
Diversity and Distribution of Resistance Markers in Pseudomonas aeruginosa International High-Risk Clones


Abstract
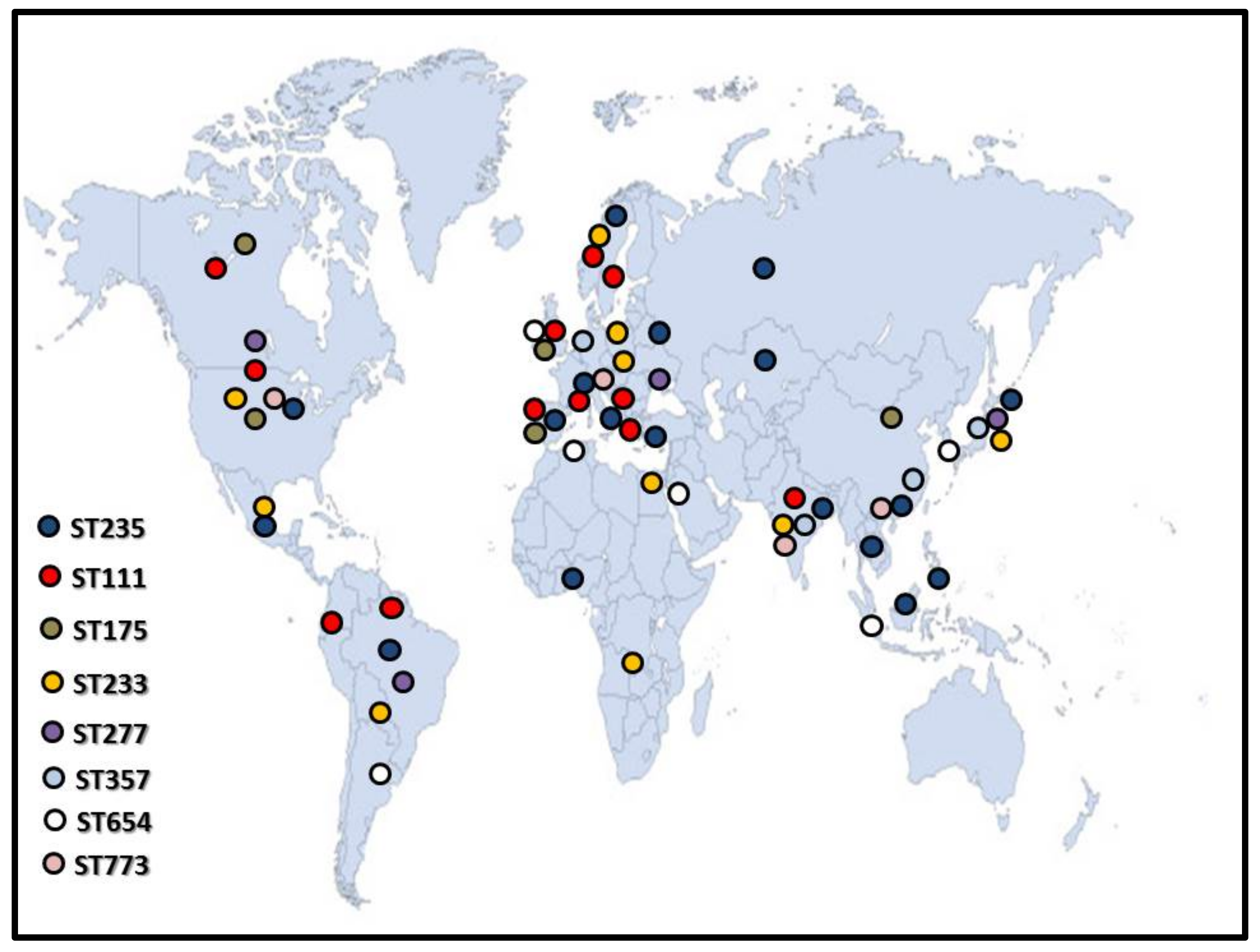
1. Introduction
2. ST235
3. ST111
4. ST175
5. ST233
6. ST277
7. ST357, ST654 and ST773
8. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bassetti, M.; Vena, A.; Croxatto, A.; Righi, E.; Guery, B. How to manage Pseudomonas aeruginosa infections. Drugs Context 2018, 7, 212527. [Google Scholar] [CrossRef] [PubMed]
- Burrows, L.L. The therapeutic pipeline for Pseudomonas aeruginosa infections. ACS Infect. Dis. 2018, 4, 1041–1047. [Google Scholar] [CrossRef]
- Römling, U.; Balsalobre, C. Biofilm infections, their resilience to therapy and innovative treatment strategies. J. Intern Med. 2012, 272, 541–561. [Google Scholar] [CrossRef]
- Rodriguez-Rojas, A.; Oliver, A.; Blazquez, J. Intrinsic and environmental mutagenesis drive diversification and persistence of Pseudomonas aeruginosa in chronic lung infections. J. Infect. Dis. 2012, 205, 121–127. [Google Scholar] [CrossRef]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Livermore, D.M. Has the era of untreatable infections arrived? J. Antimicrob. Chemother. 2009, 64 (Suppl. 1), i29–i36. [Google Scholar] [CrossRef] [PubMed]
- Botelho, J.; Grosso, F.; Peixe, L. Antibiotic resistance in Pseudomonas aeruginosa—Mechanisms, epidemiology and evolution. Drug Resist Updat. 2019, 44, 26–47. [Google Scholar] [CrossRef]
- Oliver, A.; Mulet, X.; López-Causapé, C.; Juan, C. The increasing threat of Pseudomonas aeruginosa high-risk clones. Drug Resist Updat. 2015, 21–22, 41–59. [Google Scholar] [CrossRef] [PubMed]
- Juhász, E.; Iván, M.; Pongrácz, J.; Kristóf, K. Uncommon non-fermenting Gram-negative rods as pathogens of lower respiratory tract infection. Orv. Hetil. 2018, 159, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Carattoli, A.; Fortini, D.; Galetti, R.; Garcia-Fernandez, A.; Nardi, G.; Orazi, D.; Capone, A.; Majolino, I.; Proia, A.; Mariani, B.; et al. Isolation of NDM-1-producing Pseudomonas aeruginosa sequence type ST235 from a stem cell transplant patient in Italy, May 2013. Euro. Surveill. 2013, 18, 20633. [Google Scholar] [CrossRef] [PubMed]
- Kos, V.N.; Déraspe, M.; McLaughlin, R.E.; Whiteaker, J.D.; Roy, P.H.; Alm, R.A.; Corbeil, J.; Gardner, H. The resistome of Pseudomonas aeruginosa in relationship to phenotypic susceptibility. Antimicrob. Agents Chemother. 2015, 59, 427–436. [Google Scholar] [CrossRef]
- Breidenstein, E.B.M.; de la Fuente-Núñez, C.; Hancock, R.E.W. Pseudomonas aeruginosa: All roads lead to resistance. Trends Microbiol. 2011, 19, 419–426. [Google Scholar] [CrossRef]
- Chevalier, S.; Bouffartigues, E.; Bodilis, J.; Maillot, O.; Lesouhaitier, O.; Feuilloley, M.G.; Orange, N.; Dufour, A.; Cornelis, P. Structure, function and regulation of Pseudomonas aeruginosa porins. FEMS Microbiol. Rev. 2017, 41, 698–722. [Google Scholar] [CrossRef]
- Hancock, R.E.; Speert, D.P. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and impact on treatment. Drug Resist. Updat. 2000, 3, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Livermore, D.M. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: Our worst nightmare? Clin. Infect. Dis. 2002, 34, 634–640. [Google Scholar] [CrossRef]
- Laudy, A.E.; Róg, P.; Smolińska-Król, K.; Ćmiel, M.; Słoczyńska, A.; Patzer, J.; Dzierżanowska, D.; Wolinowska, R.; Starościak, B.; Tyski, S. Prevalence of ESBL-producing Pseudomonas aeruginosa isolates in Warsaw, Poland, detected by various phenotypic and genotypic methods. PLoS ONE 2017, 12, e0180121. [Google Scholar] [CrossRef] [PubMed]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile beta-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef]
- Wright, L.L.; Turton, J.F.; Livermore, D.M.; Hopkins, K.L.; Woodford, N. Dominance of international ‘high-risk clones’ among metallo-beta-lactamase-producing Pseudomonas aeruginosa in the UK. J. Antimicrob. Chemother. 2015, 70, 103–110. [Google Scholar] [CrossRef]
- Kocsis, B.; Toth, A.; Gulyas, D.; Ligeti, B.; Katona, K.; Rokusz, L.; Szabo, D. Acquired qnrVC1 and blaNDM-1 resistance markers in an international high-risk Pseudomonas aeruginosa ST773 clone. J. Med. Microbiol. 2019, 68, 336–338. [Google Scholar] [CrossRef]
- Gaballah, A.; Elbaradei, A.; Elsheredy, A.; Kader, O. Emergence of blaVEB and blaGES among VIM-producing Pseudomonas aeruginosa clinical isolates in Alexandria, Egypt. Acta Microbiol. Immunol. Hung. 2019, 66, 131–142. [Google Scholar] [CrossRef]
- Del Barrio-Tofiño, E.; López-Causapé, C.; Oliver, A. Pseudomonas aeruginosa epidemic high-risk clones and their association with horizontally-acquired beta-lactamases: 2020 update. Int. J. Antimicrob. Agents 2020, 56, 106196. [Google Scholar] [CrossRef]
- Khalili, Y.; Yekani, M.; Goli, H.R.; Memar, M.Y. Characterization of carbapenem-resistant but cephalosporin-susceptible Pseudomonas aeruginosa. Acta Microbiol. Immunol. Hung. 2019, 66, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Gajdács, M. Carbapenem-Resistant but Cephalosporin-Susceptible Pseudomonas aeruginosa in Urinary Tract Infections: Opportunity for Colistin Sparing. Antibiotics 2020, 9, 53. [Google Scholar] [CrossRef]
- Poole, K. Aminoglycoside resistance in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2005, 49, 479–487. [Google Scholar] [CrossRef] [PubMed]
- Wachino, J.; Arakawa, Y. Exogenously acquired 16S rRNA methyltransferases found in aminoglycoside-resistant pathogenic Gram negative bacteria: An update. Drug Resist. Updat. 2012, 15, 133–148. [Google Scholar] [CrossRef] [PubMed]
- Galetti, R.; Andrade, L.N.; Varani, A.M.; Darinia, A.L.C. SPM-1-producing Pseudomonas aeruginosa ST277 carries chromosomal pack of acquired resistance genes: An example of high-risk clone associated to “intrinsic resistome”. J. Glob. Antimicrob. Resist. 2019, 16, 183–186. [Google Scholar] [CrossRef]
- Lee, J.Y.; Park, Y.K.; Chung, E.S.; Na, I.Y.; Ko, K.S. Evolved resistance to colistin and its loss due to genetic reversion in Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 25543. [Google Scholar] [CrossRef]
- Romano, K.P.; Warrier, T.; Poulsen, B.E.; Nguyen, P.H.; Loftis, A.R.; Saebi, A.; Pentelute, B.L.; Hung, D.T. Mutations in pmrB Confer Cross-Resistance between the LptD Inhibitor POL7080 and Colistin in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2019, 63, e00511-19. [Google Scholar] [CrossRef]
- Wi, Y.M.; Choi, J.Y.; Lee, J.Y.; Kang, C.I.; Chung, D.R.; Peck, K.R.; Song, J.H.; Ko, K.S. Emergence of colistin resistance in Pseudomonas aeruginosa ST235 clone in South Korea. Int. J. Antimicrob. Agents 2017, 49, 767–769. [Google Scholar] [CrossRef] [PubMed]
- Tahmasebi, H.; Dehbashi, S.; Arabestani, M.R. Co-harboring of mcr-1 and β-lactamase genes in Pseudomonas aeruginosa by high-resolution melting curve analysis (HRMA): Molecular typing of superbug strains in bloodstream infections (BSI). Infect. Genet. Evol. 2020, 85, 104518. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.J.; Lo, H.J. Molecular mechanisms of fluoroquinolone resistance. J. Microbiol. Immunol. Infect. 2003, 36, 1–9. [Google Scholar]
- Libisch, B.; Poirel, L.; Lepsanovic, Z.; Mirovic, V.; Balogh, B.; Pászti, J.; Hunyadi, Z.; Dobák, A.; Füzi, M.; Nordmann, P. Identification of PER-1 extended-spectrum beta-lactamase producing Pseudomonas aeruginosa clinical isolates of the international clonal complex CC11 from Hungary and Serbia. FEMS Immunol. Med. Microbiol. 2008, 54, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Botelho, J.; Grosso, F.; Peixe, L. ICEs are main reservoirs of ciprofloxacin modifying crpP gene in Pseudomonas aeruginosa. Genes 2020, 11, 889. [Google Scholar] [CrossRef]
- Curran, B.; Jonas, D.; Grundmann, H.; Pitt, T.; Dowson, C.G. Development of a multilocus sequence typing scheme for the opportunistic pathogen Pseudomonas aeruginosa. J. Clin. Microbiol. 2004, 42, 5644–5649. [Google Scholar] [CrossRef]
- De Sales, R.O.; Migliorini, L.B.; Puga, R.; Kocsis, B.; Severino, P. A Core Genome Multilocus Sequence Typing Scheme for Pseudomonas aeruginosa. Front. Microbiol. 2020, 11, 1049. [Google Scholar] [CrossRef] [PubMed]
- Woodford, N.; Turton, J.F.; Livermore, D.M. Multiresistant Gram-negative bacteria: The role of high-risk clones in the dissemination of antibiotic resistance. FEMS Microbiol. Rev. 2011, 35, 736–755. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.M.; Carmo, M.S.; Silbert, S.; Gales, A.C. SPM-1-producing Pseudomonas aeruginosa: Analysis of the ancestor relationship using multilocus sequence typing, pulsed-field gel electrophoresis, and automated ribotyping. Microb. Drug Resist. 2011, 17, 215–220. [Google Scholar] [CrossRef]
- Miyoshi-Akiyama, T.; Tada, T.; Ohmagari, N.; Viet Hung, N.; Tharavichitkul, P.; Pokhrel, B.M.; Gniadkowski, M.; Shimojima, M.; Kirikae, T. Emergence and Spread of Epidemic Multidrug-Resistant Pseudomonas aeruginosa. Genome Biol. Evol. 2017, 9, 3238–3245. [Google Scholar] [CrossRef]
- Bogaerts, P.; Bauraing, C.; Deplano, A.; Glupczynski, Y. Emergence and dissemination of BEL-1-producing Pseudomonas aeruginosa in Belgium. Antimicrob. Agents Chemother. 2007, 51, 1584–1585. [Google Scholar] [CrossRef]
- Botelho, J.; Grosso, F.; Sousa, C.; Peixe, L. Characterization of a new genetic environment associated with GES-6 carbapenemase from a Pseudomonas aeruginosa isolate belonging to the high-risk clone ST235. J. Antimicrob. Chemother. 2015, 70, 615–617. [Google Scholar] [CrossRef] [PubMed]
- Pollini, S.; Maradei, S.; Pecile, P.; Olivo, G.; Luzzaro, F.; Docquier, J.D.; Rossolini, G.M. FIM-1, a new acquired metallo-beta-lactamase from a Pseudomonas aeruginosa clinical isolate from Italy. Antimicrob. Agents Chemother. 2013, 57, 410–416. [Google Scholar] [CrossRef] [PubMed]
- Viedma, E.; Juan, C.; Acosta, J.; Zamorano, L.; Otero, J.R.; Sanz, F.; Chaves, F.; Oliver, A. Nosocomial spread of colistin-only-sensitive sequence type 235 Pseudomonas aeruginosa isolates producing the extended-spectrum beta-lactamases GES-1 and GES-5 in Spain. Antimicrob. Agents Chemother. 2009, 53, 4930–4933. [Google Scholar] [CrossRef]
- Juan, C.; Zamorano, L.; Mena, A.; Albertí, S.; Pérez, J.L.; Oliver, A. Metallo-beta-lactamase-producing Pseudomonas putida as a reservoir of multidrug resistance elements that can be transferred to successful Pseudomonas aeruginosa clones. J. Antimicrob. Chemother. 2010, 65, 474–478. [Google Scholar] [CrossRef] [PubMed]
- Larché, J.; Pouillot, F.; Essoh, C.; Libisch, B.; Straut, M.; Lee, J.C.; Soler, C.; Lamarca, R.; Gleize, E.; Gabard, J.; et al. Rapid identification of international multidrug-resistant Pseudomonas aeruginosa clones by multiple-locus variable number of tandem repeats analysis and investigation of their susceptibility to lytic bacteriophages. Antimicrob. Agents Chemother. 2012, 56, 6175–6180. [Google Scholar] [CrossRef]
- Pfennigwerth, N.; Geis, G.; Gatermann, S.G.; Kaase, M. Description of IMP-31, a novel metallo-beta-lactamase found in an ST235 Pseudomonas aeruginosa strain in Western Germany. J. Antimicrob. Chemother. 2015, 70, 1973–1980. [Google Scholar]
- Samuelsen, Ø.; Toleman, M.A.; Sundsfjord, A.; Rydberg, J.; Leegaard, T.M.; Walder, M.; Lia, A.; Ranheim, T.E.; Rajendra, Y.; Hermansen, N.O.; et al. Molecular Epidemiology of Metallo-beta -Lactamase-Producing Pseudomonas aeruginosa Isolates from Norway and Sweden Shows Import of International Clones and Local Clonal Expansion. Antimicrob. Agents Chemother. 2010, 54, 346–352. [Google Scholar] [CrossRef]
- Seok, Y.; Bae, I.K.; Jeong, S.H.; Kim, S.H.; Lee, H.; Lee, K. Dissemination of IMP-6 metallo-beta-lactamase-producing Pseudomonas aeruginosa sequence type 235 in Korea. J. Antimicrob. Chemother. 2011, 66, 2791–2796. [Google Scholar] [CrossRef]
- Hong, J.S.; Yoon, E.J.; Lee, H.; Jeong, S.H.; Lee, K. Clonal dissemination of Pseudomonas aeruginosa sequence type 235 isolates carrying blaIMP-6 and emergence of blaGES-24 and blaIMP-10 on novel genomic islands PAGI-15 and PAGI-16 in South Korea. Antimicrob. Agents Chemother. 2016, 60, 7216–7223. [Google Scholar] [PubMed]
- Mano, Y.; Saga, T.; Ishii, Y.; Yoshizumi, A.; Bonomo, R.A.; Yamaguchi, K.; Tateda, K. Molecular analysis of the integrons of metallo-beta-lactamase-producing Pseudomonas aeruginosa isolates collected by nationwide surveillance programs across Japan. BMC Microbiol. 2015, 21, 41. [Google Scholar]
- Treepong, P.; Kos, V.N.; Guyeux, C.; Blanc, D.S.; Bertrand, X.; Valot, B.; Hocquet, D. Global emergence of the widespread Pseudomonas aeruginosa ST235 clone. Clin. Microbiol. Infect. 2018, 24, 258. [Google Scholar] [CrossRef]
- Tada, T.; Nhung, P.H.; Miyoshi-Akiyama, T.; Shimada, K.; Tsuchiya, M.; Phuong, D.M.; Anh, N.Q.; Ohmagari, N.; Kirikae, T. Multidrug-Resistant Sequence Type 235 Pseudomonas aeruginosa Clinical Isolates Producing IMP-26 with Increased Carbapenem-Hydrolyzing Activities in Vietnam. Antimicrob. Agents Chemother. 2016, 60, 6853–6858. [Google Scholar] [CrossRef]
- Kim, M.J.; Bae, I.K.; Jeong, S.H.; Kim, S.H.; Song, J.H.; Choi, J.Y.; Yoon, S.S.; Thamlikitkul, V.; Hsueh, P.R.; Yasin, R.M.; et al. Dissemination of metallo-beta-lactamase-producing Pseudomonas aeruginosa of sequence type 235 in Asian countries. J. Antimicrob. Chemother. 2013, 68, 2820–2824. [Google Scholar] [CrossRef]
- Edelstein, M.V.; Skleenova, E.N.; Shevchenko, O.V.; D’souza, J.W.; Tapalski, D.V.; Azizov, I.S.; Sukhorukova, M.V.; Pavlukov, R.A.; Kozlov, R.S.; Toleman, M.A.; et al. Spread of extensively resistant VIM-2-positive ST235 Pseudomonas aeruginosa in Belarus, Kazakhstan, and Russia: A longitudinal epidemiological and clinical study. Lancet Infect. Dis. 2013, 13, 867–876. [Google Scholar] [CrossRef]
- Abril, D.; Marquez-Ortiz, R.A.; Castro-Cardozo, B.; Moncayo-Ortiz, J.I.; Olarte Escobar, N.M.; Corredor Rozo, Z.L.; Reyes, N.; Tovar, C.; Sánchez, H.F.; Castellanos, J.; et al. Genome plasticity favours double chromosomal Tn4401b-blaKPC-2 transposon insertion in the Pseudomonas aeruginosa ST235 clone. BMC Microbiol. 2019, 19, 45. [Google Scholar] [CrossRef]
- Vanegas, J.M.; Cienfuegos, A.V.; Ocampo, A.M.; López, L.; del Corral, H.; Roncancio, G.; Sierra, P.; Echeverri-Toro, L.; Ospina, S.; Maldonado, N.; et al. Similar frequencies of Pseudomonas aeruginosa isolates producing KPC and VIM carbapenemases in diverse genetic clones at tertiary-care hospitals in Medellin, Colombia. J. Clin. Microbiol. 2014, 52, 3978–3986. [Google Scholar] [CrossRef]
- De Paula-Petroli, S.B.; Campana, E.H.; Bocchi, M.; Bordinhão, T.; Picão, R.C.; Yamada-Ogatta, S.F.; Carrara-Marroni, F.E. Early detection of a hypervirulent KPC-2-producing Pseudomonas aeruginosa ST235 in Brazil. J. Glob. Antimicrob. Resist. 2018, 12, 153–154. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.; Beceiro, A.; Gutiérrez, O.; Albertí, S.; Garau, M.; Pérez, J.L.; Bou, G.; Oliver, A. Characterization of the new metallo-beta-lactamase VIM-13 and its integron-borne gene from a Pseudomonas aeruginosa clinical isolate in Spain. Antimicrob. Agents Chemother. 2008, 52, 3589–3596. [Google Scholar] [CrossRef] [PubMed]
- Zamorano, L.; Mulet, X.; Garau, M.; Gallegos, C.; Juan, C.; Pérez, J.L.; Oliver, A. VIM-47, a New Variant of the Autochthonous Metallo-beta-Lactamase VIM-13 from the Balearic Islands in Spain. Antimicrob. Agents Chemother. 2016, 60, 3251–3252. [Google Scholar] [CrossRef] [PubMed]
- Janvier, F.; Jeannot, K.; Tessé, S.; Robert-Nicoud, M.; Delacour, H.; Rapp, C.; Mérens, A. Molecular characterization of blaNDM-1 in a sequence type 235 Pseudomonas aeruginosa isolate from France. Antimicrob. Agents Chemother. 2013, 57, 3408–3411. [Google Scholar] [CrossRef] [PubMed]
- Loconsole, D.; Accogli, M.; Monaco, M.; Del Grosso, M.; De Robertis, A.L.; Morea, A.; Capozzi, L.; Del Sambro, L.; Simone, A.; De Letteriis, V.; et al. First detection of autochthonous extensively drug-resistant NDM-1 Pseudomonas aeruginosa ST235 from a patient with bloodstream infection in Italy, October 2019. Antimicrob. Resist. Infect. Control. 2020, 9, 73. [Google Scholar] [CrossRef] [PubMed]
- Recio, R.; Villa, J.; Viedma, E.; Orellana, M.Á.; Lora-Tamayo, J.; Chaves, F. Bacteraemia due to extensively drug-resistant Pseudomonas aeruginosa sequence type 235 high-risk clone: Facing the perfect storm. Int. J. Antimicrob. Agents. 2018, 52, 172–179. [Google Scholar] [CrossRef]
- Witney, A.A.; Gould, K.A.; Pope, C.F.; Bolt, F.; Stoker, N.G.; Cubbon, M.D.; Bradley, C.R.; Fraise, A.; Breathnach, A.S.; Butcher, P.D.; et al. Genome sequencing and characterization of an extensively drug-resistant sequence type 111 serotype O12 hospital outbreak strain of Pseudomonas aeruginosa. Clin. Microbiol. Infect. 2014, 20, O609–O618. [Google Scholar] [CrossRef] [PubMed]
- Guzvinec, M.; Izdebski, R.; Butic, I.; Jelic, M.; Abram, M.; Koscak, I.; Baraniak, A.; Hryniewicz, W.; Gniadkowski, M.; Andrasevic, A.T. Sequence types 235, 111, and 132 predominate among multidrug-resistant Pseudomonas aeruginosa clinical isolates in Croatia. Antimicrob. Agents Chemother. 2014, 58, 6277–6283. [Google Scholar] [CrossRef] [PubMed]
- Papagiannitsis, C.C.; Studentova, V.; Ruzicka, F.; Tejkalova, R.; Hrabak, J. Molecular characterization of metallo-b-lactamase-producing Pseudomonas aeruginosa in a Czech hospital (2009–2011). J. Med. Microbiol. 2013, 62, 945–947. [Google Scholar] [CrossRef][Green Version]
- Vatcheva-Dobrevska, R.; Mulet, X.; Ivanov, I.; Zamorano, L.; Dobreva, E.; Velinov, T.; Kantardjiev, T.; Oliver, A. Molecular epidemiology and multidrug resistance mechanisms of Pseudomonas aeruginosa isolates from Bulgarian hospitals. Microb. Drug Resist. 2013, 19, 355–361. [Google Scholar] [CrossRef]
- Van der Bij, A.K.; Van der Zwan, D.; Peirano, G.; Severin, J.A.; Pitout, J.D.; Van Westreenen, M.; Goessens, W.H. Metallo-beta-lactamase producing Pseudomonas aeruginosa in the Netherlands: The nationwide emergence of a single sequence type. Clin. Microbiol. Infect. 2012, 18, E369–E372. [Google Scholar] [CrossRef]
- Turton, J.F.; Wright, L.; Underwood, A.; Witney, A.A.; Chan, Y.T.; Al-Shahib, A.; Arnold, C.; Doumith, M.; Patel, B.; Planche, T.D.; et al. High-Resolution Analysis by Whole-Genome Sequencing of an International Lineage (Sequence Type 111) of Pseudomonas aeruginosa Associated with Metallo-Carbapenemases in the United Kingdom. J. Clin. Microbiol. 2015, 53, 2622–2631. [Google Scholar] [CrossRef] [PubMed]
- Arca-Suárez, J.; Fraile-Ribot, P.; Vázquez-Ucha, J.C.; Cabot, G.; Martínez-Guitián, M.; Lence, E.; González-Bello, C.; Beceiro, A.; Rodríguez-Iglesias, M.; Galán-Sánchez, F.; et al. Challenging Antimicrobial Susceptibility and Evolution of Resistance (OXA-681) during Treatment of a Long-Term Nosocomial Infection Caused by a Pseudomonas aeruginosa ST175 Clone. Antimicrob. Agents Chemother. 2019, 63, e01110-19. [Google Scholar] [CrossRef]
- Cabot, G.; López-Causapé, C.; Ocampo-Sosa, A.A.; Sommer, L.M.; Domínguez, M.A.; Zamorano, L.; Juan, C.; Tubau, F.; Rodríguez, C.; Moyà, B.; et al. Deciphering the Resistome of the Widespread Pseudomonas aeruginosa Sequence Type 175 International High-Risk Clone through Whole-Genome Sequencing. Antimicrob. Agents Chemother. 2016, 60, 7415–7423. [Google Scholar]
- Dößelmann, B.; Willmann, M.; Steglich, M.; Bunk, B.; Nübel, U.; Peter, S.; Neher, R.A. Rapid and Consistent Evolution of Colistin Resistance in Extensively Drug-Resistant Pseudomonas aeruginosa during Morbidostat Culture. Antimicrob. Agents Chemother. 2017, 61, e00043-17. [Google Scholar] [CrossRef] [PubMed]
- Taiaroa, G.; Samuelsen, Ø.; Kristensen, T.; Økstad, O.A.L.; Heikal, A. Complete Genome Sequence of Pseudomonas aeruginosa K34-7, a Carbapenem-Resistant Isolate of the High-Risk Sequence Type 233. Microbiol. Resour. Announc. 2018, 7, e00886-18. [Google Scholar] [CrossRef]
- Silveira, M.; Albano, R.; Asensi, M.; Assef, A.P. The draft genome sequence of multidrug-resistant Pseudomonas aeruginosa strain CCBH4851, a nosocomial isolate belonging to clone SP (ST277) that is prevalent in Brazil. Mem. Inst. Oswaldo. Cruz. 2014, 109, 1086–1087. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Khan, A.; Shropshire, W.C.; Hanson, B.; Dinh, A.Q.; Wanger, A.; Ostrosky-Zeichner, L.; Arias, C.A.; Miller, W.R. Simultaneous Infection with Enterobacteriaceae and Pseudomonas aeruginosa Harboring Multiple Carbapenemases in a Returning Traveler Colonized with Candida auris. Antimicrob. Agents Chemother. 2020, 64, e01466-19. [Google Scholar] [CrossRef] [PubMed]
- Correa, A.; Montealegre, M.C.; Mojica, M.F.; Maya, J.J.; Rojas, L.J.; De La Cadena, E.P.; Ruiz, S.J.; Recalde, M.; Rosso, F.; Quinn, J.P.; et al. First report of a Pseudomonas aeruginosa isolate coharboring KPC and VIM carbapenemases. Antimicrob. Agents Chemother. 2012, 56, 5422–5423. [Google Scholar] [CrossRef]
- Castanheira, M.; Toleman, M.A.; Jones, R.N.; Schmidt, F.J.; Walsh, T.R. Molecular characterization of a beta-lactamase gene, blaGIM-1, encoding a new subclass of metallo-beta-lactamase. Antimicrob. Agents Chemother. 2004, 48, 4654–4661. [Google Scholar] [CrossRef]
- Wendel, A.F.; Kolbe-Busch, S.; Ressina, S.; Schulze-Röbbecke, R.; Kindgen-Milles, D.; Lorenz, C.; Pfeffer, K.; MacKenzie, C.R. Detection and termination of an extended low-frequency hospital outbreak of GIM-1-producing Pseudomonas aeruginosa ST111 in Germany. Am. J. Infect. Control. 2015, 43, 635–639. [Google Scholar] [CrossRef]
- Viedma, E.; Juan, C.; Villa, J.; Barrado, L.; Orellana, M.A.; Sanz, F.; Otero, J.R.; Oliver, A.; Chaves, F. VIM-2-producing multidrug-resistant Pseudomonas aeruginosa ST175 clone, Spain. Emerg. Infect. Dis. 2012, 18, 1235–1241. [Google Scholar] [CrossRef]
- López-Causapé, C.; Cabot, G.; Del Barrio-Tofiño, E.; Oliver, A. The Versatile Mutational Resistome of Pseudomonas aeruginosa. Front. Microbiol. 2018, 9, 685. [Google Scholar] [CrossRef]
- Perez-Vazquez, M.; Sola-Campoy, P.J.; Zurita, Á.M.; Avila, A.; Gomez-Bertomeu, F.; Solis, S.; Lopez-Urrutia, L.; Gónzalez-Barberá, E.M.; Cercenado, E.; Bautista, V.; et al. Carbapenemase-producing Pseudomonas aeruginosa in Spain: Interregional dissemination of the high-risk clones ST175 and ST244 carrying blaVIM-2, blaVIM-1, blaIMP-8, blaVIM-20 and blaKPC-2. Int. J. Antimicrob. Agents 2020, 56, 106026. [Google Scholar] [CrossRef]
- Aguilar-Rodea, P.; Zúñiga, G.; Rodríguez-Espino, B.A.; Olivares Cervantes, A.L.; Gamiño Arroyo, A.E.; Moreno-Espinosa, S.; de la Rosa Zamboni, D.; López Martínez, B.; Castellanos-Cruz, M.d.C.; Parra-Ortega, I.; et al. Identification of extensive drug resistant Pseudomonas aeruginosa strains: New clone ST1725 and high-risk clone ST233. PLoS ONE 2017, 12, e0172882. [Google Scholar] [CrossRef] [PubMed]
- Perez, F.; Hujer, A.M.; Marshall, S.H.; Ray, A.J.; Rather, P.N.; Suwantarat, N.; Dumford, D., 3rd; O’Shea, P.; Domitrovic, T.N.J.; Salata, R.A.; et al. Extensively drug-resistant Pseudomonas aeruginosa isolates containing blaVIM-2 and elements of Salmonella genomic island 2: A new genetic resistance determinant in Northeast Ohio. Antimicrob. Agents Chemother. 2014, 58, 5929–5935. [Google Scholar] [CrossRef]
- Mudau, M.; Jacobson, R.; Minenza, N.; Kuonza, L.; Morris, V.; Engelbrecht, H.; Nicol, M.P.; Bamford, C. Outbreak of multi-drug resistant Pseudomonas aeruginosa bloodstream infection in the haematology unit of a South African Academic Hospital. PLoS ONE 2013, 8, e55985. [Google Scholar] [CrossRef]
- Koh, T.H.; Khoo, C.T.; Tan, T.T.; Arshad, M.A.; Ang, L.P.; Lau, L.J.; Hsu, L.Y.; Ooi, E.E. Multilocus sequence types of carbapenem-resistant Pseudomonas aeruginosa in Singapore carrying metallo-beta-lactamase genes, including the novel blaIMP-26 gene. J. Clin. Microbiol. 2010, 48, 2563–2564. [Google Scholar] [CrossRef]
- Zafer, M.M.; Al-Agamy, M.H.; El-Mahallawy, H.A.; Amin, M.A.; El Din Ashour, S. Dissemination of VIM-2 producing Pseudomonas aeruginosa ST233 at tertiary care hospitals in Egypt. BMC Infect. Dis. 2015, 15, 122. [Google Scholar] [CrossRef]
- Fernandes, M.R.; Sellera, F.P.; Moura, Q.; Carvalho, M.; Rosato, P.N.; Cerdeira, L.; Lincopan, N. Zooanthroponotic Transmission of Drug-Resistant Pseudomonas aeruginosa, Brazil. Emerg. Infect. Dis. 2018, 24, 1160–1162. [Google Scholar] [CrossRef] [PubMed]
- Silveira, M.C.; Albano, R.M.; Asensi, M.D.; Carvalho-Assef, A.P. Description of genomic islands associated to the multidrug-resistant Pseudomonas aeruginosa clone ST277. Infect. Genet Evol. 2016, 42, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, K.L.; Meunier, D.; Findlay, J.; Mustafa, N.; Parsons, H.; Pike, R.; Wright, L.; Woodford, N. SPM-1 metallo-beta-lactamase-producing Pseudomonas aeruginosa ST277 in the UK. J. Med. Microbiol. 2016, 65, 696–697. [Google Scholar] [CrossRef]
- Silveira, M.C.; Rocha-de-Souza, C.M.; Albano, R.M.; de Oliveira Santos, I.C.; Carvalho-Assef, A.P. Exploring the success of Brazilian endemic clone Pseudomonas aeruginosa ST277 and its association with the CRISPR-Cas system type I-C. BMC Genomics 2020, 21, 255. [Google Scholar] [CrossRef]
- Martins, W.M.B.S.; Narciso, A.C.; Cayô, R.; Santos, S.V.; Fehlberg, L.C.C.; Ramos, P.L.; da Cruz, J.B.; Gales, A.C. SPM-1-producing Pseudomonas aeruginosa ST277 clone recovered from microbiota of migratory birds. Diagn Microbiol. Infect. Dis. 2018, 90, 221–227. [Google Scholar] [CrossRef]
- Turano, H.; Gomes, F.; Medeiros, M.; Oliveira, S.; Fontes, L.C.; Sato, M.I.; Lincopan, N. Presence of high-risk clones of OXA-23-producing Acinetobacter baumannii (ST79) and SPM-1-producing Pseudomonas aeruginosa (ST277) in environmental water samples in Brazil. Diagn. Microbiol. Infect. Dis. 2016, 86, 80–82. [Google Scholar] [CrossRef] [PubMed]
- Papagiannitsis, C.C.; Medvecky, M.; Chudejova, K.; Skalova, A.; Rotova, V.; Spanelova, P.; Jakubu, V.; Zemlickova, H.; Hrabak, J. Molecular Characterization of Carbapenemase-Producing Pseudomonas aeruginosa of Czech Origin and Evidence for Clonal Spread of Extensively Resistant Sequence Type 357 Expressing IMP-7 Metallo-β-Lactamase. Antimicrob. Agents Chemother. 2017, 61, e01811-1. [Google Scholar] [CrossRef] [PubMed]
- Hrabák, J.; Cervená, D.; Izdebski, R.; Duljasz, W.; Gniadkowski, M.; Fridrichová, M.; Urbásková, P.; Zemlicková, H. Regional spread of Pseudomonas aeruginosa ST357 producing IMP-7 metallo-β-lactamase in Central Europe. J. Clin. Microbiol. 2011, 49, 474–475. [Google Scholar] [CrossRef]
- Kainuma, A.; Momiyama, K.; Kimura, T.; Akiyama, K.; Inoue, K.; Naito, Y.; Kinoshita, M.; Shimizu, M.; Kato, H.; Shime, N.; et al. An outbreak of fluoroquinolone-resistant Pseudomonas aeruginosa ST357 harboring the exoU gene. J. Infect. Chemother. 2018, 24, 615–622. [Google Scholar] [CrossRef]
- Al-Zahrani, I.A.; Al-Ahmadi, B.M. Dissemination of VIM-producing Pseudomonas aeruginosa associated with high-risk clone ST654 in a tertiary and quaternary hospital in Makkah, Saudi Arabia. J. Chemother. 2021, 33, 12–20. [Google Scholar] [CrossRef]
- Pasteran, F.; Faccone, D.; Gomez, S.; De Bunder, S.; Spinelli, F.; Rapoport, M.; Petroni, A.; Galas, M.; Corso, A. Detection of an international multiresistant clone belonging to sequence type 654 involved in the dissemination of KPC-producing Pseudomonas aeruginosa in Argentina. J. Antimicrob. Chemother. 2012, 67, 1291–1293. [Google Scholar] [CrossRef]
- Yousefi, S.; Nahaei, M.R.; Farajnia, S.; Aghazadeh, M.; Iversen, A.; Edquist, P.; Maãtallah, M.; Giske, C.G. A multiresistant clone of Pseudomonas aeruginosa sequence type 773 spreading in a burn unit in Orumieh, Iran. APMIS 2013, 121, 146–152. [Google Scholar] [CrossRef]
- Chmielarczyk, A.; Pobiega, M.; Ziółkowski, G.; Pomorska-Wesołowska, M.; Romaniszyn, D.; Krawczyk, L.; Wójkowska-Mach, J. Severe infections caused by multidrug-resistant non-fermentative bacilli in southern Poland. Adv. Clin. Exp. Med. 2018, 27, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Papagiannitsis, C.C.; Verra, A.; Galani, V.; Xitsas, S.; Bitar, I.; Hrabak, J.; Petinaki, E. Unravelling the Features of Success of VIM-Producing ST111 and ST235 Pseudomonas aeruginosa in a Greek Hospital. Microorganisms 2020, 8, 1884. [Google Scholar] [CrossRef]
- Nascimento, A.P.; Ortiz, M.F.; Martins, W.M.; Morais, G.L.; Fehlberg, L.C.; Almeida, L.G.; Ciapina, L.P.; Gales, A.C.; Vasconcelos, A.T. Intraclonal Genome Stability of the Metallo-beta-lactamase SPM-1-producing Pseudomonas aeruginosa ST277, an Endemic Clone Disseminated in Brazilian Hospitals. Front. Microbiol. 2016, 7, 1946. [Google Scholar] [CrossRef]
- Botelho, J.; Roberts, A.P.; Leon-Sampedro, R.; Grosso, F.; Peixe, L. Carbapenemases on the move: It’s good to be on ICEs. Mob. DNA 2018, 9, 37. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Reference | Clone | Accession Number |
|---|---|---|
| Loconsole et al., 2020 [61] | ST235 | JAABOY000000000 |
| Tada et al., 2016 [52] | ST235, ST277 | (SRA): DRA003741 |
| Witney et al., 2014 [63] | ST111 | PRJEB4573 |
| Kos et al., 2015 [15] | ST111 | JUAZ00000000 |
| Turton et al., 2015 [68] | ST111 | ERS716506 to ERS716591 |
| Arca-Suárez et al., 2019 [69] | ST175 | SRX5389644 |
| Cabot et al., 2016 [70] | ST175 | ERS1280254 to ERS1280271 and ERS1280273 to ERS1280276 (European Nucleotide Archive) |
| Dößelmann et al., 2017 [71] | ST233 | CP017293 and CP017294 |
| Taiaroa et al., 2018 [72] | ST233 | CP029707 CP029708 |
| Silveira et al., 2014 [73] | ST277 | JPSS00000000 |
| Galetti et al., 2019 [27] | ST277 | MVOB00000000 |
| Kos et al., 2015 [15] | ST654 | JTYC00000000 |
| Kocsis et al., 2019 [20] | ST773 | RHDU00000000 |
| Khan et al., 2020 [74] | ST773 | BioSample: SAMN12307670 |
| The Most Common Clinically Relevant Resistance Genes in P. aeruginosa | |||||
|---|---|---|---|---|---|
| Clone | Beta-Lactamases | Aminoglycoside Modifying Enzymes | Resistance to Fluoroquinolones | Resistance to Colistin | Other |
| ST235 | blaPSE-1, blaBEL-1, blaPER-1, blaGES-variants, blaCTX-M, blaOXA-2, blaKPC-2, blaPAO, blaIMP-variants, blaVIM-2 and variants, blaFIM-1, blaNDM-1 | aacA7, aph(6)-Id, aadA1, aac(6′)-Ib-cr | qnrVC1, mutations in gyrA and parC | Strains susceptible to colistin. Sporadic reports of mcr carriage | fosA, cmlA7, sul-1, tetA, tetGcatB7, |
| ST111 | blaPSE-1, blaGES-7, blaVIM-2, blaKPC-2, blaIMP-1 and variants, blaVEB-1, blaOXA-2, and variants blaNDM-1 | aac(6)-Ib, aacA29a | mutations in gyrA, parC, or parE | Colistin-resistant strain is confirmed | mexZ efflux pump, oprD, sul-1, catB7 |
| ST175 | blaCARB-12, blaIMP-1, blaVIM-2, blaTEM-1A, blaAER1, blaOXA-2 variants, like blaOXA-681 | aac(6)-1b, aacA4′-3 | qnrVC-1, mutations in gyrA and parC | No data available | oprD, ampR, mexZ efflux pumps, catB7 |
| ST233 | blaVIM-2, blaIMP-1, blaOXA-4, blaOXA-50, blaPDC-3, blaPAO, blaNDM-1 | aac(3)-Id, aadA2, aph(3′)-IIb | mutations in gyrA and parC | Colistin-resistant strain is confirmed | dfrB5, fosA, sul1, tetG, catB7, cmlA1 |
| ST277 | blaSPM-1, blaIMP-1, blaOXA-56, blaPAO | aacA1, aacA4, aac(6)-Ib, aac(6)-Ib-cr, aph(3′)-IIb, aadA7 | mutations in gyrA and parC | No data available | fosA, sul-1, catB7 |
| ST357 | blaPER-1, blaGES-5, blaLCR-1, blaIMP-1 variants, blaVIM-2, blaOXA-2 variants, blaVEB1-like | aadA11, aadA2, aadB, aacA3, aacA4, aph(3)-VIa, aac(6)-Ib, aph(6)-Id | mutations in gyrA and parC | No data available | fos1, fosE, sul-1 |
| ST654 | blaKPC-2, blaIMP-1, blaVIM-2, blaNDM-1 | aacA4, aacA5, aadb | mutations in gyrA and parC | No data available | sul-1 |
| ST773 | blaOXA-50-like variants, blaVEB-1, blaIMP-2, blaPAO, blaVIM-2, blaNDM-1 | rmtB, aph(3′)-Iib, aac(3)-Ia, aadA1, aadB | qnrVC1, mutations in gyrA and parC | Strains susceptible only to colistin are detected | fosA, sul-1, tetG, catB7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocsis, B.; Gulyás, D.; Szabó, D. Diversity and Distribution of Resistance Markers in Pseudomonas aeruginosa International High-Risk Clones. Microorganisms 2021, 9, 359. https://doi.org/10.3390/microorganisms9020359
Kocsis B, Gulyás D, Szabó D. Diversity and Distribution of Resistance Markers in Pseudomonas aeruginosa International High-Risk Clones. Microorganisms. 2021; 9(2):359. https://doi.org/10.3390/microorganisms9020359
Chicago/Turabian StyleKocsis, Béla, Dániel Gulyás, and Dóra Szabó. 2021. "Diversity and Distribution of Resistance Markers in Pseudomonas aeruginosa International High-Risk Clones" Microorganisms 9, no. 2: 359. https://doi.org/10.3390/microorganisms9020359
APA StyleKocsis, B., Gulyás, D., & Szabó, D. (2021). Diversity and Distribution of Resistance Markers in Pseudomonas aeruginosa International High-Risk Clones. Microorganisms, 9(2), 359. https://doi.org/10.3390/microorganisms9020359