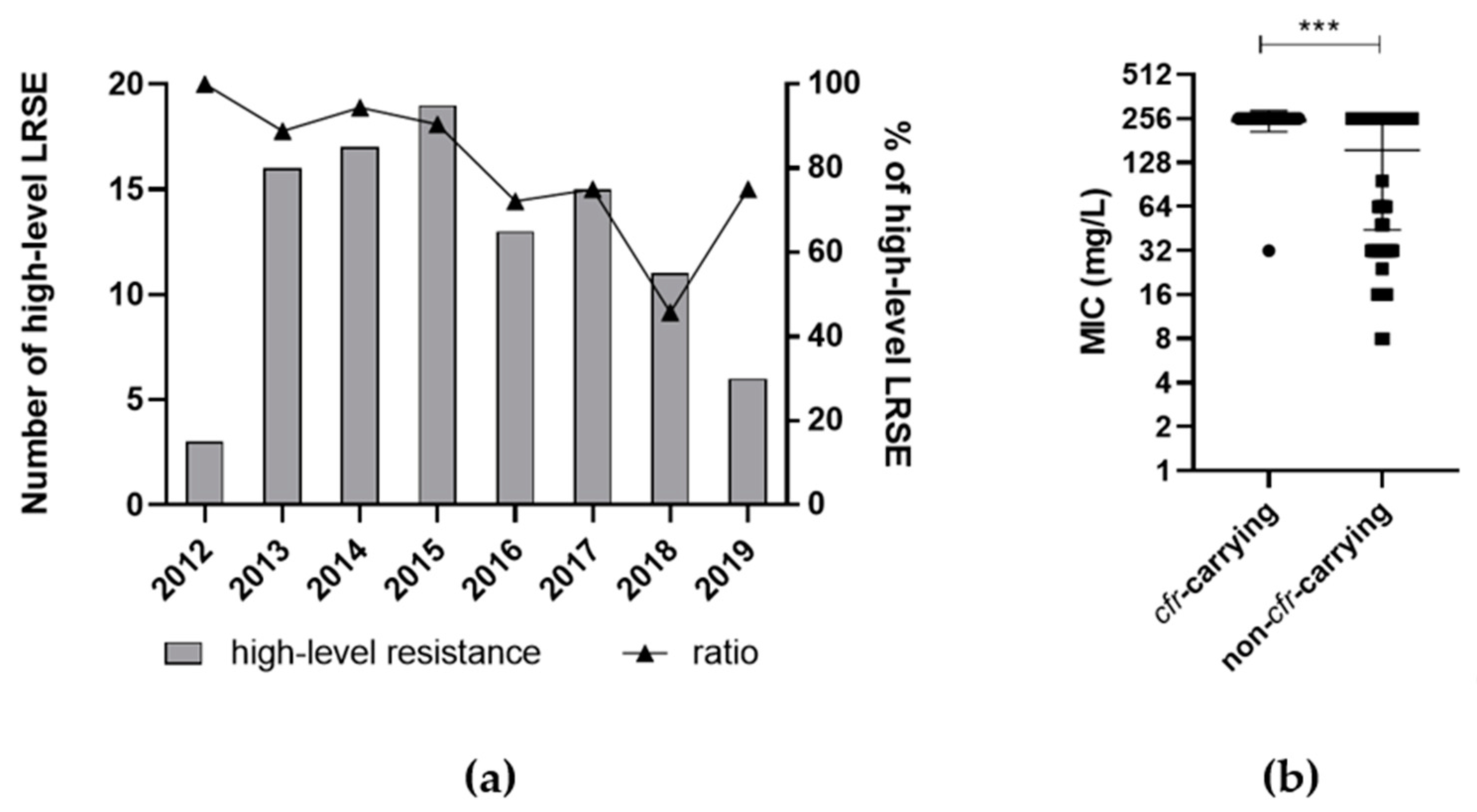
3.4. Phenotypic Linezolid Resistance Differs by Mechanism
3.5. Type of Resistance Mechanism or ST Was Not Dependent on Linezolid Use
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Otto, M. Staphylococcus epidermidis—The “accidental” pathogen. Nat. Rev. Microbiol. 2009, 7, 555–567. [Google Scholar] [CrossRef]
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-negative staphylococci. Clin. Microbiol. Rev. 2014, 27, 870–926. [Google Scholar] [CrossRef]
- Méric, G.; Miragaia, M.; de Been, M.; Yahara, K.; Pascoe, B.; Mageiros, L.; Mikhail, J.; Harris, L.G.; Wilkinson, T.S.; Rolo, J.; et al. Ecological Overlap and Horizontal Gene Transfer in Staphylococcus aureus and Staphylococcus epidermidis. Genome Biol. Evol. 2015, 7, 1313–1328. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.E.; Deshpande, L.M.; Jones, R.N. Linezolid update: Stable in vitro activity following more than a decade of clinical use and summary of associated resistance mechanisms. Drug Resist. Updat. Rev. Comment. Antimicrob. Anticancer Chemother. 2014, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nathwani, D.; Davey, P.G.; Marwick, C.A. MRSA: Treating people with infection. BMJ Clin. Evid. 2010, 10, 922. [Google Scholar]
- Swaney, S.M.; Aoki, H.; Ganoza, M.C.; Shinabarger, D.L. The oxazolidinone linezolid inhibits initiation of protein synthesis in bacteria. Antimicrob. Agents Chemother. 1998, 42, 3251–3255. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.E.; Deshpande, L.; Streit, J.M.; Sader, H.S.; Castanheira, M.; Hogan, P.A.; Flamm, R.K. ZAAPS programme results for 2016: An activity and spectrum analysis of linezolid using clinical isolates from medical centres in 42 countries. J. Antimicrob. Chemother. 2018, 73, 1880–1887. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Mendes, R.E.; Streit, J.M.; Hogan, P.A.; Flamm, R.K. Five-Year Summary of In Vitro Activity and Resistance Mechanisms of Linezolid against Clinically Important Gram-Positive Cocci in the United States from the LEADER Surveillance Program (2011 to 2015). Antimicrob. Agents Chemother. 2017, 61, e00609-17. [Google Scholar] [CrossRef] [PubMed]
- Eliopoulos, G.M.; Meka, V.G.; Gold, H.S. Antimicrobial Resistance to Linezolid. Clin. Infect. Dis. 2004, 39, 1010–1015. [Google Scholar] [CrossRef]
- Wong, A.; Reddy, S.P.; Smyth, D.S.; Aguero-Rosenfeld, M.E.; Sakoulas, G.; Robinson, D.A. Polyphyletic emergence of linezolid-resistant staphylococci in the United States. Antimicrob. Agents Chemother. 2010, 54, 742–748. [Google Scholar] [CrossRef]
- Zhu, W.; Tenover, F.C.; Limor, J.; Lonsway, D.; Prince, D.; Dunne, W.M.J.; Patel, J.B. Use of pyrosequencing to identify point mutations in domain V of 23S rRNA genes of linezolid-resistant Staphylococcus aureus and Staphylococcus epidermidis. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2007, 26, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Cidral, T.A.; Carvalho, M.C.; Figueiredo, A.M.S.; de Melo, M.C.N. Emergence of methicillin-resistant coagulase-negative staphylococci resistant to linezolid with rRNA gene C2190T and G2603T mutations. APMIS 2015, 123, 867–871. [Google Scholar] [CrossRef] [PubMed]
- Lincopan, N.; de Almeida, L.M.; de Araujo, M.E.; Mamizuka, E.M. Linezolid resistance in Staphylococcus epidermidis associated with a G2603T mutation in the 23S rRNA gene. Int. J. Antimicrob. Agents 2009, 34, 281–282. [Google Scholar] [CrossRef]
- Mendes, R.E.; Deshpande, L.M.; Farrell, D.J.; Spanu, T.; Fadda, G.; Jones, R.N. Assessment of linezolid resistance mechanisms among Staphylococcus epidermidis causing bacteraemia in Rome, Italy. J. Antimicrob. Chemother. 2010, 65, 2329–2335. [Google Scholar] [CrossRef] [PubMed]
- Escribano, I.; Rodríguez, J.C.; Llorca, B.; García-Pachon, E.; Ruiz, M.; Royo, G. Importance of the Efflux Pump Systems in the Resistance of Mycobacterium tuberculosis to Fluoroquinolones and Linezolid. Chemotherapy 2007, 53, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Mulanovich, V.E.; Huband, M.D.; McCurdy, S.P.; Lemmon, M.M.; Lescoe, M.; Jiang, Y.; Rolston, K.V.I.; LaSala, P.R. Emergence of linezolid-resistant coagulase-negative Staphylococcus in a cancer centre linked to increased linezolid utilization. J. Antimicrob. Chemother. 2010, 65, 2001–2004. [Google Scholar] [CrossRef] [PubMed]
- Bonilla, H.; Huband, M.D.; Seidel, J.; Schmidt, H.; Lescoe, M.; McCurdy, S.P.; Lemmon, M.M.; Brennan, L.A.; Tait-Kamradt, A.; Puzniak, L.; et al. Multicity Outbreak of Linezolid-Resistant Staphylococcus epidermidis Associated with Clonal Spread of a cfr-Containing Strain. Clin. Infect. Dis. 2010, 51, 796–800. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, W.; Wang, J.; Wu, C.; Shen, Z.; Fu, X.; Yan, Y.; Zhang, Q.; Schwarz, S.; Shen, J. Distribution of the multidrug resistance gene cfr in Staphylococcus species isolates from swine farms in China. Antimicrob. Agents Chemother. 2012, 56, 1485–1490. [Google Scholar] [CrossRef]
- Tewhey, R.; Gu, B.; Kelesidis, T.; Charlton, C.; Bobenchik, A.; Hindler, J.; Schork, N.J.; Humphries, R.M. Mechanisms of linezolid resistance among coagulase-negative staphylococci determined by whole-genome sequencing. MBio 2014, 5, e00894-14. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.; Strommenger, B.; Steglich, M.; Zimmermann, O.; Fenner, I.; Lensing, C.; Dagwadordsch, U.; Kekulé, A.S.; Werner, G.; Layer, F. Linezolid resistance in clinical isolates of Staphylococcus epidermidis from German hospitals and characterization of two cfr-carrying plasmids. J. Antimicrob. Chemother. 2015, 70, 1630–1638. [Google Scholar] [CrossRef] [PubMed]
- Dortet, L.; Glaser, P.; Kassis-Chikhani, N.; Girlich, D.; Ichai, P.; Boudon, M.; Samuel, D.; Creton, E.; Imanci, D.; Bonnin, R.; et al. Long-lasting successful dissemination of resistance to oxazolidinones in MDR Staphylococcus epidermidis clinical isolates in a tertiary care hospital in France. J. Antimicrob. Chemother. 2018, 73, 41–51. [Google Scholar] [CrossRef] [PubMed]
- EUCAST Disk Diffusion—Manual v 2.0 to v 7.0. Available online: https://www.eucast.org/ast_of_bacteria/previous_versions_of_documents/ (accessed on 21 January 2021).
- EUCAST Clinical Breakpoints—Bacteria v 2.0 to v 9.0. Available online: https://www.eucast.org/ast_of_bacteria/previous_versions_of_documents/ (accessed on 29 January 2021).
- Lepuschitz, S.; Weinmaier, T.; Mrazek, K.; Beisken, S.; Weinberger, J.; Posch, A.E. Analytical Performance Validation of Next-Generation Sequencing Based Clinical Microbiology Assays Using a K-mer Analysis Workflow. Front. Microbiol. 2020, 11, 1883. [Google Scholar] [CrossRef]
- Wood, D.E.; Lu, J.; Langmead, B. Improved metagenomic analysis with Kraken 2. Genome Biol. 2019, 20, 257. [Google Scholar] [CrossRef]
- Ferreira, I.; Beisken, S.; Lueftinger, L.; Weinmaier, T.; Klein, M.; Bacher, J.; Patel, R.; von Haeseler, A.; Posch, A.E. Species Identification and Antibiotic Resistance Prediction by Analysis of Whole-Genome Sequence Data by Use of ARESdb: An Analysis of Isolates from the Unyvero Lower Respiratory Tract Infection Trial. J. Clin. Microbiol. 2020, 58, e00273-20. [Google Scholar] [CrossRef] [PubMed]
- Meka, V.G.; Gold, H.S.; Cooke, A.; Venkataraman, L.; Eliopoulos, G.M.; Moellering Jr, R.C.; Jenkins, S.G. Reversion to susceptibility in a linezolid-resistant clinical isolate of Staphylococcus aureus. J. Antimicrob. Chemother. 2004, 54, 818–820. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed]
- Broad Institute. GitHub Repository Picard Toolkit. Available online: http://broadinstitute.github.io/picard/ (accessed on 11 May 2020).
- Li, H. A statistical framework for SNP calling, mutation discovery, association mapping and population genetical parameter estimation from sequencing data. Bioinformatics 2011, 27, 2987–2993. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. mlst Github. Available online: https://github.com/tseemann/mlst (accessed on 1 May 2020).
- Katz, L.; Griswold, T.; Morrison, S.; Caravas, J.; Zhang, S.; Bakker, H.; Deng, X.; Carleton, H. Mashtree: A rapid comparison of whole genome sequence files. J. Open Source Softw. 2019, 4, 1762. [Google Scholar] [CrossRef]
- Huber, S.; Weinberger, J.; Pilecky, M.; Lorenz, I.; Schildberger, A.; Weber, V.; Fuchs, S.; Posch, W.; Knabl, L.; Würzner, R.; et al. A high leukocyte count and administration of hydrocortisone hamper PCR-based diagnostics for bloodstream infections. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2021. [Google Scholar] [CrossRef]
- GLASS Whole-Genome Sequencing for Surveillance of Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2020; License: CC BY-NC-SA 3.0 IGO.
- Endimiani, A.; Blackford, M.; Dasenbrook, E.C.; Reed, M.D.; Bajaksouszian, S.; Hujer, A.M.; Rudin, S.D.; Hujer, K.M.; Perreten, V.; Rice, L.B.; et al. Emergence of linezolid-resistant Staphylococcus aureus after prolonged treatment of cystic fibrosis patients in Cleveland, Ohio. Antimicrob. Agents Chemother. 2011, 55, 1684–1692. [Google Scholar] [CrossRef] [PubMed]
- Locke, J.B.; Hilgers, M.; Shaw, K.J. Novel ribosomal mutations in Staphylococcus aureus strains identified through selection with the oxazolidinones linezolid and torezolid (TR-700). Antimicrob. Agents Chemother. 2009, 53, 5265–5274. [Google Scholar] [CrossRef] [PubMed]
- Morroni, G.; Brenciani, A.; Vincenzi, C.; Barocci, S.; Tili, E.; Manso, E.; Mingoia, M.; Menzo, S.; Varaldo, P.E.; Giovanetti, E. A clone of linezolid-resistant Staphylococcus epidermidis bearing the G2576T mutation is endemic in an Italian hospital. J. Hosp. Infect. 2016, 94, 203–206. [Google Scholar] [CrossRef]
- Ishiwada, N.; Takaya, A.; Kimura, A.; Watanabe, M.; Hino, M.; Ochiai, H.; Matsui, M.; Shibayama, K.; Yamamoto, T. Linezolid-resistant Staphylococcus epidermidis associated with long-term, repeated linezolid use in a pediatric patient. J. Infect. Chemother. Off. J. Japan Soc. Chemother. 2016, 22, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Musumeci, R.; Calaresu, E.; Gerosa, J.; Oggioni, D.; Bramati, S.; Morelli, P.; Mura, I.; Piana, A.; Are, B.M.; Cocuzza, C.E. Resistance to linezolid in Staphylococcus spp. clinical isolates associated with ribosomal binding site modifications: Novel mutation in domain V of 23S rRNA. New Microbiol. 2016, 39, 269–273. [Google Scholar] [PubMed]
- Li, X.; Arias, C.A.; Aitken, S.L.; Galloway Peña, J.; Panesso, D.; Chang, M.; Diaz, L.; Rios, R.; Numan, Y.; Ghaoui, S.; et al. Clonal Emergence of Invasive Multidrug-Resistant Staphylococcus epidermidis Deconvoluted via a Combination of Whole-Genome Sequencing and Microbiome Analyses. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2018, 67, 398–406. [Google Scholar] [CrossRef]
- Bouiller, K.; Ilic, D.; Wicky, P.H.; Cholley, P.; Chirouze, C.; Bertrand, X. Spread of clonal linezolid-resistant Staphylococcus epidermidis in an intensive care unit associated with linezolid exposure. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2020, 39, 1271–1277. [Google Scholar] [CrossRef] [PubMed]
- Treviño, M.; Martínez-Lamas, L.; Romero-Jung, P.A.; Giráldez, J.M.; Alvarez-Escudero, J.; Regueiro, B.J. Endemic linezolid-resistant Staphylococcus epidermidis in a critical care unit. Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 527–533. [Google Scholar] [CrossRef]
- Liakopoulos, A.; Spiliopoulou, I.; Damani, A.; Kanellopoulou, M.; Schoina, S.; Papafragas, E.; Marangos, M.; Fligou, F.; Zakynthinos, E.; Makris, D.; et al. Dissemination of two international linezolid-resistant Staphylococcus epidermidis clones in Greek hospitals. J. Antimicrob. Chemother. 2010, 65, 1070–1071. [Google Scholar] [CrossRef]
- Bouiller, K.; Bador, J.; Bruyère, R.; Toitot, A.; Prin, S.; Quenot, J.-P.; Charles, P.-E. Recent exposure to linezolid is strongly associated with the isolation of linezolid-resistant coagulase-negative staphylococci in patients with related infection or colonisation: A case–control study in an intensive care unit. Int. J. Antimicrob. Agents 2017, 50, 693–694. [Google Scholar] [CrossRef]
- Kelly, S.; Collins, J.; Maguire, M.; Gowing, C.; Flanagan, M.; Donnelly, M.; Murphy, P.G. An outbreak of colonization with linezolid-resistant Staphylococcus epidermidis in an intensive therapy unit. J. Antimicrob. Chemother. 2008, 61, 901–907. [Google Scholar] [CrossRef] [PubMed]
- Seral, C.; Sáenz, Y.; Algarate, S.; Duran, E.; Luque, P.; Torres, C.; Castillo, F.J. Nosocomial outbreak of methicillin- and linezolid-resistant Staphylococcus epidermidis associated with catheter-related infections in intensive care unit patients. Int. J. Med. Microbiol. 2011, 301, 354–358. [Google Scholar] [CrossRef]
- Russo, A.; Campanile, F.; Falcone, M.; Tascini, C.; Bassetti, M.; Goldoni, P.; Trancassini, M.; Della Siega, P.; Menichetti, F.; Stefani, S.; et al. Linezolid-resistant staphylococcal bacteraemia: A multicentre case-case-control study in Italy. Int. J. Antimicrob. Agents 2015, 45, 255–261. [Google Scholar] [CrossRef]
- Besier, S.; Ludwig, A.; Zander, J.; Brade, V.; Wichelhaus, T.A. Linezolid resistance in Staphylococcus aureus: Gene dosage effect, stability, fitness costs, and cross-resistances. Antimicrob. Agents Chemother. 2008, 52, 1570–1572. [Google Scholar] [CrossRef]
- LaMarre, J.; Mendes, R.E.; Szal, T.; Schwarz, S.; Jones, R.N.; Mankin, A.S. The genetic environment of the cfr gene and the presence of other mechanisms account for the very high linezolid resistance of Staphylococcus epidermidis isolate 426-3147L. Antimicrob. Agents Chemother. 2013, 57, 1173–1179. [Google Scholar] [CrossRef]
- Shen, J.; Wang, Y.; Schwarz, S. Presence and dissemination of the multiresistance gene cfr in Gram-positive and Gram-negative bacteria. J. Antimicrob. Chemother. 2013, 68, 1697–1706. [Google Scholar] [CrossRef] [PubMed]
- Locke, J.B.; Morales, G.; Hilgers, M.; Kedar, G.C.; Rahawi, S.; Picazo, J.J.; Shaw, K.J.; Stein, J.L. Elevated linezolid resistance in clinical cfr-positive Staphylococcus aureus isolates is associated with co-occurring mutations in ribosomal protein L3. Antimicrob. Agents Chemother. 2010, 54, 5352–5355. [Google Scholar] [CrossRef] [PubMed]
- Baos, E.; Candel, F.J.; Merino, P.; Pena, I.; Picazo, J.J. Characterization and monitoring of linezolid-resistant clinical isolates of Staphylococcus epidermidis in an intensive care unit 4 years after an outbreak of infection by cfr-mediated linezolid-resistant Staphylococcus aureus. Diagn. Microbiol. Infect. Dis. 2013, 76, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Ruiz de Gopegui, E.; Juan, C.; Zamorano, L.; Pérez, J.L.; Oliver, A. Transferable multidrug resistance plasmid carrying cfr associated with tet (L), ant (4′)-Ia, and dfrK genes from a clinical methicillin-resistant Staphylococcus aureus ST125 strain. Antimicrob. Agents Chemother. 2012, 56, 2139–2142. [Google Scholar] [CrossRef]
- Song, Y.; Lv, Y.; Cui, L.; Li, Y.; Ke, Q.; Zhao, Y. cfr-mediated linezolid-resistant clinical isolates of methicillin-resistant coagulase-negative staphylococci from China. J. Glob. Antimicrob. Resist. 2017, 8, 1–5. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Isolate Count (n) per Mutation | ||||
|---|---|---|---|---|
| 23S G2603U | L3 H146Q | cfr | EmrE | n (%) |
| x | 58 (45.0) | |||
| x | 25 (18.6) | |||
| x | x | 28 (21.7) | ||
| x | x | 4 (3.1) | ||
| x | x | 1 (0.8) | ||
| x | x | 2 (1.6) | ||
| 12 (9.3) * | ||||
| 91 (70.5) | 30 (23.3) | 30 (23.3) | 1 (0.8) | 129 |
| Level of Linezolid Resistance | MIC (mg/L) | Number of LRSE Isolates |
|---|---|---|
| high | ≥256 | 83 (64.3) |
| 96 | 1 (0.8) | |
| 64 | 8 (6.2) | |
| 48 | 3 (2.3) | |
| low | 32 | 21 (16.3) |
| 24 | 1 (0.8) | |
| 16 | 9 (7.0) | |
| 8 | 3 (2.3) | |
| Total | 129 (100.0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huber, S.; Knoll, M.A.; Berktold, M.; Würzner, R.; Brindlmayer, A.; Weber, V.; Posch, A.E.; Mrazek, K.; Lepuschitz, S.; Ante, M.; et al. Genomic and Phenotypic Analysis of Linezolid-Resistant Staphylococcus epidermidis in a Tertiary Hospital in Innsbruck, Austria. Microorganisms 2021, 9, 1023. https://doi.org/10.3390/microorganisms9051023
Huber S, Knoll MA, Berktold M, Würzner R, Brindlmayer A, Weber V, Posch AE, Mrazek K, Lepuschitz S, Ante M, et al. Genomic and Phenotypic Analysis of Linezolid-Resistant Staphylococcus epidermidis in a Tertiary Hospital in Innsbruck, Austria. Microorganisms. 2021; 9(5):1023. https://doi.org/10.3390/microorganisms9051023
Chicago/Turabian StyleHuber, Silke, Miriam A. Knoll, Michael Berktold, Reinhard Würzner, Anita Brindlmayer, Viktoria Weber, Andreas E. Posch, Katharina Mrazek, Sarah Lepuschitz, Michael Ante, and et al. 2021. "Genomic and Phenotypic Analysis of Linezolid-Resistant Staphylococcus epidermidis in a Tertiary Hospital in Innsbruck, Austria" Microorganisms 9, no. 5: 1023. https://doi.org/10.3390/microorganisms9051023
APA StyleHuber, S., Knoll, M. A., Berktold, M., Würzner, R., Brindlmayer, A., Weber, V., Posch, A. E., Mrazek, K., Lepuschitz, S., Ante, M., Beisken, S., Orth-Höller, D., & Weinberger, J. (2021). Genomic and Phenotypic Analysis of Linezolid-Resistant Staphylococcus epidermidis in a Tertiary Hospital in Innsbruck, Austria. Microorganisms, 9(5), 1023. https://doi.org/10.3390/microorganisms9051023