
The Two-Component System 09 Regulates Pneumococcal Carbohydrate Metabolism and Capsule Expression

 , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Molecular Biological Techniques
2.3. Generation of Pneumococcal Mutants
2.4. RNA Purification
2.5. RNA-Sequencing
2.6. Quantitative Real-Time PCR (qPCR)
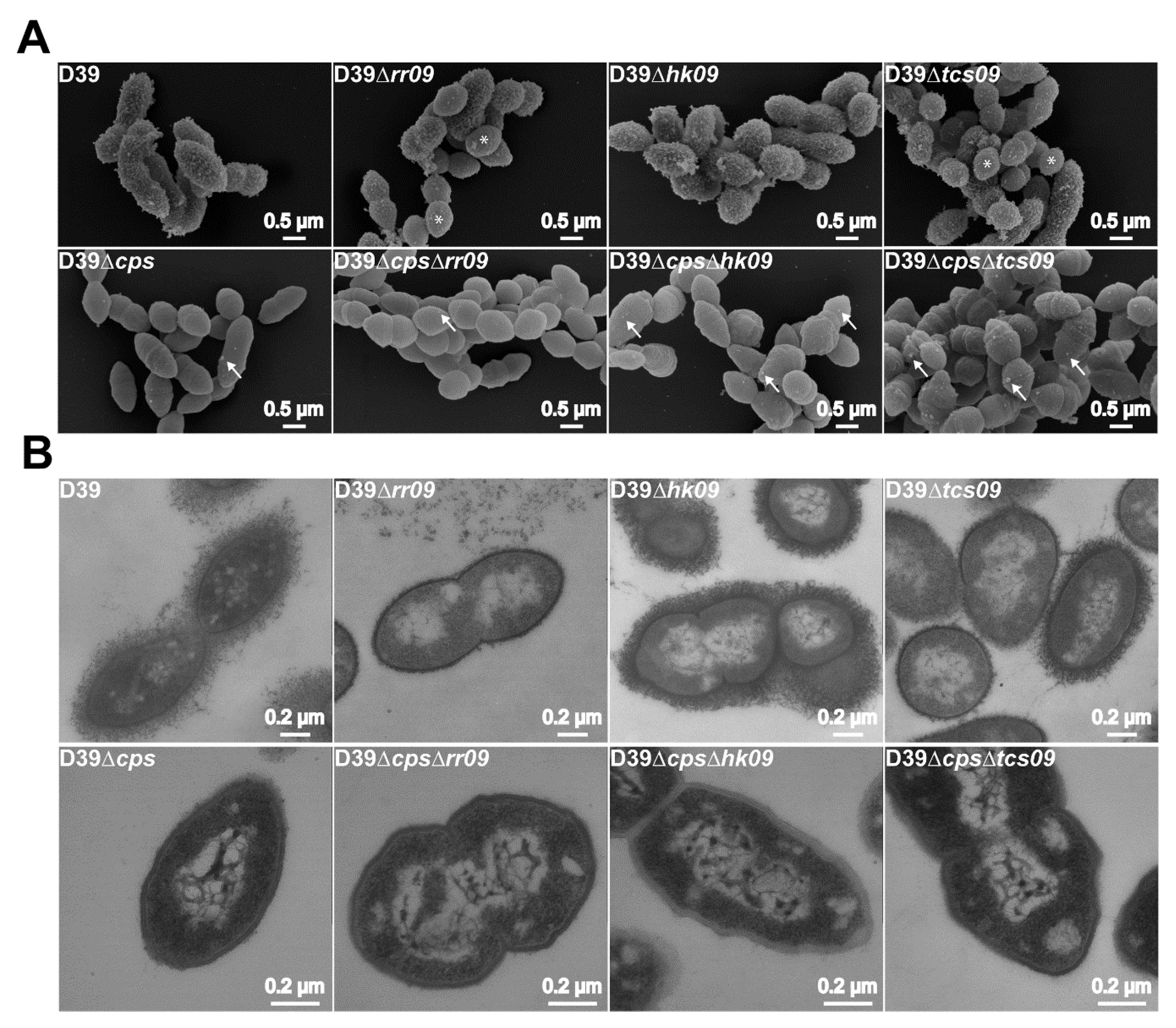
2.7. Field Emission Scanning Electron Microscopy (FESEM)
2.8. Transmission Electron Microscopy (TEM)
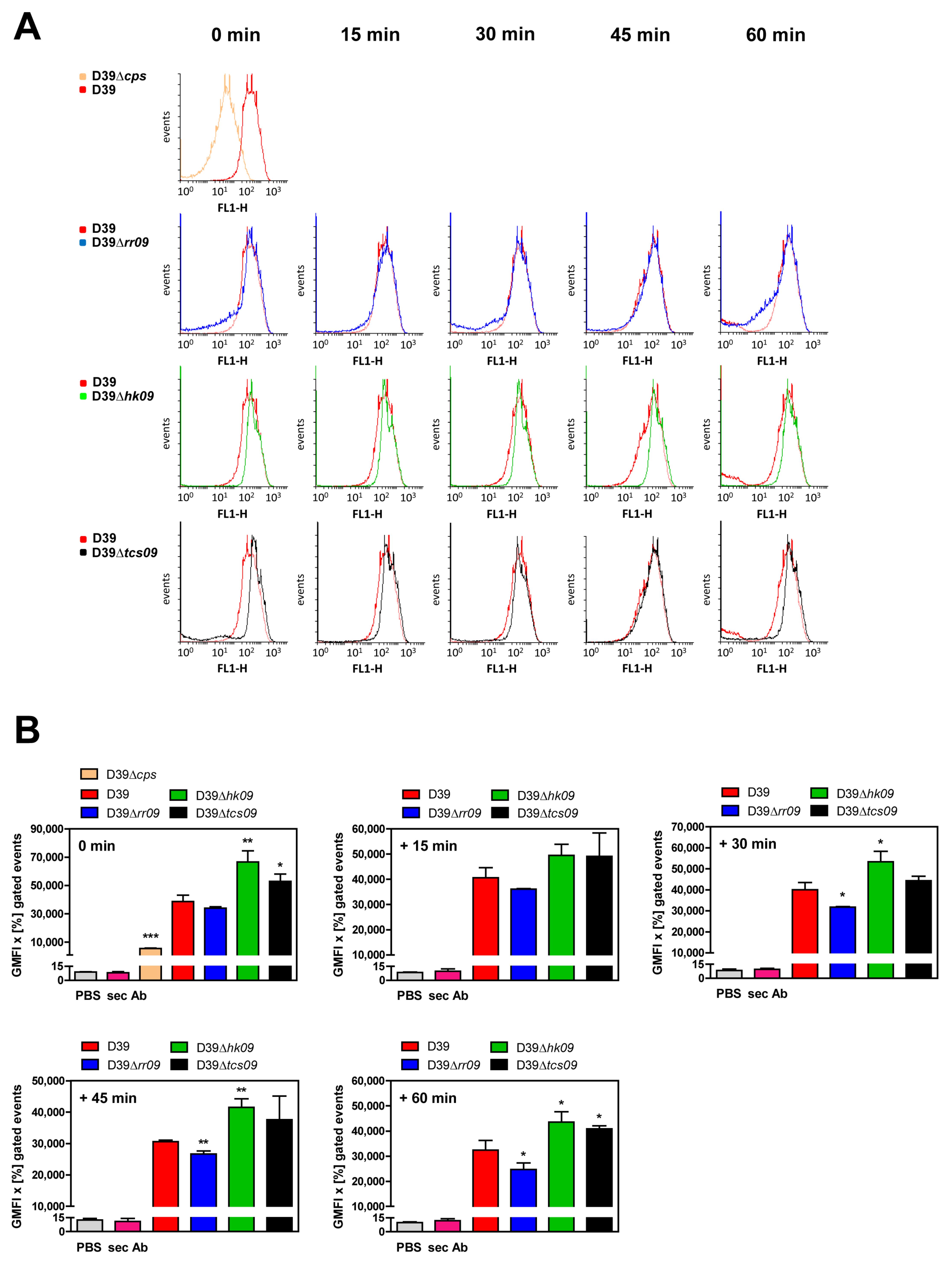
2.9. Flow Cytometric Analysis of Capsular Polysaccharide Abundance
2.10. Visualization of Pneumococcal Colony Phase Variation
2.11. Autolysis Assay with Triton X-100
2.12. Hydrogen Peroxide Toxicity Test
2.13. Statistical Analysis
3. Results
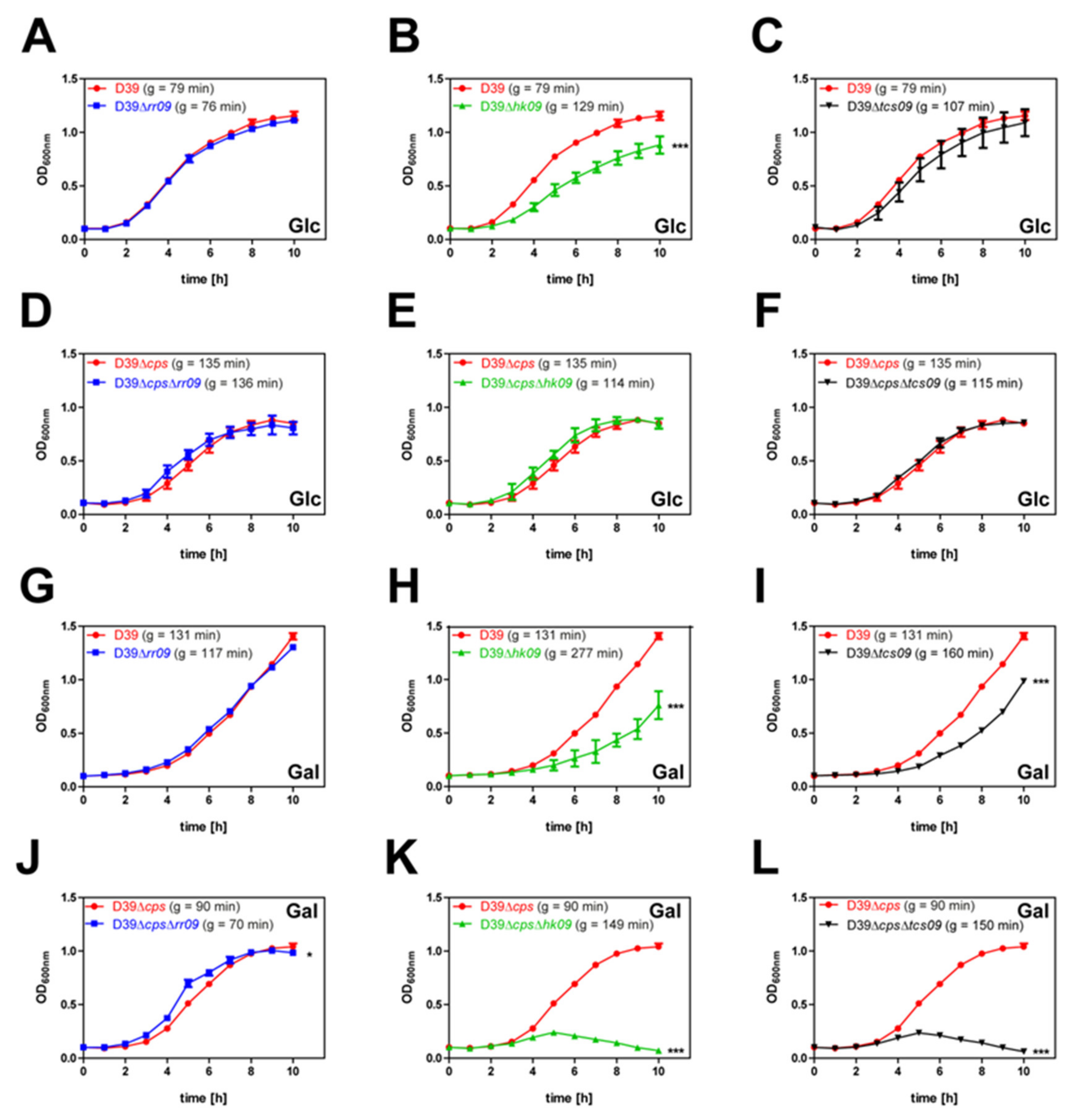
3.1. HK09 of TCS09 Is Essential for Optimal Growth Of Pneumococci under Defined In Vitro Conditions
3.2. Analysis of the D39 Transcriptome under In Vitro Conditions
3.3. HK09 Regulates Capsule Expression and Carbohydrate Metabolism
3.4. Phenotypic Characterization of TCS09-Deficient Pneumococci
3.5. Impact of TCS09 on the CPS Amount on the Pneumococcal Surface
3.6. Impact of TCS09 on Pneumococcal Phase Variation
3.7. Autolysis Assay with Triton X-100
3.8. Impact of TCS09 on Resistance against Oxidative Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- West, A.H.; Stock, A.M. Histidine kinases and response regulator proteins in two-component signaling systems. Trends Biochem. Sci. 2001, 26, 369–376. [Google Scholar] [CrossRef]
- Rowland, M.A.; Deeds, E.J. Crosstalk and the evolution of specificity in two-component signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 5550. [Google Scholar] [CrossRef]
- Lange, R.; Wagner, C.; de Saizieu, A.; Flint, N.; Molnos, J.; Stieger, M.; Caspers, P.; Kamber, M.; Keck, W.; Amrein, K.E. Domain organization and molecular characterization of 13 two-component systems identified by genome sequencing of Streptococcus pneumoniae. Gene 1999, 237, 223–234. [Google Scholar] [CrossRef]
- Throup, J.P.; Koretke, K.K.; Bryant, A.P.; Ingraham, K.A.; Chalker, A.F.; Ge, Y.; Marra, A.; Wallis, N.G.; Brown, J.R.; Holmes, D.J.; et al. A genomic analysis of two-component signal transduction in Streptococcus pneumoniae. Mol. Microbiol. 2000, 35, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Mejia, A.; Gámez, G.; Hammerschmidt, S. Streptococcus pneumoniae two-component regulatory systems: The interplay of the pneumococcus with its environment. Int. J. Med. Microbiol. 2018, 308, 722–737. [Google Scholar] [CrossRef] [PubMed]
- Reinoso-Vizcaíno, N.M.; Cian, M.B.; Cortes, P.R.; Olivero, N.B.; Hernandez-Morfa, M.; Piñas, G.E.; Badapanda, C.; Rathore, A.; Perez, D.R.; Echenique, J. The pneumococcal two-component system SirRH is linked to enhanced intracellular survival of Streptococcus pneumoniae in influenza-infected pulmonary cells. PLoS Pathog. 2020, 16, e1008761. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Mejia, A.; Gámez, G.; Hirschmann, S.; Kluger, V.; Rath, H.; Böhm, S.; Voss, F.; Kakar, N.; Petruschka, L.; Völker, U.; et al. Pneumococcal Metabolic Adaptation and Colonization Are Regulated by the Two-Component Regulatory System 08. mSphere 2018, 3, e00165-18. [Google Scholar] [CrossRef]
- Grebe, T.W.; Stock, J.B. The Histidine Protein Kinase Superfamily. In Advances in Microbial Physiology; Poole, R.K., Ed.; Academic Press: Cambridge, MA, USA, 1999; Volume 41, pp. 139–227. [Google Scholar]
- Gao, R.; Mack, T.R.; Stock, A.M. Bacterial response regulators: Versatile regulatory strategies from common domains. Trends Biochem. Sci. 2007, 32, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Stock, A.M. Biological Insights from Structures of Two-Component Proteins. Annu. Rev. Microbiol. 2009, 63, 133–154. [Google Scholar] [CrossRef] [PubMed]
- Nikolskaya, A.N.; Galperin, M.Y. A novel type of conserved DNA-binding domain in the transcriptional regulators of the AlgR/AgrA/LytR family. Nucleic Acids Res. 2002, 30, 2453–2459. [Google Scholar] [CrossRef]
- Hava, D.L.; Camilli, A. Large-scale identification of serotype 4 Streptococcus pneumoniae virulence factors. Mol. Microbiol. 2002, 45, 1389–1406. [Google Scholar] [CrossRef] [PubMed]
- Novak, R.; Charpentier, E.; Braun, J.S.; Park, E.; Murti, S.; Tuomanen, E.; Masure, R. Extracellular targeting of choline-binding proteins in Streptococcus pneumoniae by a zinc metalloprotease. Mol. Microbiol. 2000, 36, 366–376. [Google Scholar] [CrossRef]
- Blue, C.E.; Paterson, G.K.; Kerr, A.R.; Bergé, M.; Claverys, J.P.; Mitchell, T.J. ZmpB, a Novel Virulence Factor of Streptococcus pneumoniae That Induces Tumor Necrosis Factor Alpha Production in the Respiratory Tract. Infect. Immun. 2003, 71, 4925. [Google Scholar] [CrossRef] [PubMed]
- Hendriksen, W.T.; Silva, N.; Bootsma, H.J.; Blue, C.E.; Paterson, G.K.; Kerr, A.R.; de Jong, A.; Kuipers, O.P.; Hermans, P.W.M.; Mitchell, T.J. Regulation of Gene Expression in Streptococcus pneumoniae by Response Regulator 09 Is Strain Dependent. J. Bacteriol. 2007, 189, 1382. [Google Scholar] [CrossRef] [PubMed]
- Blue, C.E.; Mitchell, T.J. Contribution of a Response Regulator to the Virulence of Streptococcus pneumoniae Is Strain Dependent. Infect. Immun. 2003, 71, 4405. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, J.; Li, J.-W.; Li, J.; Huang, Y.; Wang, S.; Zhang, J.-R. Regulation of pneumococcal epigenetic and colony phases by multiple two-component regulatory systems. PLoS Pathog. 2020, 16, e1008417. [Google Scholar] [CrossRef]
- Li, J.; Wang, J.; Ruiz-Cruz, S.; Espinosa, M.; Zhang, J.-R.; Bravo, A. In vitro DNA Inversions Mediated by the PsrA Site-Specific Tyrosine Recombinase of Streptococcus pneumoniae. Front. Mol. Biosci. 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Gierok, P.; Petruschka, L.; Lalk, M.; Mäder, U.; Hammerschmidt, S. Regulation of the Arginine Deiminase System by ArgR2 Interferes with Arginine Metabolism and Fitness of Streptococcus pneumoniae. mBio 2014, 5, e01858-14. [Google Scholar] [CrossRef]
- Pracht, D.; Elm, C.; Gerber, J.; Bergmann, S.; Rohde, M.; Seiler, M.; Kim, K.S.; Jenkinson, H.F.; Nau, R.; Hammerschmidt, S. PavA of Streptococcus pneumoniae Modulates Adherence, Invasion, and Meningeal Inflammation. Infect. Immun. 2005, 73, 2680. [Google Scholar] [CrossRef] [PubMed]
- Rennemeier, C.; Hammerschmidt, S.; Niemann, S.; Inamura, S.; Zähringer, U.; Kehrel, B.E. Thrombospondin-1 promotes cellular adherence of Gram-positive pathogens via recognition of peptidoglycan. FASEB J. 2007, 21, 3118–3132. [Google Scholar] [CrossRef]
- Chen, S.; Songkumarn, P.; Liu, J.; Wang, G.-L. A Versatile Zero Background T-Vector System for Gene Cloning and Functional Genomics. Plant Physiol. 2009, 150, 1111. [Google Scholar] [CrossRef]
- Shishkin, A.A.; Giannoukos, G.; Kucukural, A.; Ciulla, D.; Busby, M.; Surka, C.; Chen, J.; Bhattacharyya, R.P.; Rudy, R.F.; Patel, M.M.; et al. Simultaneous generation of many RNA-seq libraries in a single reaction. Nat. Methods 2015, 12, 323–325. [Google Scholar] [CrossRef]
- Bhattacharyya, R.P.; Bandyopadhyay, N.; Ma, P.; Son, S.S.; Liu, J.; He, L.L.; Wu, L.; Khafizov, R.; Boykin, R.; Cerqueira, G.C.; et al. Simultaneous detection of genotype and phenotype enables rapid and accurate antibiotic susceptibility determination. Nat. Med. 2019, 25, 1858–1864. [Google Scholar] [CrossRef] [PubMed]
- Metsalu, T.; Vilo, J. ClustVis: A web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Schmidt, F.; Kakar, N.; Meyer, T.C.; Depke, M.; Masouris, I.; Burchhardt, G.; Gómez-Mejia, A.; Dhople, V.; Håvarstein, L.S.; Sun, Z.; et al. In vivo proteomics identifies the competence regulon and AliB oligopeptide transporter as pathogenic factors in pneumococcal meningitis. PLoS Pathog. 2019, 15, e1007987. [Google Scholar] [CrossRef]
- Heß, N.; Waldow, F.; Kohler, T.P.; Rohde, M.; Kreikemeyer, B.; Gómez-Mejia, A.; Hain, T.; Schwudke, D.; Vollmer, W.; Hammerschmidt, S.; et al. Lipoteichoic acid deficiency permits normal growth but impairs virulence of Streptococcus pneumoniae. Nat. Commun. 2017, 8, 2093. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.; Shafeeq, S.; Ahmed, H.; Kuipers, O.P. N-acetylgalatosamine-Mediated Regulation of the aga Operon by AgaR in Streptococcus pneumoniae. Front. Cell. Infect. Microbiol. 2016, 6, 101. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Xu, H.; Zheng, Y.; Wang, L.; Zhang, X.; Yin, Y. CpsR, a GntR family regulator, transcriptionally regulates capsular polysaccharide biosynthesis and governs bacterial virulence in Streptococcus pneumoniae. Sci. Rep. 2016, 6, 29255. [Google Scholar] [CrossRef] [PubMed]
- Bienert, G.P.; Schjoerring, J.K.; Jahn, T.P. Membrane transport of hydrogen peroxide. Biochim. Biophys. Acta BBA Biomembr. 2006, 1758, 994–1003. [Google Scholar] [CrossRef] [PubMed]
- Linley, E.; Denyer, S.P.; McDonnell, G.; Simons, C.; Maillard, J.-Y. Use of hydrogen peroxide as a biocide: New consideration of its mechanisms of biocidal action. J. Antimicrob. Chemother. 2012, 67, 1589–1596. [Google Scholar] [CrossRef] [PubMed]
- Seaver, L.C.; Imlay, J.A. Hydrogen Peroxide Fluxes and Compartmentalization inside Growing Escherichia coli. J. Bacteriol. 2001, 183, 7182. [Google Scholar] [CrossRef]
- Sousa-Lopes, A.; Antunes, F.; Cyrne, L.; Marinho, H.S. Decreased cellular permeability to H2O2 protects Saccharomyces cerevisiae cells in stationary phase against oxidative stress. FEBS Lett. 2004, 578, 152–156. [Google Scholar] [CrossRef]
- Doran, K.S.; Fulde, M.; Gratz, N.; Kim, B.J.; Nau, R.; Prasadarao, N.; Schubert-Unkmeir, A.; Tuomanen, E.I.; Valentin-Weigand, P. Host-pathogen interactions in bacterial meningitis. Acta Neuropathol. 2016, 131, 185–209. [Google Scholar] [CrossRef]
- Subramanian, K.; Henriques-Normark, B.; Normark, S. Emerging concepts in the pathogenesis of the Streptococcus pneumoniae: From nasopharyngeal colonizer to intracellular pathogen. Cell Microbiol. 2019, 21, e13077. [Google Scholar] [CrossRef]
- Weiser, J.N.; Ferreira, D.M.; Paton, J.C. Streptococcus pneumoniae: Transmission, colonization and invasion. Nat. Rev. Microbiol. 2018, 16, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Ogunniyi, A.D.; Giammarinaro, P.; Paton, J.C. The genes encoding virulence-associated proteins and the capsule of Streptococcus pneumoniae are upregulated and differentially expressed in vivo. Microbiology 2002, 148, 2045–2053. [Google Scholar] [CrossRef]
- Orihuela, C.J.; Radin, J.N.; Sublett, J.E.; Gao, G.; Kaushal, D.; Tuomanen, E.I. Microarray analysis of pneumococcal gene expression during invasive disease. Infect.Immun. 2004, 72, 5582–5596. [Google Scholar] [CrossRef] [PubMed]
- Schulz, C.; Hammerschmidt, S. Exploitation of physiology and metabolomics to identify pneumococcal vaccine candidates. Expert Rev. Vaccines 2013, 12, 1061–1075. [Google Scholar] [CrossRef]
- Carvalho, S.M.; Kloosterman, T.G.; Kuipers, O.P.; Neves, A.R. CcpA Ensures Optimal Metabolic Fitness of Streptococcus pneumoniae. PLoS ONE 2011, 6, e26707. [Google Scholar] [CrossRef]
- Hartel, T.; Eylert, E.; Schulz, C.; Petruschka, L.; Gierok, P.; Grubmuller, S.; Lalk, M.; Eisenreich, W.; Hammerschmidt, S. Characterization of central carbon metabolism of Streptococcus pneumoniae by isotopologue profiling. J. Biol. Chem. 2012, 287, 4260–4274. [Google Scholar] [CrossRef] [PubMed]
- Leonard, A.; Gierok, P.; Methling, K.; Gomez-Mejia, A.; Hammerschmidt, S.; Lalk, M. Metabolic inventory of Streptococcus pneumoniae growing in a chemical defined environment. Int. J. Med. Microbiol. 2018, 308, 705–712. [Google Scholar] [CrossRef] [PubMed]
- Leonard, A.; Lalk, M. Infection and metabolism—Streptococcus pneumoniae metabolism facing the host environment. Cytokine 2018, 112, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Tettelin, H.; Nelson, K.E.; Paulsen, I.T.; Eisen, J.A.; Read, T.D.; Peterson, S.; Heidelberg, J.; DeBoy, R.T.; Haft, D.H.; Dodson, R.J. Complete Genome Sequence of a Virulent Isolate of Streptococcus pneumoniae. Science 2001, 293, 498. [Google Scholar] [CrossRef]
- Afzal, M.; Shafeeq, S.; Kuipers, O.P. LacR is a repressor of lacABCD and LacT is an activator of lacTFEG, constituting the lac gene cluster in Streptococcus pneumoniae. Appl. Env. Microbiol. 2014, 80, 5349–5358. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, P.; Al-Bayati, F.A.; Andrew, P.W.; Neves, A.R.; Yesilkaya, H. Lactate dehydrogenase is the key enzyme for pneumococcal pyruvate metabolism and pneumococcal survival in blood. Infect. Immun. 2014, 82, 5099–5109. [Google Scholar] [CrossRef] [PubMed]
- Bidossi, A.; Mulas, L.; Decorosi, F.; Colomba, L.; Ricci, S.; Pozzi, G.; Deutscher, J.; Viti, C.; Oggioni, M.R. A Functional Genomics Approach to Establish the Complement of Carbohydrate Transporters in Streptococcus pneumoniae. PLoS ONE 2012, 7, e33320. [Google Scholar] [CrossRef]
- Slager, J.; Aprianto, R.; Veening, J.-W. Deep genome annotation of the opportunistic human pathogen Streptococcus pneumoniae D39. Nucleic Acids Res. 2018, 46, 9971–9989. [Google Scholar] [CrossRef]
- Andersson, B.; Dahmén, J.; Frejd, T.; Leffler, H.; Magnusson, G.; Noori, G.; Edén, C.S. Identification of an active disaccharide unit of a glycoconjugate receptor for pneumococci attaching to human pharyngeal epithelial cells. J. Exp. Med. 1983, 158, 559–570. [Google Scholar] [CrossRef]
- Jeong, J.K.; Kwon, O.; Lee, Y.M.; Oh, D.-B.; Lee, J.M.; Kim, S.; Kim, E.-H.; Le, T.N.; Rhee, D.-K.; Kang, H.A. Characterization of the Streptococcus pneumoniae BgaC Protein as a Novel Surface β-Galactosidase with Specific Hydrolysis Activity for the Galβ1-3GlcNAc Moiety of Oligosaccharides. J. Bacteriol. 2009, 191, 3011–3023. [Google Scholar] [CrossRef][Green Version]
- Aprianto, R.; Slager, J.; Holsappel, S.; Veening, J.-W. High-resolution analysis of the pneumococcal transcriptome under a wide range of infection-relevant conditions. Nucleic Acids Res. 2018, 46, 9990–10006. [Google Scholar] [CrossRef] [PubMed]
- Shaper, M.; Hollingshead, S.K.; Benjamin, W.H., Jr.; Briles, D.E. PspA protects Streptococcus pneumoniae from killing by apolactoferrin, and antibody to PspA enhances killing of pneumococci by apolactoferrin [corrected]. Infect. Immun. 2004, 72, 5031–5040. [Google Scholar] [CrossRef]
- Tu, A.-H.T.; Fulgham, R.L.; McCrory, M.A.; Briles, D.E.; Szalai, A.J. Pneumococcal Surface Protein A Inhibits Complement Activation by Streptococcus pneumoniae. Infect. Immun. 1999, 67, 4720. [Google Scholar] [CrossRef] [PubMed]
- Hammerschmidt, S.; Bethe, G.; Remane, P.H.; Chhatwal, G.S. Identification of Pneumococcal Surface Protein A as a Lactoferrin-Binding Protein of Streptococcus pneumoniae. Infect. Immun. 1999, 67, 1683. [Google Scholar] [CrossRef] [PubMed]
- Kallio, A.; Sepponen, K.; Hermand, P.; Denoël, P.; Godfroid, F.; Melin, M. Role of Pht Proteins in Attachment of Streptococcus pneumoniae to Respiratory Epithelial Cells. Infect. Immun. 2014, 82, 1683. [Google Scholar] [CrossRef] [PubMed]
- Plumptre, C.D.; Hughes, C.E.; Harvey, R.M.; Eijkelkamp, B.A.; McDevitt, C.A.; Paton, J.C. Overlapping Functionality of the Pht Proteins in Zinc Homeostasis of Streptococcus pneumoniae. Infect. Immun. 2014, 82, 4315. [Google Scholar] [CrossRef]
- Hathaway, L.J.; Brugger, S.D.; Morand, B.; Bangert, M.; Rotzetter, J.U.; Hauser, C.; Graber, W.A.; Gore, S.; Kadioglu, A.; Mühlemann, K. Capsule Type of Streptococcus pneumoniae Determines Growth Phenotype. PLoS Pathog. 2012, 8, e1002574. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.O.; Weiser, J.N. Association of Intrastrain Phase Variation in Quantity of Capsular Polysaccharide and Teichoic Acid with the Virulence of Streptococcus pneumoniae. J. Infect. Dis. 1998, 177, 368–377. [Google Scholar] [CrossRef]
- Dorward, D.W.; Garon, C.F.; Judd, R.C. Export and intercellular transfer of DNA via membrane blebs of Neisseria gonorrhoeae. J. Bacteriol. 1989, 171, 2499. [Google Scholar] [CrossRef] [PubMed]
- Hoekstra, D.; van der Laan, J.W.; de Leij, L.; Witholt, B. Release of outer membrane fragments from normally growing Escherichia coli. Biochim. Biophys. Acta BBA Biomembr. 1976, 455, 889–899. [Google Scholar] [CrossRef]
- Olaya-Abril, A.; Prados-Rosales, R.; McConnell, M.J.; Martín-Peña, R.; González-Reyes, J.A.; Jiménez-Munguía, I.; Gómez-Gascón, L.; Fernández, J.; Luque-García, J.L.; García-Lidón, C.; et al. Characterization of protective extracellular membrane-derived vesicles produced by Streptococcus pneumoniae. J. Proteom. 2014, 106, 46–60. [Google Scholar] [CrossRef] [PubMed]
- Elhenawy, W.; Debelyy, M.O.; Feldman, M.F. Preferential Packing of Acidic Glycosidases and Proteases into Bacteroides Outer Membrane Vesicles. mBio 2014, 5, e00909-14. [Google Scholar] [CrossRef]
- Horstman, A.L.; Kuehn, M.J. Bacterial Surface Association of Heat-labile Enterotoxin through Lipopolysaccharide after Secretion via the General Secretory Pathway. J. Biol. Chem. 2002, 277, 32538–32545. [Google Scholar] [CrossRef] [PubMed]
- Manning, A.J.; Kuehn, M.J. Contribution of bacterial outer membrane vesicles to innate bacterial defense. BMC Microbiol. 2011, 11, 258. [Google Scholar] [CrossRef]
- Mashburn, L.M.; Whiteley, M. Membrane vesicles traffic signals and facilitate group activities in a prokaryote. Nature 2005, 437, 422–425. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, M.; Cárcamo-Oyarce, G.; Yamamoto, T.; Eisenstein, F.; Hsiao, C.-C.; Kurosawa, M.; Gademann, K.; Pilhofer, M.; Nomura, N.; Eberl, L. Prophage-triggered membrane vesicle formation through peptidoglycan damage in Bacillus subtilis. Nat. Commun. 2017, 8, 481. [Google Scholar] [CrossRef] [PubMed]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Cárcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Thompson, C.D.; Weidenmaier, C.; Lee, J.C. Release of Staphylococcus aureus extracellular vesicles and their application as a vaccine platform. Nat. Commun. 2018, 9, 1379. [Google Scholar] [CrossRef]
- Fernebro, J.; Andersson, I.; Sublett, J.; Morfeldt, E.; Novak, R.; Tuomanen, E.; Normark, S.; Normark, B.H. Capsular Expression in Streptococcus pneumoniae Negatively Affects Spontaneous and Antibiotic-Induced Lysis and Contributes to Antibiotic Tolerance. J. Infect. Dis. 2004, 189, 328–338. [Google Scholar] [CrossRef] [PubMed][Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Capsule Type | Resistance | Knockout Genes | Reference |
|---|---|---|---|---|
| D39 | 2 | - | - | [20] |
| D39Δrr09 | 2 | erythromycin | spd_0574 | This study |
| D39Δhk09 | 2 | erythromycin | spd_0575 | This study |
| D39Δtcs09 | 2 | erythromycin | spd_0574, spd_0575 | This study |
| D39Δcps | 2 | kanamycin | spd_0312–spd_0333 | [21] |
| D39ΔcpsΔrr09 | 2 | kanamycin, erythromycin | spd_0312–spd_0333, spd_0574 | This study |
| D39ΔcpsΔhk09 | 2 | kanamycin, erythromycin | spd_0312–spd_0333, spd_0575 | This study |
| D39ΔcpsΔtcs09 | 2 | kanamycin, erythromycin | spd_0312–spd_0333 spd_0574, spd_0575 | This study |
| No. | Primer | Sequence 5′-3′ | Restriction Site |
|---|---|---|---|
| 100 | MC-Erm-R | CCCGGGGAAATTTTGATATCGATAAGCTTGAATTCCCGTAGGCGCTAGGGACCTC | HindIII |
| 105 | Erm Forw | GATGATGATGATCCCGGGTACCAAGCTTGAATTCACGGTTCGTGTTCGTGCTG | HindIII |
| 106 | Erm rev | AGTGAGTGAGTCCCGGGCTCGAGAAGCTTGAATTCGTAGGCGCTAGGGACCTC | HindIII |
| 1122 | rr09_Start Forw | CAGAACGAGCTTTCTCAAACC | |
| 1123 | rr09_End Rev | GCCTCCATTTTGCTAGACGA | |
| 1124 | rr09_End Forw | CTCACTGAAGCTTGAGATCGCAGAGAAGGTTGG | HindIII |
| 1125 | rr09_Start Rev | ATCATCGCTAGCGTAGGCTGCTACATTGACC | NheI |
| 1126 | hk09_Start Forw | GGTCAATGTAGCAGCCTACGA | |
| 1127 | hk09_End Rev | CGCACCTCCGATTAATTTTG | |
| 1128 | hk09_End Forw | CTCACTGAAGCTTCGATCAACGGCTCAAACTTC | HindIII |
| 1129 | hk09_Start Rev | ATCATCGCTAGCCAACCAGAGCTAGGAGAA | NheI |
| 1232 | rr09_sr_fw HindIII | ATCATCAAGCTTGTAGGCTGCTACATTGACC | HindIII |
| 1234 | Nhe_erm f | GCTTGCTAGCGACGGTTCGTGTTCGTGCTG | NheI |
| Plasmids | Properties | Source |
|---|---|---|
| pGXT | pGEM-T Easy backbone, linker region with toxic gene ccdB (Flanked by two XcmI sites), TA cloning vector (3047 bp), AmpR, Pt7, lacZ | [22] |
| pGSP72N | pSP72 derivative, HindIII restriction site eliminated, generation of a NheI restriction site | This study |
| pSP72 | Cloning vector (2462 bp), AmpR | Promega |
| pGXTΔrr09::ermR | pGXT derivative vector with the subcloned 5′ and 3′-end homolog fragments of spd_0574 interrupted by ermB resistance gene cassette for mutagenesis | This study |
| pGSP72NΔhk09::ermR | pGSP72N derivative vector with the subcloned 5′ and 3′-end homolog fragments of spd_0575 interrupted by ermB resistance gene cassette for mutagenesis | This study |
| pSP72Δtcs09::ermR | pSP72 derivative vector with the subcloned 5′ end homolog fragment of spd_0574 and 3′ end homolog fragment of spd_0575 interrupted by ermB resistance gene cassette for mutagenesis | This study |
| Target Gene | Primer | Sequence 5′➔3′ |
|---|---|---|
| enolase (spd_1012) | enoRT_F enoRT_R | CGGACGTGGTATGGTTCCA TAGCCAATGATAGCTTCAGCA |
| agaR (spd_0064) | qPCR agaR Forward qPCR agaR Reverse | TCATTTCAATGTACGATGTCAG GTTTGCGTGCACGTGAAACG |
| gadV (spd_0066) | qPCR gadV Forward qPCR gadV Reverse | ACGACGAAGTTGTCAACAACG GAGACGTTGGCTATCGTATTT |
| celA (spd_0277) | qPCR celA Forward qPCR celA Reverse | ATGTTATGACTGCTGGTCGTC CCATTTCAGCAAAAAGTGCTAT |
| eng (spd_0335) | qPCR eng Forward qPCR eng Reverse | GTCCGGTTCTTGCAGATAGC CCACCTTAGGCGCTTCAAAA |
| spd_1588 | qPCR spd_1588 Forward qPCR spd_1588 Reverse | GTATTCATCTACTTAGCTGGAG ACATCACAACTAAAATGGATAATA |
| Strain | Growth Rate [h−1] | Max OD600nm | Generation Time ± SD [min] |
|---|---|---|---|
| glucose | |||
| D39 | 0.5252 | 1.199 | 79.3 ± 4.3 |
| D39∆rr09 | 0.5503 | 1.125 | 75.7 ± 2.6 |
| D39∆hk09 | 0.3226 | 0.943 | 128.7 ± 8.4 |
| D39∆tcs09 | 0.3859 | 1.173 | 107.3 ± 11.2 |
| D39∆cps | 0.3224 | 0.910 | 135.3 ± 36.8 |
| D39∆cps∆rr09 | 0.3296 | 0.927 | 135.8 ± 46.3 |
| D39∆cps∆hk09 | 0.3804 | 0.900 | 114.3 ± 35.6 |
| D39∆cps∆tcs09 | 0.3664 | 0.880 | 114.5 ± 14.4 |
| galactose | |||
| D39 | 0.3174 | 1.436 | 131.2 ± 4.5 |
| D39∆rr09 | 0.3546 | 1.313 | 117.4 ± 3.4 |
| D39∆hk09 | 0.1878 | 0.909 | 277.4 ± 148.3 |
| D39∆tcs09 | 0.2611 | 1.002 | 160.3 ± 14.4 |
| D39∆cps | 0.4614 | 1.073 | 90.1 ± 5.1 |
| D39∆cps∆rr09 | 0.5917 | 1.027 | 70.2 ± 1.9 |
| D39∆cps∆hk09 | 0.2830 | 0.251 | 148.6 ± 17.9 |
| D39∆cps∆tcs09 | 0.2779 | 0.241 | 149.7 ± 5.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirschmann, S.; Gómez-Mejia, A.; Mäder, U.; Karsunke, J.; Driesch, D.; Rohde, M.; Häussler, S.; Burchhardt, G.; Hammerschmidt, S. The Two-Component System 09 Regulates Pneumococcal Carbohydrate Metabolism and Capsule Expression. Microorganisms 2021, 9, 468. https://doi.org/10.3390/microorganisms9030468
Hirschmann S, Gómez-Mejia A, Mäder U, Karsunke J, Driesch D, Rohde M, Häussler S, Burchhardt G, Hammerschmidt S. The Two-Component System 09 Regulates Pneumococcal Carbohydrate Metabolism and Capsule Expression. Microorganisms. 2021; 9(3):468. https://doi.org/10.3390/microorganisms9030468
Chicago/Turabian StyleHirschmann, Stephanie, Alejandro Gómez-Mejia, Ulrike Mäder, Julia Karsunke, Dominik Driesch, Manfred Rohde, Susanne Häussler, Gerhard Burchhardt, and Sven Hammerschmidt. 2021. "The Two-Component System 09 Regulates Pneumococcal Carbohydrate Metabolism and Capsule Expression" Microorganisms 9, no. 3: 468. https://doi.org/10.3390/microorganisms9030468
APA StyleHirschmann, S., Gómez-Mejia, A., Mäder, U., Karsunke, J., Driesch, D., Rohde, M., Häussler, S., Burchhardt, G., & Hammerschmidt, S. (2021). The Two-Component System 09 Regulates Pneumococcal Carbohydrate Metabolism and Capsule Expression. Microorganisms, 9(3), 468. https://doi.org/10.3390/microorganisms9030468