
Metagenomic Insight: Dietary Thiamine Supplementation Promoted the Growth of Carbohydrate-Associated Microorganisms and Enzymes in the Rumen of Saanen Goats Fed High-Concentrate Diets

Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Sample Collection
2.3. Total DNA Extraction, Library Construction, and Metagenomics Sequencing
2.4. Sequence Quality Control and Genome Assembly
2.5. Gene Prediction and Functional Database Annotations
2.6. Statistical Analysis
3. Results
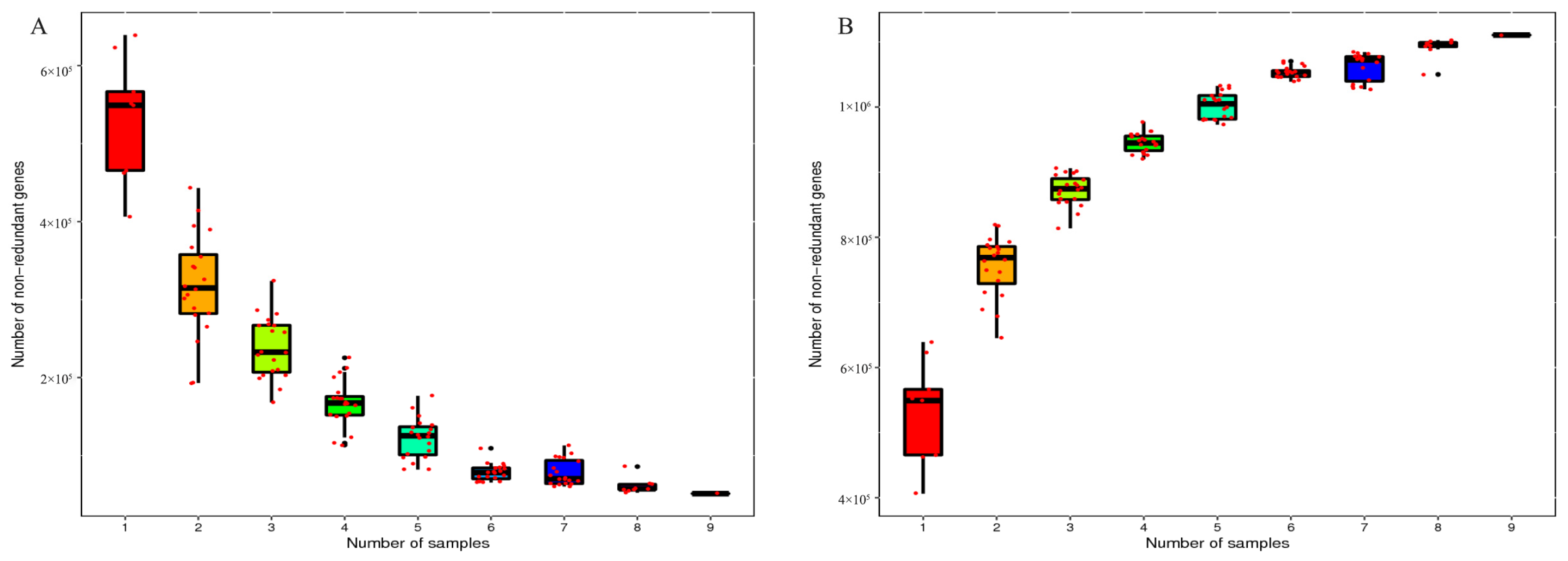
3.1. Sequencing Information
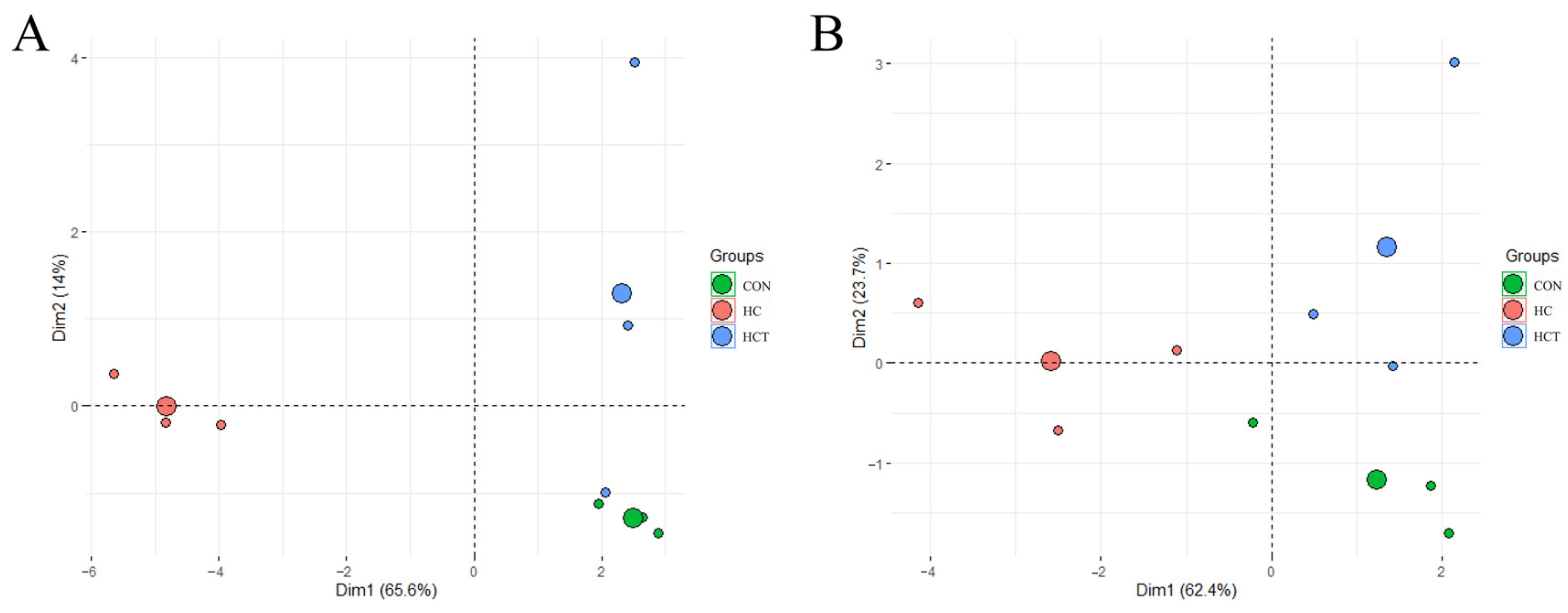
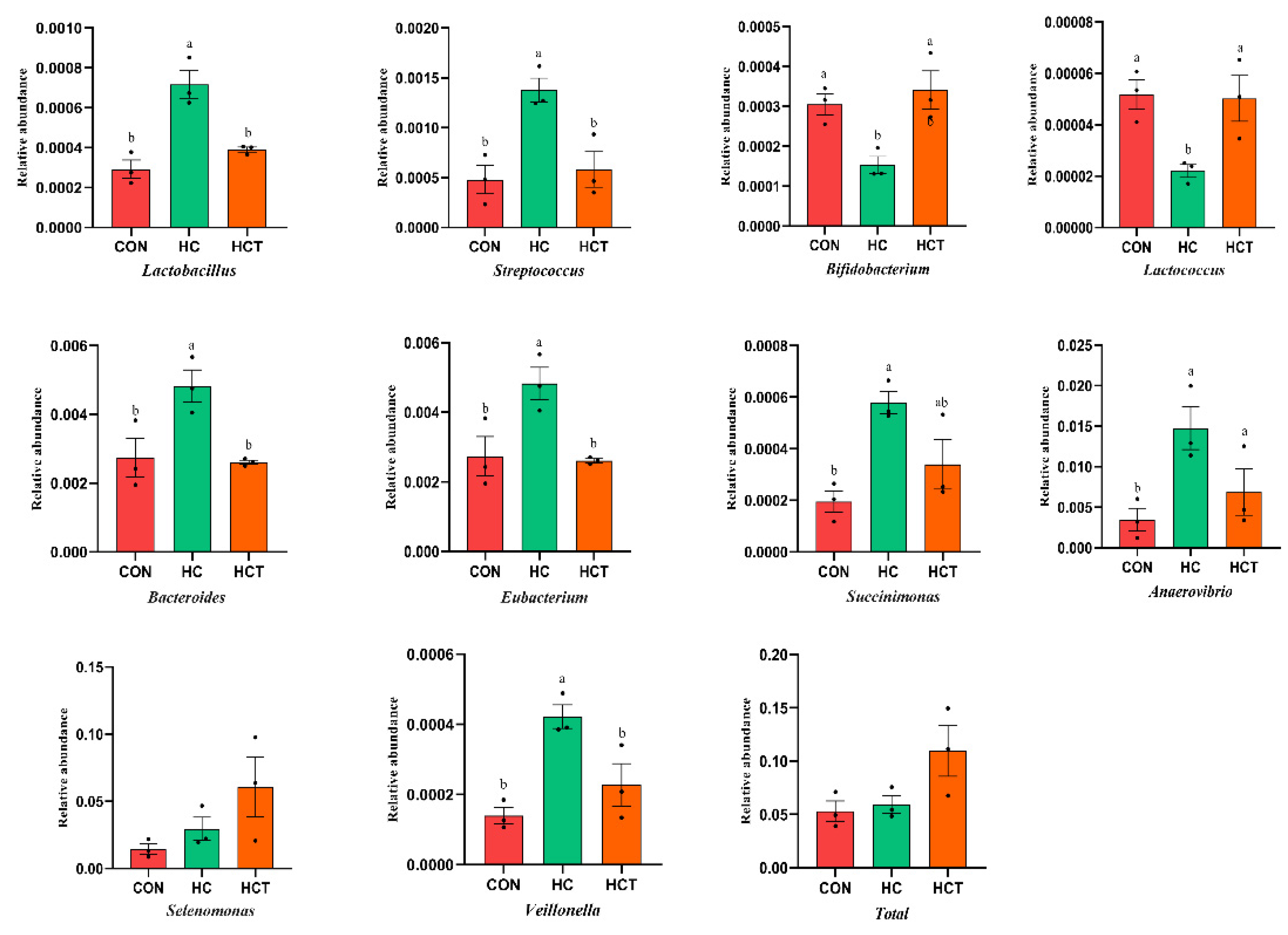
3.2. Effects of Thiamine Supplementation on the Profiles of Carbohydrate-Related Microorganisms and Enzymes
3.3. Effects of Thiamine Supplementation on Carbohydrate-Related Microorganisms Related Fiber and Starch Degradation
3.4. Effects of Thiamine Supplementation on Fiber-Degrading Enzymes and Starch Degrading Enzymes
3.5. Relationships between CAZymes and Animal Performance
4. Discussion
4.1. Effects of Thiamine Supplementation on Fiber-Degrading Microorganisms and Enzymes
4.2. Effects of Thiamine Supplementation on Starch-Degrading Microorganisms and Enzymes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bergman, E.N. Energy contributions of volatile fatty acids from the gastrointestinal tract in various species. Physiol. Rev. 1990, 70, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Huo, W.J.; Mao, S.Y.; Zhu, W.Y. Microbiome-metabolome analysis reveals unhealthy alterations in the composition and metabolism of ruminal microbiota with increasing dietary grain in a goat model. Environ. Microbiol. 2016, 18, 525–541. [Google Scholar] [CrossRef]
- Wang, H.R.; Pan, X.H.; Wang, C.; Wang, M.; Yu, L. Effects of different dietary concentrate to forage ratio and supplementation on the rumen fermentation and ruminal bacterial community in dairy cows. Anim. Prod. Sci. 2014, 55, 189–193. [Google Scholar] [CrossRef]
- Mccann, J.C.; Luan, S.; Cardoso, F.C.; Hooman, D.; Ehsan, K.; Loor, J.J. Induction of subacute ruminal acidosis affects the ruminal microbiome and epithelium. Front. Microbiol. 2016, 7, 701. [Google Scholar] [CrossRef] [PubMed]
- Kleen, J.L.; Hooijer, G.A.; Rehage, J.; Noordhuizen, J. Subacute ruminal acidosis in dairy cows: Ministry Agriculture Food and Rural Affairs. J. Vet. Med. 2003, 50, 30–31. [Google Scholar] [CrossRef]
- Zhang, H.; Peng, A.L.; Zhao, F.F.; Yu, L.H.; Wang, H.R. Thiamine ameliorates inflammation of the ruminal epithelium of Saanen goats suffering from subacute ruminal acidosis. J. Dairy Sci. 2019, 103, 1931–1943. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Xue, F.; Nan, X.; Tang, Z.; Wang, K.; Beckers, Y.; Jiang, L.; Xiong, B. Illumina sequencing approach to characterize thiamine metabolism related bacteria and the impacts of thiamine supplementation on ruminal microbiota in dairy cows fed high-grain diets. Front. Microbiol. 2017, 8, 1818. [Google Scholar] [CrossRef] [PubMed]
- Subramanya, S.B.; Subramanian, V.S.; Said, H.M. Chronic alcohol consumption and intestinal thiamin absorption: Effects on physiological and molecular parameters of the uptake process. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, 23–31. [Google Scholar] [CrossRef]
- Pan, X.H.; Nan, X.M.; Yang, L.; Jiang, L.; Xiong, B.H. Thiamine status, metabolism and application in dairy cows: A review. Br. J. Nurt. 2018, 120, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Karapinar, T.; Dabak, M.; Kizil, O. Thiamine status of feedlot cattle fed a high-concentrate diet. Can. Vet. J. 2010, 51, 1251–1253. [Google Scholar] [CrossRef]
- Brent, B.E. Relationship of acidosis to other feedlot ailments. J. Anim. Sci. 1976, 43, 930–935. [Google Scholar] [CrossRef]
- Xue, F.; Nan, X.; Sun, F.; Pan, X.; Guo, Y.; Jiang, L.; Xiong, B. Metagenome sequencing to analyze the impacts of thiamine supplementation on ruminal fungi in dairy cows fed high-concentrate diets. AMB Express 2018, 159, 1–12. [Google Scholar] [CrossRef]
- Xue, F.; Nan, X.; Li, Y.; Pan, X.; Guo, Y.; Jiang, L.; Xiong, B. Metagenomic insights into effects of thiamine supplementation on ruminal nonmethanogen archaea in high-concentrate diets feeding dairy cows. BMC Vet. Res. 2019, 15, 1–11. [Google Scholar] [CrossRef]
- Qin, N.; Yang, F.; Li, A. Alterations of the human gut microbiome in liver cirrhosis. Nature 2014, 513, 59–64. [Google Scholar] [CrossRef]
- Mende, D.R.; Waller, A.S.; Sunagawa, S. Assessment of metagenomic assembly using simulated next generation sequencing data. PLoS ONE 2012, 7, e31386. [Google Scholar] [CrossRef]
- Nielsen, H.B.; Almeida, M.; Juncker, A.S. Identification and assembly of genomes and genetic elements in complex metagenomic samples without using reference genomes. Nat. Biotechnol. 2014, 32, 822–828. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Z.; Godzik, A. Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 2006, 22, 1658–1659. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.M.; Niu, B.F.; Zhu, Z.W.; Wu, S.T.; Li, W.Z. CD-HIT: Accelerated for clustering the next-generation sequencing data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef]
- Li, J.; Jia, H.; Cai, X.; Zhong, H.; Feng, Q.; Sunagawa, S.; Arumugam, M.; Kultima, J.R.; Prifti, E.; Nielsen, T.; et al. An integrated catalog of reference genes in the human gut microbiome. Nat. Biotechnol. 2014, 32, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, Y.; Cai, Z.; Li, S.; Zhu, J.; Zhang, F.; Liang, S.; Zhang, W.; Guan, Y.; Shen, D.; et al. A metagenome-wide association study of gut microbiota in type 2 diabetes. Nature 2012, 490, 55–60. [Google Scholar] [CrossRef]
- Cantarel, B.L.; Coutinho, P.M.; Rancurel, C.; Bernard, T.; Lombard, V.; Henrissat, B. The Carbohydrate-Active EnZymes database (CAZy): An expert resource for Glycogenomics. Nucleic Acids Res. 2009, 37, 233–238. [Google Scholar] [CrossRef]
- Feng, Q.; Liang, S.; Jia, H.; Stadlmayr, A.; Tang, L.; Lan, Z.; Zhang, D.; Xia, H.; Xu, X.; Jie, Z.; et al. Gut microbiome development along the colorectal adenoma carcinoma sequence. Nat. Commun. 2015, 6, 6528. [Google Scholar] [CrossRef]
- Bäckhed, F.; Roswall, J.; Peng, Y.; Feng, Q.; Jia, H.; Kovatcheva-Datchary, P.; Li, Y.; Xia, Y.; Xie, H.; Zhong, H.; et al. Dynamics and Stabilization of the Human Gut Microbiome during the First Year of Life. Cell Host Microbe 2015, 17, 690–703. [Google Scholar] [CrossRef] [PubMed]
- Krause, D.O.; Denman, S.E.; Mackie, R.I.; Morrison, M.; Rae, A.L.; Attwood, G.T.; McSweeney, C.S. Opportunities to improve fiber degradation in the rumen:microbiology, ecology, and genomics. FEMS Microbiol. Rev. 2003, 27, 663–693. [Google Scholar] [CrossRef]
- Puniya, A.K.; Singh, R.; Kamra, D.N. Rumen Microbiology: From Evolution to Revolution; Springer Press: New York, NY, USA, 2015; pp. 97–109. ISBN 978-81-322-2400-6. [Google Scholar] [CrossRef]
- Xia, Y.; Kong, Y.; Seviour, R.; Yang, H.E.; Forster, R.; Vasanthan, T.; McAllister, T. In situ identification and quantification of starch-hydrolyzing bacteria attached to barley and corn grain in the rumen of cows fed barley-based diets. FEMS Microbiol. Ecol. 2015, 91, fiv077. [Google Scholar] [CrossRef][Green Version]
- Comtet-Marre, S.; Parisot, N.; Lepercq, P.; Chaucheyras-Durand, F.; Mosoni, P.; Peyretaillade, E.; Bayat, A.R.; Shingfield, K.J.; Peyret, P.; Forano, E. Metatranscriptomics reveals the active bacterial and Eukaryotic Fibrolytic communities in the rumen of dairy cow fed a mixed diet. Front. Microbiol. 2017, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Akin, D.E.; Borneman, W.S. Role of Rumen Fungi in Fiber Degradation. J. Dairy Sci. 1990, 73, 3023–3032. [Google Scholar] [CrossRef]
- Hespell, R.B.; Akin, D.E.; Dehority, B.A. Bacteria, fungi, and protozoa of the rumen. In Gastrointestinal Microbiology; Mackie, R.I., White, B.A., Isaacson, R.E., Eds.; Chapman and Hall: New York, NY, USA, 1997; pp. 59–141. [Google Scholar]
- Cho, K.K.; Kim, S.C.; Woo, J.H.; Bok, J.D.; Choi, Y.J. Molecular cloning and expression of a novel family A endoglucanase gene from Fibrobacter succinogenes S85 in Escherichia coli. Enzyme Microb. Technol. 2000, 27, 475–481. [Google Scholar] [CrossRef]
- Ohara, H.; Karita, S.; Kimura, T.; Sakka, K.; Ohmiya, K. Characterization of the cellulolytic complex (cellulosome) from Ru-minococcus albus. Biosci. Biotechnol. Biochem. 2000, 64, 254–260. [Google Scholar] [CrossRef]
- Forsberg, C.W.; Cheng, K.J.; White, B.A. Polysaccharide degradation in the rumen and large intestine. In Gastrointestinal Microbiology; Mackie, R.I., White, B.A., Eds.; Chapman and Hall: New York, NY, USA, 1997; pp. 319–379. [Google Scholar]
- Goad, D.W.; Goad, C.L.; Nagaraja, T.G. Ruminal microbial and fermentative changes associated with experimentally induced subacute acidosis in steers. J. Anim. Sci. 1998, 76, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.Y.; Liu, J.H.; Jiang, L.S.; Mao, S.Y. Effect of high-concentrate diets on microbial composition, function, and the VFAs formation process in the rumen of dairy cows. Anim. Feed Sci. Tech. 2020, 269, 114619. [Google Scholar] [CrossRef]
- Bainbridge, M.L.; Cersosimo, L.M.; Wright, A.D.; Kraft, J. Rumen bacterial communities shift across a lactation in Holstein, Jersey and Holstein x Jersey dairy cows and correlate torumen function, bacterial fatty acid composition and production parameters. FEMS Microbiol. Ecol. 2016, 92, fiw059. [Google Scholar] [CrossRef]
- Flint, H.J.; Bayer, E.A.; Rincon, M.T.; Lamed, R.; White, B.A. Polysaccharide utilization by gut bacteria: Potential for new insights from genomic analysis. Nat. Rev. Microbiol. 2008, 6, 121–131. [Google Scholar] [CrossRef]
- Tapio, I.; Fischer, D.; Blasco, L.; Tapio, M.; Wallace, R.J.; Bayat, A.R.; Ventto, L.; Kahala, M.; Negussie, E.; Shingfield, K.J.; et al. Taxon abundance, diversity, cooccurrence and network analysis of the ruminal microbiota in response to dietary changes in dairy cows. PLoS ONE 2017, 12, e0180260. [Google Scholar] [CrossRef]
- Haitjema, C.H.; Gilmore, S.P.; Henske, J.K.; Solomon, K.V.; De Groot, R.; Kuo, A.; Mondo, S.J.; Salamov, A.A.; LaButti, K.; Zhao, Z.; et al. A parts list for fungal cellulosomes revealed by comparative genomics. Nat. Microbiol. 2017, 2, 17087. [Google Scholar] [CrossRef]
- Shimizu, M.; Masuo, S.; Itoh, E.; Zhou, S.; Kato, M.; Takaya, N. Thiamine synthesis regulates the fermentation mechanisms in the fungus Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2016, 80, 1768–1775. [Google Scholar] [CrossRef]
- Xue, F.; Pan, X.; Jiang, L.; Guo, Y.; Xiong, B. GC-MS analysis of the ruminal metabolome response to thiamine supplementation during high grain feeding in dairy cows. Metabolomics 2018, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hook, S.E.; Steele, M.A.; Northwood, K.S. Impact of subacute ruminal acidosis (SARA) adaptation and recovery on the density and diversity of bacteria in the rumen of dairy cows. FEMS Microbiol. Ecol. 2011, 78, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Vihinen, M.; Mantsala, P. Microbial amylolytic enzymes. Crit. Rev. Biochem. Mol. Biol. 1989, 24, 329–418. [Google Scholar] [CrossRef] [PubMed]
- Hamlin, L.J.; Hungate, R.E. Culture and physiology of a starch-digesting bacterium (Bacteroides amylophilus n. sp.) from the bovine rumen. J. Bacteriol. 1956, 72, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Mackie, R.I.; Gilchrist, F.M. Changes in Lactate-Producing and Lactate-Utilizing Bacteria in Relation to pH in the Rumen of Sheep During Stepwise Adaptation to a High-Concentrate Diet. Appl. Environ. Microbiol. 1979, 38, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Nagaraja, T.G.; Titgemeyer, E.C. Ruminal acidosis in beef cattle: The current microbiological and nutritional outlook. J. Dairy Sci. 2007, 1, E17–E38. [Google Scholar] [CrossRef] [PubMed]
- Tajima, K.; Aminov, R.I.; Nagamine, T.; Matsui, H.; Nakamura, M.; Benno, Y. Diet-dependent shifts in the bacterial population of the rumen revealed with real-time PCR. Appl. Environ. Microbiol. 2001, 67, 2766–2774. [Google Scholar] [CrossRef]
- Zhao, Y.; Xue, F.; Hua, D.; Wang, Y.; Pan, X.; Nan, X.; Sun, F.; Jiang, L.; Xiong, B. Metagenomic insights into effects of thiamine supplementation on carbohydrate-active enzymes’ profile in dairy cows fed high-concentrate diets. Animals 2020, 10, 304. [Google Scholar] [CrossRef] [PubMed]
- Elizabeth, F.B.; Alisdair, B.B. Insights into the recognition of the human glycome by microbial carbohydrate-binding modules. Curr. Opin. Struct. Biol. 2012, 22, 570–577. [Google Scholar] [CrossRef]
- Abbott, D.W.; van Bueren, A.L. Using structure to inform carbohydrate binding module function. Curr. Opin. Struct. Biol. 2014, 28, 32–40. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | CON | HC | HCT |
|---|---|---|---|
| Ingredient (% of DM) | |||
| Chinese wildrye hay | 70.00 | 30.00 | 30.00 |
| Corn grain | 14.00 | 58.90 | 58.90 |
| Soybean meal | 13.00 | 8.45 | 8.45 |
| Calcium hydrophosphate | 1.42 | 0.53 | 0.53 |
| Limestone | 0.58 | 1.12 | 1.12 |
| Salt | 0.50 | 0.50 | 0.50 |
| Premix 2 | 0.50 | 0.50 | 0.50 |
| Nutrient composition | |||
| ME (MJ/kg of DM) | 8.81 | 11.72 | 11.72 |
| CP (% of DM) | 10.81 | 10.79 | 10.79 |
| NDF (% of DM) | 44.28 | 26.71 | 26.71 |
| ADF (% of DM) | 23.89 | 13.27 | 13.27 |
| Starch (% of DM) | 11.50 | 48.38 | 48.38 |
| Calcium (% of DM) | 0.81 | 0.78 | 0.78 |
| Phosphorus (% of DM) | 0.47 | 0.42 | 0.42 |
| Thiamine (mg/kg of DM) | 1.20 | 1.90 | 201.90 |
| Classification | Enzyme | Description | CON | HC | HCT | SEM | p-Value |
|---|---|---|---|---|---|---|---|
| Cellulose | CBM6 | binding to cellulose | 3.12 × 10−4 a | 1.43 × 10−4 b | 2.16 × 10−4 ab | 4.49 × 10−5 | 0.026 |
| CBM16 | binding to cellulose | 4.01 × 10−6 ab | 2.42 × 10−7 b | 6.26 × 10−6 a | 1.79 × 10−6 | 0.040 | |
| GH3 | β-glucosidase | 1.95 × 10−3 a | 9.30 × 10−4 b | 2.03 × 10−3 a | 3.13 × 10−4 | 0.022 | |
| GH51 | endoglucanase | 4.89 × 10−4 a | 1.88 × 10−4 b | 3.03 × 10−4 b | 8.84 × 10−5 | 0.038 | |
| GH9 | β-xylosidase | 2.86 × 10−4 b | 6.13 × 10−4 a | 2.56 × 10−4 b | 4.73 × 10−5 | <0.001 | |
| GH148 | β-1,3-glucanase | 1.92 × 10−5 a | 4.38 × 10−6 b | 6.67 × 10−6 b | 1.99 × 10−6 | 0.001 | |
| GH45 | endoglucanase | 8.11 × 10−6 | 1.76 × 10−5 | 7.69 × 10−6 | 8.20 × 10−6 | 0.442 | |
| GH8 | chitosanase | 8.00 × 10−5 | 4.78 × 10−5 | 5.66 × 10−5 | 3.22 × 10−5 | 0.257 | |
| Hemicelluloses | GH2 | β-galactosidase | 1.73 × 10−3 a | 6.68 × 10−4 b | 1.26 × 10−3 a | 1.65 × 10−4 | 0.002 |
| GT2 | β-galactosidase | 2.68 × 10−3 a | 1.25 × 10−3 b | 2.44 × 10−3 a | 3.73 × 10−4 | 0.018 | |
| GH35 | β-galactosidase | 2.10 × 10−4 a | 7.05 × 10−5 b | 1.72 × 10−4 ab | 3.77 × 10−5 | 0.024 | |
| GH5 | chitosanase | 6.09 × 10−4 a | 3.05 × 10−4 b | 3.80 × 10−4 b | 9.06 × 10−5 | 0.035 | |
| GH67 | α-glucuronidase | 1.01 × 10−4 a | 2.79 × 10−5 b | 6.92 × 10−5 ab | 1.65 × 10−5 | 0.013 | |
| CE1 | Acetyl xyla esterase | 7.45 × 10−4 a | 3.19 × 10−4 b | 5.37 × 10−4 ab | 1.23 × 10−4 | 0.037 | |
| Total | 2.39 × 10−2 a | 1.27 × 10−2 b | 2.03 × 10−2 a | 2.40 × 10−3 | 0.009 |
| Enzyme | Description | CON | HC | HCT | SEM | p-Value |
|---|---|---|---|---|---|---|
| CBM41 | starch-binding | 4.38 × 10−6 b | 2.35 × 10−6 bc | 8.54 × 10−6 a | 1.19 × 10−6 | 0.005 |
| GH97 | glucoamylase | 7.08 × 10−4 a | 1.75 × 10−4 b | 4.94 × 10−4 a | 1.02 × 10−4 | 0.006 |
| GH133 | amylo-α-1,6-glucosidase | 4.73 × 10−5 b | 1.60 × 10−4 a | 1.98 × 10−4 a | 1.92 × 10−5 | 0.001 |
| GH13 | α-amylase | 1.07 × 10−3 a | 5.32 × 10−4 b | 1.19 × 10−3 a | 1.59 × 10−4 | 0.018 |
| GH31 | α-glucosidase | 2.96 × 10−4 b | 5.43 × 10−4 a | 6.83 × 10−4 a | 7.57 × 10−5 | 0.006 |
| GH77 | amylomaltase | 3.55 × 10−4 | 3.92 × 10−4 | 4.33 × 10−4 | 3.79 × 10−5 | 0.417 |
| GH57 | α-amylase | 1.41 × 10−4 | 1.14 × 10−4 | 1.79 × 10−4 | 4.83 × 10−5 | 0.452 |
| GH4 | α-glucosidase | 2.07 × 10−5 | 1.35 × 10−5 | 2.40 × 10−5 | 5.39 × 10−6 | 0.461 |
| GH63 | α-1,3-glucosidase | 2.17 × 10−5 | 2.25 × 10−5 | 2.41 × 10−5 | 5.43 × 10−6 | 0.219 |
| Total | 5.45 × 10−3 b | 4.23 × 10−3 b | 6.776 × 10−3 a | 6.12 × 10−4 | 0.018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Wang, C.; Peng, A.; Zhang, H.; Wang, H. Metagenomic Insight: Dietary Thiamine Supplementation Promoted the Growth of Carbohydrate-Associated Microorganisms and Enzymes in the Rumen of Saanen Goats Fed High-Concentrate Diets. Microorganisms 2021, 9, 632. https://doi.org/10.3390/microorganisms9030632
Zhang Y, Wang C, Peng A, Zhang H, Wang H. Metagenomic Insight: Dietary Thiamine Supplementation Promoted the Growth of Carbohydrate-Associated Microorganisms and Enzymes in the Rumen of Saanen Goats Fed High-Concentrate Diets. Microorganisms. 2021; 9(3):632. https://doi.org/10.3390/microorganisms9030632
Chicago/Turabian StyleZhang, Ying, Chao Wang, Along Peng, Hao Zhang, and Hongrong Wang. 2021. "Metagenomic Insight: Dietary Thiamine Supplementation Promoted the Growth of Carbohydrate-Associated Microorganisms and Enzymes in the Rumen of Saanen Goats Fed High-Concentrate Diets" Microorganisms 9, no. 3: 632. https://doi.org/10.3390/microorganisms9030632
APA StyleZhang, Y., Wang, C., Peng, A., Zhang, H., & Wang, H. (2021). Metagenomic Insight: Dietary Thiamine Supplementation Promoted the Growth of Carbohydrate-Associated Microorganisms and Enzymes in the Rumen of Saanen Goats Fed High-Concentrate Diets. Microorganisms, 9(3), 632. https://doi.org/10.3390/microorganisms9030632