
Health-Promoting Role of Lactiplantibacillus plantarum Isolated from Fermented Foods




Abstract
1. Introduction
2. Selection Criteria for Health-Promoting Bacteria
3. Genomic Insight into Food-Borne Lpb. plantarum Species
3.1. Production of Exopolysaccharides
3.2. Production of Bacteriocins
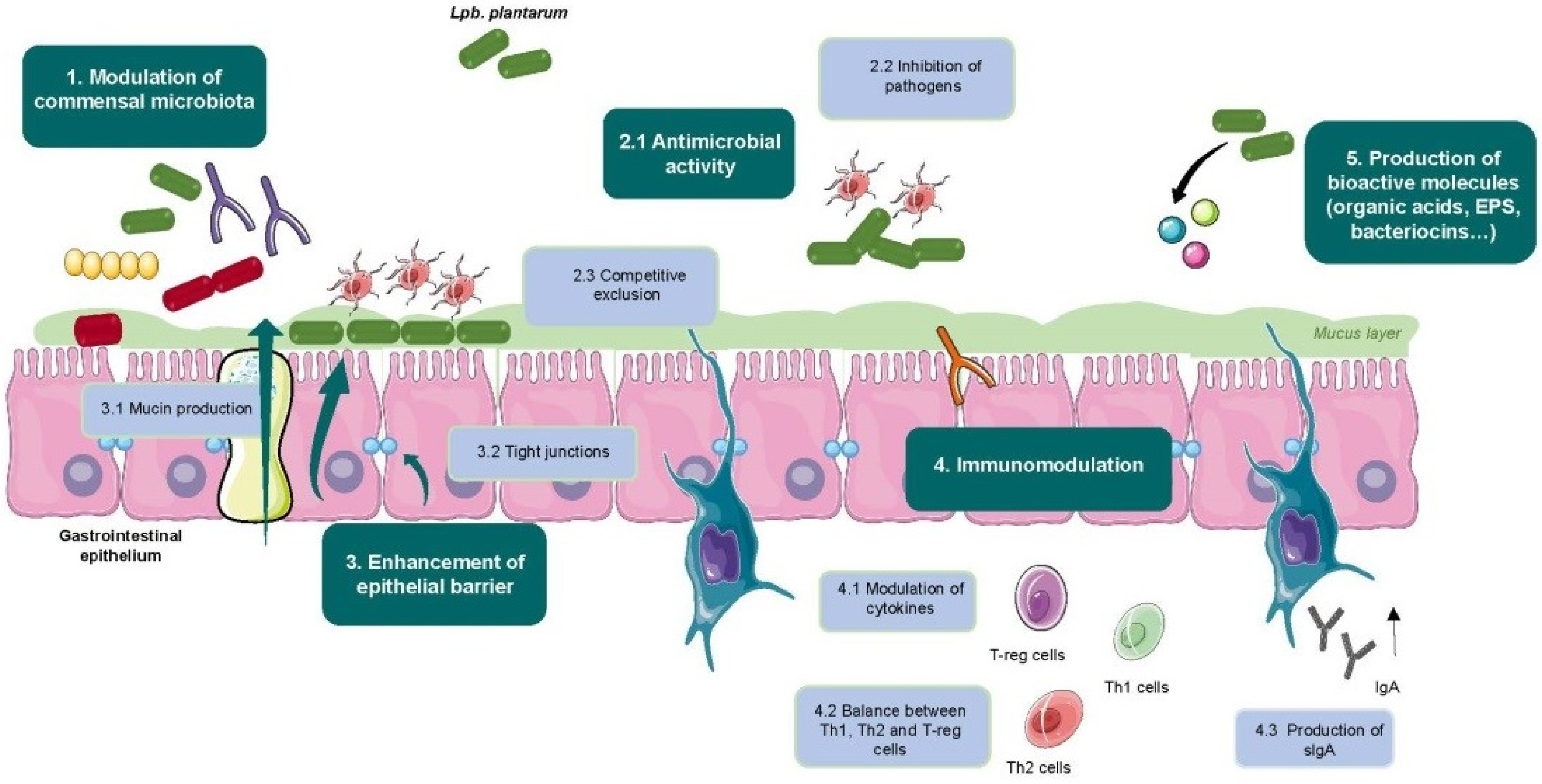
4. Health Benefits of Food-Associated Lpb. plantarum Strains
4.1. Antimicrobial Activity
4.2. Antigenotoxic and Antimutagenic Activity
4.3. Bile Salt Hydrolase Activity
4.4. Antioxidant Properties
4.5. Immune Modulation
5. Clinical Studies
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pasolli, E.; De Filippis, F.; Mauriello, I.E.; Cumbo, F.; Walsh, A.M.; Leech, J.; Cotter, P.D.; Segata, N.; Ercolini, D. Large-scale genome-wide analysis links lactic acid bacteria from food with the gut microbiome. Nat. Commun. 2020, 11, 2610. [Google Scholar] [CrossRef]
- Marco, M.L.; Heeney, D.; Binda, S.; Cifelli, C.J.; Cotter, P.D.; Foligné, B.; Gänzle, M.; Kort, R.; Pasin, G.; Pihlanto, A.; et al. Health benefits of fermented foods: Microbiota and beyond. Curr. Opin. Biotechnol. 2017, 44, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Marco, M.L.; Hill, C.; Hutkins, R.; Slavin, J.; Tancredi, D.J.; Merenstein, D.; Sanders, M.E. Should There Be a Recommended Daily Intake of Microbes? J. Nutr. 2020, 150, 3061–3067. [Google Scholar] [CrossRef] [PubMed]
- Kok, C.R.; Hutkins, R. Yogurt and other fermented foods as sources of health-promoting bacteria. Nutr. Rev. 2018, 76, 4–15. [Google Scholar] [CrossRef]
- Rezac, S.; Kok, C.R.; Heermann, M.; Hutkins, R. Fermented foods as a dietary source of live organisms. Front. Microbiol. 2018, 9, 1785. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wittouck, S.; Salvetti, E.; Franz, C.M.A.P.; Harris, H.M.B.; Mattarelli, P.; O’Toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef] [PubMed]
- Corsetti, A.; Prete, R.; Garcia-Gonzalez, N. Lactic Acid Bacteria: Lactobacillus spp. Lactobacillus Plantarum; Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Behera, S.S.; Ray, R.C.; Zdolec, N. Lactobacillus plantarum with functional properties: An approach to increase safety and shelf-life of fermented foods. Biomed. Res. Int. 2018, 2018, 9361614. [Google Scholar] [CrossRef]
- Aponte, M.; Blaiotta, G.; Croce, F.L.; Mazzaglia, A.; Farina, V.; Settanni, L.; Moschetti, G. Use of selected autochthonous lactic acid bacteria for Spanish-style table olive fermentation. Food Microbiol. 2012, 30, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Perpetuini, G.; Prete, R.; Garcia-Gonzalez, N.; Khairul Alam, M.; Corsetti, A. Table olives more than a fermented food. Foods 2020, 9, 178. [Google Scholar] [CrossRef]
- Food and Agriculture Organization; World Health Organization. Probiotics in Food, Health and Nutritional Properties and Guidelines for Evaluation; FAO Food Nutrition: Rome, Italy, 2006. [Google Scholar]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef]
- Binda, S.; Hill, C.; Johansen, E.; Obis, D.; Pot, B.; Sanders, M.E.; Tremblay, A.; Ouwehand, A.C. Criteria to qualify microorganisms as ”Probiotic” in foods and dietary supplements. Front. Microbiol. 2020, 11, 1662. [Google Scholar] [CrossRef] [PubMed]
- Nuraida, L. A review: Health promoting lactic acid bacteria in traditional Indonesian fermented foods. Food Sci. Hum. Wellness 2015, 4, 47–55. [Google Scholar] [CrossRef]
- Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; Bastos, M.L.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; Gropp, J.; et al. Guidance on the characterisation of microorganisms used as feed additives or as production organisms. EFSA J. 2018, 16, e05206. [Google Scholar] [CrossRef] [PubMed]
- Seddik, H.A.; Bendali, F.; Gancel, F.; Fliss, I.; Spano, G.; Drider, D. Lactobacillus plantarum and its probiotic and food potentialities. Probiotics Antimicrob. Proteins 2017, 9, 111–122. [Google Scholar] [CrossRef]
- Upadrasta, A.; Stanton, C.; Hill, C.; Fitzgerald, G.; Ross, R. Stress Responses of Lactic Acid Bacteria, 1st ed.; Papadimitriou, E.T.A.K., Ed.; Springer: New York, NY, USA, 2011. [Google Scholar]
- Liu, Q.; Yu, Z.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Surface components and metabolites of probiotics for regulation of intestinal epithelial barrier. Microb. Cell Fact. 2020, 19, 23. [Google Scholar] [CrossRef] [PubMed]
- Odenwald, M.A.; Turner, J.R. The intestinal epithelial barrier: A therapeutic target? Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 9–21. [Google Scholar] [CrossRef]
- Derrien, M.; van Hylckama Vlieg, J.E. Fate, activity, and impact of ingested bacteria within the human gut microbiota. Trends Microbiol. 2015, 23, 354–366. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Boekhorst, J.; van Kranenburg, R.; Molenaar, D.; Kuipers, O.P.; Leer, R.; Tarchini, R.; Peters, S.A.; Sandbrink, H.M.; Fiers, M.W.; et al. Complete genome sequence of Lactobacillus plantarum WCFS1. Proc. Natl. Acad. Sci. USA 2003, 100, 1990–1995. [Google Scholar] [CrossRef] [PubMed]
- Evanovich, E.; de Souza Mendonça Mattos, P.J.; Guerreiro, J.F. Comparative Genomic Analysis of Lactobacillus plantarum: An Overview. Int. J. Genomics 2019, 2019, 4973214. [Google Scholar] [CrossRef]
- Siezen, R.J.; Tzeneva, V.A.; Castioni, A.; Wels, M.; Phan, H.T.; Rademaker, J.L.; Starrenburg, M.J.; Kleerebezem, M.; Molenaar, D.; van Hylckama Vlieg, J.E. Phenotypic and genomic diversity of Lactobacillus plantarum strains isolated from various environmental niches. Environ. Microbiol. 2010, 12, 758–773. [Google Scholar] [CrossRef]
- Siezen, R.J.; Francke, C.; Renckens, B.; Boekhorst, J.; Wels, M.; Kleerebezem, M.; van Hijum, S.A. Complete resequencing and reannotation of the Lactobacillus plantarum WCFS1 genome. J. Bacteriol. 2012, 194, 195–196. [Google Scholar] [CrossRef]
- Crawley, A.B.; Henriksen, E.D.; Stout, E.; Brandt, K.; Barrangou, R. Characterizing the activity of abundant, diverse and active CRISPR-Cas systems in lactobacilli. Sci. Rep. 2018, 8, 11544. [Google Scholar] [CrossRef] [PubMed]
- Sanilbaba, P.; Cakmak, G. Exopolysaccharides production by lactic acid bacteria. App. Microbiol. 2016, 2, 451–469. [Google Scholar] [CrossRef]
- Kelleher, P.; Murphy, J.; Mahony, J.; van Sinderen, D. Next-generation sequencing as an approach to dairy starter selection. Dairy Sci. Technol. 2015, 95, 545–568. [Google Scholar] [CrossRef] [PubMed]
- Nwodo, U.U.; Green, E.; Okoh, A.I. Bacterial exopolysaccharides: Functionality and prospects. Int. J. Mol. Sci. 2012, 13, 14002–14015. [Google Scholar] [CrossRef] [PubMed]
- Limoli, D.H.; Jones, C.J.; Wozniak, D.J. Bacterial extracellular polysaccharides in biofilm formation and function. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [PubMed]
- Deo, D.; Davray, D.; Kulkarni, R. A Diverse repertoire of exopolysaccharide biosynthesis gene clusters in. Microorganisms 2019, 7, 444. [Google Scholar] [CrossRef]
- Remus, D.M.; van Kranenburg, R.; van Swam, I.I.; Taverne, N.; Bongers, R.S.; Wels, M.; Wells, J.M.; Bron, P.A.; Kleerebezem, M. Impact of 4 Lactobacillus plantarum capsular polysaccharide clusters on surface glycan composition and host cell signaling. Microb. Cell Fact. 2012, 11, 149. [Google Scholar] [CrossRef]
- Lee, I.C.; Caggianiello, G.; van Swam, I.I.; Taverne, N.; Meijerink, M.; Bron, P.A.; Spano, G.; Kleerebezem, M. Strain-specific features of extracellular polysaccharides and their impact on Lactobacillus plantarum-host interactions. Appl. Environ. Microbiol. 2016, 82, 3959–3970. [Google Scholar] [CrossRef]
- Lamontanara, A.; Caggianiello, G.; Orrù, L.; Capozzi, V.; Michelotti, V.; Bayjanov, J.R.; Renckens, B.; van Hijum, S.A.; Cattivelli, L.; Spano, G. Draft genome sequence of Lactobacillus plantarum Lp90 Isolated from wine. Genome Announc. 2015, 3. [Google Scholar] [CrossRef]
- Li, P.; Li, X.; Gu, Q.; Lou, X.Y.; Zhang, X.M.; Song, D.F.; Zhang, C. Comparative genomic analysis of Lactobacillus plantarum ZJ316 reveals its genetic adaptation and potential probiotic profiles. J. Zhejiang Univ. Sci. B 2016, 17, 569–579. [Google Scholar] [CrossRef]
- Torino, M.I.; Font de Valdez, G.; Mozzi, F. Biopolymers from lactic acid bacteria. Novel applications in foods and beverages. Front. Microbiol. 2015, 6, 834. [Google Scholar] [CrossRef] [PubMed]
- Angelin, J.; Kavitha, M. Exopolysaccharides from probiotic bacteria and their health potential. Int. J. Biol. Macromol. 2020, 162, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Ayyash, M.; Abu-Jdayil, B.; Itsaranuwat, P.; Galiwango, E.; Tamiello-Rosa, C.; Abdullah, H.; Esposito, G.; Hunashal, Y.; Obaid, R.S.; Hamed, F. Characterization, bioactivities, and rheological properties of exopolysaccharide produced by novel probiotic Lactobacillus plantarum C70 isolated from camel milk. Int. J. Biol. Macromol. 2020, 144, 938–946. [Google Scholar] [CrossRef] [PubMed]
- Adesulu-Dahunsi, A.T.; Jeyaram, K.; Sanni, A.I.; Banwo, K. Production of exopolysaccharide by strains of Lactobacillus plantarum YP175 and OF101 isolated from traditional fermented cereal beverage. PeerJ 2018, 6, e5326. [Google Scholar] [CrossRef]
- Xu, Y.; Cui, Y.; Wang, X.; Yue, F.; Shan, Y.; Liu, B.; Zhou, Y.; Yi, Y.; Lü, X. Purification, characterization and bioactivity of exopolysaccharides produced by Lactobacillus plantarum KX041. Int. J. Biol. Macromol. 2019, 128, 480–492. [Google Scholar] [CrossRef]
- Ismail, B.; Nampoothiri, K. Exposition of antitumour activity of a chemically characterized exopolysaccharide from a probiotic Lactobacillus plantarum MTCC 9510. Biologia 2013, 68, 1041–1047. [Google Scholar] [CrossRef]
- Patel, A.; Lindström, C.; Patel, A.; Prajapati, J.; Holst, O. Probiotic properties of exopolysaccharide producing lactic acid bacteria isolated from vegetables and traditional Indian fermented foods. Int. J. Fermented Foods 2012, 1, 87–101. [Google Scholar]
- Zacharof, M.; Lovitt, R. Bacteriocins produced by lactic acid bacteria a review article. APCBEE Proc. 2012, 2, 50–56. [Google Scholar] [CrossRef]
- Settanni, L.; Corsetti, A. Application of bacteriocins in vegetable food biopreservation. Int. J. Food Microbiol. 2008, 121, 123–138. [Google Scholar] [CrossRef]
- Meade, E.; Slattery, M.A.; Garvey, M. Bacteriocins, potent antimicrobial peptides and the fight against multi drug resistant species: Resistance is futile? Antibiotics (Basel) 2020, 9, 32. [Google Scholar] [CrossRef]
- Todorov, S.D. Bacteriocins from Lactobacillus plantarum—Production, genetic organization and mode of action: Produção, organização genética e modo de ação. Braz. J. Microbiol. 2009, 40, 209–221. [Google Scholar] [CrossRef]
- Deegan, L.; Cotter, P.; Hill, C.; Ross, P. Bacteriocins: Biological tools for bio-preservation and shelf-life extension. Int. Dairy J. 2006, 16, 1058–1071. [Google Scholar] [CrossRef]
- Da Silva Sabo, S.; Vitolo, M.; González, J.M.D.; Oliveira, R.P.S. Overview of Lactobacillus plantarum as a promising bacteriocin producer among lactic acid bacteria. Food Res. Int. 2014, 64, 527–536. [Google Scholar] [CrossRef]
- Cotter, P.D.; Hill, C.; Ross, R.P. Bacteriocins: Developing innate immunity for food. Nat. Rev. Microbiol. 2005, 3, 777–788. [Google Scholar] [CrossRef] [PubMed]
- Florez, A.B.; Mayo, B. Genome analysis of Lactobacillus plantarum LL441 and genetic characterization of the locus for the lantibiotic plantaricin C. Front. Microbiol. 2018, 9, 1916. [Google Scholar] [CrossRef] [PubMed]
- Holo, H.; Jeknic, Z.; Daeschel, M.; Stevanovic, S.; Nes, I.F. Plantaricin W from Lactobacillus plantarum belongs to a new family of two-peptide lantibiotics. Microbiology 2001, 147, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Atrih, A.; Rekhif, N.; Milliere, J.; Lefebvre, G. Detection and characterisation of a bacteriocin produced by Lactobacillus plantarum C19. J. Microbiol. 1993, 39, 1173–1179. [Google Scholar]
- Van Reenen, C.A.; Dicks, L.M.; Chikindas, M.L. Isolation, purification and partial characterization of plantaricin 423, a bacteriocin produced by Lactobacillus plantarum. J. Appl. Microbiol. 1998, 84, 1131–1137. [Google Scholar] [CrossRef]
- Todorov, S.D.; Perin, L.M.; Carneiro, B.M.; Rahal, P.; Holzapfel, W.; Nero, L.A. Safety of Lactobacillus plantarum ST8Sh and its bacteriocin. Probiotics Antimicrob. Proteins 2017, 9, 334–344. [Google Scholar] [CrossRef]
- Diep, D.B.; Myhre, R.; Johnsborg, O.; Aakra, A.; Nes, I.F. Inducible bacteriocin production in Lactobacillus is regulated by differential expression of the pln operons and by two antagonizing response regulators, the activity of which is enhanced upon phosphorylation. Mol. Microbiol. 2003, 47, 483–494. [Google Scholar] [CrossRef]
- Enan, G.; Essaway, A.; Uyttendaele, M.; Debeverea, J. Antibacterial activity of Lactobacillus plantarum UG1 isolated from dry sausages: Characterisation, production, and bactericidal action of plantaricin UG1. Int. J. Food Microbiol. 1996, 30, 189–215. [Google Scholar] [CrossRef]
- Kanatani, K.; Oshimura, M. Plasmid-associated bacteriocin production by a Lactobacillus plantarum strain. Biosci. Biotechnol. Biochem. 1994, 58, 2084–2086. [Google Scholar] [CrossRef]
- Rekhif, N.; Atrih, A.; Lefebvre, G. Activity of plantaricin SA6, a bacteriocin produced by Lactobacillus plantarum SA6 isolated from fermented sausage. J. Appl. Bacteriol. 1995, 78, 349–358. [Google Scholar] [CrossRef]
- Ehrmann, M.; Remiger, A.; Eijsink, V.; Vogel, R. A gene cluster encoding plantaricin 1.25 beta and other bacteriocin-like peptides in Lactobacillus plantarum TMW1.25. Biochem. Bioph. Acta Gene Struct. Expr. 2000, 1490, 355–361. [Google Scholar] [CrossRef]
- Fricourt, B.; Barefoot, S.; Testin, R.; Hayasaka, S. Detection and activity of plantaricin F an antibacterial substance from Lactobacillus plantarum BF001 isolated from processed channel catfish. J. Food Protect. 1994, 57, 698–702. [Google Scholar] [CrossRef]
- Gonzalez, B.; Arca, P.; Mayo, B.; Suárez, J. Detection, purification and partial characterisation of plantaricin C, a bacteriocin produced by a Lactobacillus plantarum strain of dairy origin. Appl. Environ. Microbiol. 1994, 60, 2158–2163. [Google Scholar] [CrossRef]
- Turner, D.L.; Brennan, L.; Meyer, H.E.; Lohaus, C.; Siethoff, C.; Costa, H.S.; Gonzalez, B.; Santos, H.; Suárez, J.E. Solution structure of plantaricin C, a novel lantibiotic. Eur. J. Biochem. 1999, 264, 833–839. [Google Scholar] [CrossRef] [PubMed]
- Rekhif, N.; Atrih, A.; Lefebvre, G. Characterisation and partial purification of plantaricin LC74, a bacteriocin produced by Lactobacillus plantarum LC74. Biotechnol. Lett. 1994, 16, 771–776. [Google Scholar] [CrossRef]
- Wen, L.S.; Philip, K.; Ajam, N. Purification, characterization and mode of action of plantaricin K25 produced by Lactobacillus plantarum. Food Control 2016, 60, 430–439. [Google Scholar] [CrossRef]
- Todorov, S.; Onno, B.; Sorokine, O.; Chobert, J.; Ivanova, I.; Dousset, X. Detection and characterisation of a novel antibacterial substance produced by Lactobacillus plantarum ST31 isolated from sourdough. Int. J. Food Microbiol. 1999, 48, 167–177. [Google Scholar] [CrossRef]
- Chen, Y.S.; Wang, Y.C.; Chow, Y.S.; Yanagida, F.; Liao, C.C.; Chiu, C.M. Purification and characterization of plantaricin Y, a novel bacteriocin produced by Lactobacillus plantarum 510. Arch. Microbiol. 2014, 196, 193–199. [Google Scholar] [CrossRef]
- Jiménez-Díaz, R.; Ruiz-Barba, J.L.; Cathcart, D.P.; Holo, H.; Nes, I.F.; Sletten, K.H.; Warner, P.J. Purification and partial amino acid sequence of plantaricin S, a bacteriocin produced by Lactobacillus plantarum LPCO10, the activity of which depends on the complementary action of two peptides. Appl. Environ. Microbiol. 1995, 61, 4459–4463. [Google Scholar] [CrossRef]
- Hu, M.; Zhao, H.; Zhang, C.; Yu, J.; Lu, Z. Purification and characterization of plantaricin 163, a novel bacteriocin produced by Lactobacillus plantarum 163 isolated from traditional Chinese fermented vegetables. J. Agric. Food Chem. 2013, 61, 11676–11682. [Google Scholar] [CrossRef]
- Zhao, S.; Han, J.; Bie, X.; Lu, Z.; Zhang, C.; Lv, F. Purification and characterization of Plantaricin JLA-9: A Novel bacteriocin against Bacillus spp. produced by Lactobacillus plantarum JLA-9 from Suan-Tsai, a traditional Chinese fermented cabbage. J. Agric. Food Chem. 2016, 64, 2754–2764. [Google Scholar] [CrossRef] [PubMed]
- Andersse, E.; Diep, D.; Nes, I.; Eijsink, V.; Nissen-Meyer, J. Antagonistic activity of Lactobacillus plantarum C11: Two new two-peptide bacteriocins, plantaricins EF and JK, and the induction factor plantaricin A. Appl. Environ. Microbiol. 1998, 64, 2269–2272. [Google Scholar] [CrossRef] [PubMed]
- Daeschel, M.; McKeneney, M.; McDonald, L. Bacteriocidal activity of Lactobacillus plantarum C-11. Food Microbiol. 1990, 7, 91–98. [Google Scholar] [CrossRef]
- Zhu, X.; Zhao, Y.; Sun, Y.; Gu, Q. Purification and characterisation of plantaricin ZJ008, a novel bacteriocin against Staphylococcus spp. from Lactobacillus plantarum ZJ008. Food Chem. 2014, 165, 216–223. [Google Scholar] [CrossRef]
- Song, D.F.; Zhu, M.Y.; Gu, Q. Purification and characterization of Plantaricin ZJ5, a new bacteriocin produced by Lactobacillus plantarum ZJ5. PLoS ONE 2014, 9, e105549. [Google Scholar] [CrossRef]
- Kato, T.; Matsuda, T.; Ogawa, E.; Ogawa, H.; Kato, H.; Doi, U.; Nakamura, R. Plantaricin-149, a bacteriocin produced by Lactobacillus plantarum NRIC 149. J. Ferment. Bioeng. 1994, 77, 277–282. [Google Scholar] [CrossRef]
- Franz, C.M.; Du Toit, M.; Olasupo, N.A.; Schillinger, U.; Holzapfel, W.H. Plantaricin D, a bacteriocin produced by Lactobacillus plantarum BFE 905 ready-to-eat salad. Lett. Appl. Microbiol. 1998, 26, 231–235. [Google Scholar] [CrossRef]
- Mourad, K.; Halima, Z.K. Detection and activity of plantaricin OL15 a bacteriocin produced by Lactobacillus plantarum OL15 isolated from algerian fermented olives. Grasas Aceites 2005, 56, 192–197. [Google Scholar] [CrossRef]
- Lv, X.; Lin, Y.; Jie, Y.; Sun, M.; Zhang, B.; Bai, F.; Zhao, H.; Li, J. Purification, characterization, and action mechanism of plantaricin DL3, a novel bacteriocin against Pseudomonas aeruginosa produced by Lactobacillus plantarum DL3 from Chinese Suan-Tsai. Eur. Food Res. Technol. 2018, 244, 323–331. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, L.; Yi, H.; Han, X.; Chi, C. Identification and characterization of plantaricin Q7, a novel plantaricin produced by Lactobacillus plantarum Q7. LWT-Food Sci. Technol. 2016, 71, 386–390. [Google Scholar] [CrossRef]
- Gong, H.S.; Meng, X.C.; Wang, H. Plantaricin MG active against Gram-negative bacteria produced by Lactobacillus plantarum KLDS1.0391 isolated from “Jiaoke”, a traditional fermented cream from China. Food Control 2010, 21, 89–96. [Google Scholar] [CrossRef]
- Wang, Y.; Shang, N.; Qin, Y.; Zhang, Y.; Zhang, J.; Li, P. The complete genome sequence of Lactobacillus plantarum LPL-1, a novel antibacterial probiotic producing class IIa bacteriocin. J. Biotechnol. 2018, 266, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qin, Y.; Xie, Q.; Zhang, Y.; Hu, J.; Li, P. Purification and characterization of plantaricin LPL-1, a novel class IIa bacteriocin produced by Lactobacillus plantarum LPL-1 isolated from fermented fish. Front. Microbiol. 2018, 9, 2276. [Google Scholar] [CrossRef]
- Wang, Y.; Qin, Y.; Zhang, Y.; Wu, R.; Li, P. Antibacterial mechanism of plantaricin LPL-1, a novel class IIa bacteriocin against Listeria monocytogenes. Food Control 2019, 97, 87–93. [Google Scholar] [CrossRef]
- Du, H.; Yang, J.; Lu, X.; Lu, Z.; Bie, X.; Zhao, H.; Zhang, C.; Lu, F. Purification, characterization, and mode of action of plantaricin GZ1-27, a novel bacteriocin against Bacillus cereus. J. Agric. Food Chem. 2018, 66, 4716–4724. [Google Scholar] [CrossRef]
- Pei, J.; Li, X.; Han, H.; Tao, Y. Purification and characterization of plantaricin SLG1, a novel bacteriocin produced by Lb. plantarum isolated from yak cheese. Food Control 2018, 84, 111–117. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, L.; Hao, Y.; Zhong, S.; Liu, H.; Han, T.; Xie, Y. Isolation and partial characterization of a bacteriocin produced by Lactobacillus plantarum BM-1 isolated from a traditionally fermented Chinese meat product. Microbiol. Immunol. 2013, 57, 746–755. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.; Todorov, S.; Ivanova, I.; Belguesmia, Y.; Choiset, Y.; Rabesona, H.; Chobert, J.-M.; Haertlé, T.; Franco, B. Characterization of a two-peptide plantaricin produced by Lactobacillus plantarum MBSa4 isolated from Brazilian salami. Food Control 2016, 60, 103–112. [Google Scholar] [CrossRef]
- Dinev, T.; Beev, G.; Tzanova, M.; Denev, S.; Dermendzhieva, D.; Stoyanova, A. Antimicrobial activity of Lactobacillus plantarum against pathogenic and food spoilage microorganisms: A review. Bulg. J. Vet. Med. 2018, 21, 253–268. [Google Scholar] [CrossRef]
- Al-Tawaha, R.; Meng, C. Potential benefits of Lactobacillus plantarum as probiotic and its advantages in human health and industrial applications: A review. Adv. Environ. Biol. 2018, 12, 16–27. [Google Scholar]
- Yu, H.J.; Chen, Y.F.; Yang, H.J.; Yang, J.; Xue, J.G.; Li, C.K.; Kwok, L.Y.; Zhang, H.P.; Sun, T.S. Screening for Lactobacillus plantarum with potential inhibitory activity against enteric pathogens. Ann. Microbiol. 2015, 65, 1257–1265. [Google Scholar] [CrossRef]
- Arena, M.P.; Silvain, A.; Normanno, G.; Grieco, F.; Drider, D.; Spano, G.; Fiocco, D. Use of Lactobacillus plantarum strains as a bio-control strategy against food-borne pathogenic microorganisms. Front. Microbiol. 2016, 7, 464. [Google Scholar] [CrossRef]
- Tremonte, P.; Pannella, G.; Succi, M.; Tipaldi, L.; Sturchio, M.; Coppola, R.; Luongo, D.; Sorrentino, E. Antimicrobial activity of Lactobacillus plantarum strains isolated from different environments: A preliminary study. Int. Food Res. J. 2017, 24, 852–859. [Google Scholar]
- Lau, L.Y.J.; Chye, F.Y. Antagonistic effects of Lactobacillus plantarum 0612 on the adhesion of selected foodborne enteropathogens in various colonic environments. Food Control 2018, 91, 237–247. [Google Scholar]
- Bonatsou, S.; Tassou, C.C.; Panagou, E.Z.; Nychas, G.E. Table olive fermentation using starter cultures with multifunctional potential. Microorganisms 2017, 5, 30. [Google Scholar] [CrossRef]
- Russo, P.; Arena, M.P.; Fiocco, D.; Capozzi, V.; Drider, D.; Spano, G. Lactobacillus plantarum with broad antifungal activity: A promising approach to increase safety and shelf-life of cereal-based products. Int. J. Food Microbiol. 2017, 247, 48–54. [Google Scholar] [CrossRef]
- Dalié, D.; Deschamps, A.; Richard-Forget, F. Lactic acid bacteria–potential for control of mould growth and mycotoxins: A review. Food Control 2010, 21, 370–380. [Google Scholar] [CrossRef]
- Ryu, E.H.; Yang, E.J.; Woo, E.R.; Chang, H.C. Purification and characterization of antifungal compounds from Lactobacillus plantarum HD1 isolated from kimchi. Food Microbiol. 2014, 41, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Crowley, S.; Mahony, J.; van Sinderen, D. Current perspectives on antifungal lactic acid bacteria as natural bio-preservatives. Trends Food Sci. Technol. 2013, 33, 93–109. [Google Scholar] [CrossRef]
- Galié, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898. [Google Scholar] [CrossRef]
- Kumar, M.; Dhaka, P.; Vijay, D.; Vergis, J.; Mohan, V.; Kumar, A.; Kurkure, N.V.; Barbuddhe, S.B.; Malik, S.V.; Rawool, D.B. Antimicrobial effects of Lactobacillus plantarum and Lactobacillus acidophilus against multidrug-resistant enteroaggregative Escherichia coli. Int. J. Antimicrob. Agents 2016, 48, 265–270. [Google Scholar] [CrossRef]
- McNicholl, A.G.; Molina-Infante, J.; Lucendo, A.J.; Calleja, J.L.; Pérez-Aisa, Á.; Modolell, I.; Aldeguer, X.; Calafat, M.; Comino, L.; Ramas, M.; et al. Probiotic supplementation with Lactobacillus plantarum and Pediococcus acidilactici for Helicobacter pylori therapy: A randomized, double-blind, placebo-controlled trial. Helicobacter 2018, 23, e12529. [Google Scholar] [CrossRef]
- Onbas, T.; Osmanagaoglu, O.; Kiran, F. Potential properties of Lactobacillus plantarum F-10 as a bio-control strategy for wound infections. Probiotics Antimicrob. Proteins 2019, 11, 1110–1123. [Google Scholar] [CrossRef] [PubMed]
- Falagas, M.E.; Betsi, G.I.; Athanasiou, S. Probiotics for prevention of recurrent vulvovaginal candidiasis: A review. J. Antimicrob. Chemother. 2006, 58, 266–272. [Google Scholar] [CrossRef]
- Kang, C.-H.; Kim, Y.; Han, S.H.; Kim, J.-S.; Paek, N.-S.; So, J.-S. In vitro probiotic properties of vaginal Lactobacillus fermentum MG901 and Lactobacillus plantarum MG989 against Candida albicans. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 228, 232–237. [Google Scholar] [CrossRef]
- Beck, B.R.; Park, G.-S.; Lee, Y.H.; Im, S.; Jeong, D.Y.; Kang, J. Whole genome analysis of Lactobacillus plantarum strains isolated from kimchi and determination of probiotic properties to treat mucosal infections by Candida albicans and Gardnerella vaginalis. Front. Microbiol. 2019, 10, 433. [Google Scholar] [CrossRef]
- Cenci, G.; Rossi, J.; Trotta, F.; Caldini, G. Lactic acid bacteria isolated from dairy products inhibit genotoxic effect of 4-nitroquinoline-1-oxide in SOS-chromotest. Syst. Appl. Microbiol. 2002, 25, 483–490. [Google Scholar] [CrossRef]
- Felton, J.S.; Knize, M.G.; Wu, R.W.; Colvin, M.E.; Hatch, F.T.; Malfatti, M.A. Mutagenic potency of food-derived heterocyclic amines. Mutat. Res. 2007, 616, 90–94. [Google Scholar] [CrossRef]
- Raman, M.; Ambalam, P.; Kondepudi, K.K.; Pithva, S.; Kothari, C.; Patel, A.T.; Purama, R.K.; Dave, J.M.; Vyas, B.R. Potential of probiotics, prebiotics and synbiotics for management of colorectal cancer. Gut Microbes 2013, 4, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Le Magueresse-Battistoni, B.; Labaronne, E.; Vidal, H.; Naville, D. Endocrine disrupting chemicals in mixture and obesity, diabetes and related metabolic disorders. World J. Biol. Chem. 2017, 8, 108–119. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, N.; Prete, R.; Perugini, M.; Merola, C.; Battista, N.; Corsetti, A. Probiotic antigenotoxic activity as a DNA bioprotective tool: A minireview with focus on endocrine disruptors. FEMS Microbiol. Lett. 2020, 367. [Google Scholar] [CrossRef]
- Caldini, G.; Trotta, F.; Villarini, M.; Moretti, M.; Pasquini, R.; Scassellati-Sforzolini, G.; Cenci, G. Screening of potential lactobacilli antigenotoxicity by microbial and mammalian cell-based tests. Int. J. Food Microbiol. 2005, 102, 37–47. [Google Scholar] [CrossRef]
- Hsieh, M.L.; Chou, C.C. Mutagenicity and antimutagenic effect of soymilk fermented with lactic acid bacteria and bifidobacteria. Int. J. Food Microbiol. 2006, 111, 43–47. [Google Scholar] [CrossRef]
- Davis, C.D.; Milner, J.A. Gastrointestinal microflora, food components and colon cancer prevention. J. Nutr. Biochem. 2009, 20, 743–752. [Google Scholar] [CrossRef]
- Federici, E.; Prete, R.; Lazzi, C.; Pellegrini, N.; Moretti, M.; Corsetti, A.; Cenci, G. Bacterial composition, genotoxicity, and cytotoxicity of fecal samples from individuals consuming omnivorous or vegetarian diets. Front. Microbiol. 2017, 8, 300. [Google Scholar] [CrossRef] [PubMed]
- Walia, S.; Keshani; Sood, S.; Kanwar, S.S. Exhibiotion of DNA-bioprotective activity by microflora of traditional fermented foods of North-Western Himalayas. Food Res. Int. 2014, 55, 176–180. [Google Scholar] [CrossRef]
- Prete, R.; Tofalo, R.; Federici, E.; Ciarrocchi, A.; Cenci, G.; Corsetti, A. Food-associated Lactobacillus plantarum and inhibit the genotoxic effect of 4-Nitroquinoline-1-oxide. Front. Microbiol. 2017, 8, 2349. [Google Scholar] [CrossRef] [PubMed]
- Kurhan, Ş.; Çakir, I. DNA-bioprotective effects of lactic acid bacteria against aflatoxin B1. Curr. Res. Nutr. Food Sci. J. 2016, 4, 87–91. [Google Scholar] [CrossRef]
- Bartkiene, E.; Zavistanaviciute, P.; Lele, V.; Ruzauskas, M.; Bartkevics, V.; Bernatoniene, J.; Gallo, P.; Tenore, G.C.; Santini, A. Lactobacillus plantarum LUHS135 and paracasei LUHS244 as functional starter cultures for the food fermentation industry: Characterisation, mycotoxin-reducing properties, optimisation of biomass growth and sustainable encapsulation by using dairy by-products. LWT 2018, 93, 649–658. [Google Scholar] [CrossRef]
- Zhai, Q.; Yu, L.; Li, T.; Zhu, J.; Zhang, C.; Zhao, J.; Zhang, H.; Chen, W. Effect of dietary probiotic supplementation on intestinal microbiota and physiological conditions of Nile tilapia (Oreochromis niloticus) under waterborne cadmium exposure. Antonie Van Leeuwenhoek 2017, 110, 501–513. [Google Scholar] [CrossRef] [PubMed]
- Zang, L.; Ma, Y.; Huang, W.; Ling, Y.; Sun, L.; Wang, X.; Zeng, A.; Dahlgren, R.A.; Wang, C.; Wang, H. Dietary Lactobacillus plantarum ST-III alleviates the toxic effects of triclosan on zebrafish (Danio rerio) via gut microbiota modulation. Fish Shellfish Immunol. 2019, 84, 1157–1169. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.-T.; Yang, C.-X.; Luo, B.-B.; Zhou, K.; Liu, S.-L. Efficiency of dairy strains of lactic acid bacteria to bind bisphenol A in phosphate buffer saline. Food Control 2017, 73, 1203–1209. [Google Scholar] [CrossRef]
- Trinder, M.; Bisanz, J.E.; Burton, J.P.; Reid, G. Probiotic lactobacilli: A potential prophylactic treatment for reducing pesticide absorption in humans and wildlife. Benef. Microbes 2015, 6, 841–847. [Google Scholar] [CrossRef]
- Garcia-Gonzalez, N.; Prete, R.; Battista, N.; Corsetti, A. Adhesion properties of food-associated Lactobacillus plantarum strains on human intestinal epithelial cells and modulation of IL-8 release. Front. Microbiol. 2018, 9, 2392. [Google Scholar] [CrossRef]
- Prete, R.; Garcia-Gonzalez, N.; Di Mattia, C.D.; Corsetti, A.; Battista, N. Food-borne Lactiplantibacillus plantarum protect normal intestinal cells against inflammation by modulating reactive oxygen species and IL-23/IL-17 axis. Sci. Rep. 2020, 10, 16340. [Google Scholar] [CrossRef]
- Chiang, J.Y. Bile acid metabolism and signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar] [CrossRef] [PubMed]
- Urdaneta, V.; Casadesús, J. Interactions between bacteria and bile salts in the gastrointestinal and hepatobiliary tracts. Front. Med. 2017, 4, 163. [Google Scholar] [CrossRef]
- Ren, J.; Sun, K.; Wu, Z.; Yao, J.; Guo, B. All 4 bile salt hydrolase proteins are responsible for the hydrolysis activity in Lactobacillus plantarum ST-III. J. Food Sci. 2011, 76, M622–M628. [Google Scholar] [CrossRef] [PubMed]
- Begley, M.; Hill, C.; Gahan, C.G. Bile salt hydrolase activity in probiotics. Appl. Environ. Microbiol. 2006, 72, 1729–1738. [Google Scholar] [CrossRef] [PubMed]
- Long, S.L.; Gahan, C.G.M.; Joyce, S.A. Interactions between gut bacteria and bile in health and disease. Mol. Asp. Med. 2017, 56, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.B.; Yi, S.H.; Lee, B.H. Purification and characterization of three different types of bile salt hydrolases from Bifidobacterium strains. J. Dairy Sci. 2004, 87, 258–266. [Google Scholar] [CrossRef]
- Kishinaka, M.; Umeda, A.; Kuroki, S. High concentrations of conjugated bile acids inhibit bacterial growth of Clostridium perfringens and induce its extracellular cholylglycine hydrolase. Steroids 1994, 59, 485–489. [Google Scholar] [CrossRef]
- Wijaya, A.; Hermann, A.; Abriouel, H.; Specht, I.; Yousif, N.M.; Holzapfel, W.H.; Franz, C.M. Cloning of the bile salt hydrolase (bsh) gene from Enterococcus faecium FAIR-E 345 and chromosomal location of bsh genes in food enterococci. J. Food Prot. 2004, 67, 2772–2778. [Google Scholar] [CrossRef]
- Begley, M.; Sleator, R.D.; Gahan, C.G.; Hill, C. Contribution of three bile-associated loci, bsh, pva, and btlB, to gastrointestinal persistence and bile tolerance of Listeria monocytogenes. Infect. Immun. 2005, 73, 894–904. [Google Scholar] [CrossRef]
- Chae, J.P.; Valeriano, V.D.; Kim, G.B.; Kang, D.K. Molecular cloning, characterization and comparison of bile salt hydrolases from Lactobacillus johnsonii PF01. J. Appl. Microbiol. 2013, 114, 121–133. [Google Scholar] [CrossRef]
- Jayashree, S.; Pooja, S.; Pushpanathan, M.; Rajendhran, J.; Gunasekaran, P. Identification and characterization of bile salt hydrolase genes from the genome of Lactobacillus fermentum MTCC 8711. Appl. Biochem. Biotechnol. 2014, 174, 855–866. [Google Scholar] [CrossRef]
- Christiaens, H.; Leer, R.J.; Pouwels, P.H.; Verstraete, W. Cloning and expression of a conjugated bile acid hydrolase gene from Lactobacillus plantarum by using a direct plate assay. Appl. Environ. Microbiol. 1992, 58, 3792–3798. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.M.; Bongers, R.S.; de Vos, W.M.; Kleerebezem, M. Functional analysis of four bile salt hydrolase and penicillin acylase family members in Lactobacillus plantarum WCFS1. Appl. Environ. Microbiol. 2008, 74, 4719–4726. [Google Scholar] [CrossRef]
- Prete, R.; Long, S.L.; Gallardo, A.L.; Gahan, C.G.; Corsetti, A.; Joyce, S.A. Beneficial bile acid metabolism from Lactobacillus plantarum of food origin. Sci. Rep. 2020, 10, 1165. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.; Gioiello, A.; Noriega, L.; Strehle, A.; Oury, J.; Rizzo, G.; Macchiarulo, A.; Yamamoto, H.; Mataki, C.; Pruzanski, M.; et al. TGR5-mediated bile acid sensing controls glucose homeostasis. Cell Metab. 2009, 10, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Joyce, S.A.; MacSharry, J.; Casey, P.G.; Kinsella, M.; Murphy, E.F.; Shanahan, F.; Hill, C.; Gahan, C.G. Regulation of host weight gain and lipid metabolism by bacterial bile acid modification in the gut. Proc. Natl. Acad. Sci. USA 2014, 111, 7421–7426. [Google Scholar] [CrossRef]
- Wan, Y.Y.; Sheng, L. Regulation of bile acid receptor activity (☆). Liver Res. 2018, 2, 180–185. [Google Scholar] [CrossRef]
- Joyce, S.A.; Shanahan, F.; Hill, C.; Gahan, C.G. Bacterial bile salt hydrolase in host metabolism: Potential for influencing gastrointestinal microbe-host crosstalk. Gut Microbes 2014, 5, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.C.; Luo, X.G.; Wang, C.X.; Ma, D.Y.; Wang, Y.; He, Y.Y.; Li, W.; Zhou, H.; Zhang, T.C. Cloning and analysis of bile salt hydrolase genes from Lactobacillus plantarum CGMCC No. 8198. Biotechnol. Lett. 2014, 36, 975–983. [Google Scholar] [CrossRef]
- Jones, M.L.; Tomaro-Duchesneau, C.; Martoni, C.J.; Prakash, S. Cholesterol lowering with bile salt hydrolase-active probiotic bacteria, mechanism of action, clinical evidence, and future direction for heart health applications. Expert Opin. Biol. Ther. 2013, 13, 631–642. [Google Scholar] [CrossRef]
- Pols, T.W.H.; Puchner, T.; Korkmaz, H.I.; Vos, M.; Soeters, M.R.; de Vries, C.J.M. Lithocholic acid controls adaptive immune responses by inhibition of Th1 activation through the Vitamin D receptor. PLoS ONE 2017, 12, e0176715. [Google Scholar] [CrossRef]
- Govindarajan, K.; MacSharry, J.; Casey, P.G.; Shanahan, F.; Joyce, S.A.; Gahan, C.G. Unconjugated bile acids influence expression of circadian genes: A potential mechanism for microbe-host crosstalk. PLoS ONE 2016, 11, e0167319. [Google Scholar] [CrossRef]
- Amaretti, A.; di Nunzio, M.; Pompei, A.; Raimondi, S.; Rossi, M.; Bordoni, A. Antioxidant properties of potentially probiotic bacteria: In vitro and in vivo activities. Appl. Microbiol. Biotechnol. 2013, 97, 809–817. [Google Scholar] [CrossRef]
- Li, S.; Zhao, Y.; Zhang, L.; Zhang, X.; Huang, L.; Li, D.; Niu, C.; Yang, Z.; Wang, Q. Antioxidant activity of Lactobacillus plantarum strains isolated from traditional Chinese fermented foods. Food Chem. 2012, 135, 1914–1919. [Google Scholar] [CrossRef] [PubMed]
- Mu, G.; Gao, Y.; Tuo, Y.; Li, H.; Zhang, Y.; Qian, F.; Jiang, S. Assessing and comparing antioxidant activities of lactobacilli strains by using different chemical and cellular antioxidant methods. J. Dairy Sci. 2018, 101, 10792–10806. [Google Scholar] [CrossRef] [PubMed]
- Xing, J.Y.; Song, G.P.; Deng, J.P.; Jiang, L.Z.; Xiong, P.; Yang, B.J.; Liu, S.S. Antitumor effects and mechanism of novel emodin rhamnoside derivatives against human cancer cells in vitro. PLoS ONE 2015, 10, e0144781. [Google Scholar] [CrossRef] [PubMed]
- Devi, S.M.; Aishwarya, S.; Halami, P.M. Discrimination and divergence among Lactobacillus plantarum-group (LPG) isolates with reference to their probiotic functionalities from vegetable origin. Syst. Appl. Microbiol. 2016, 39, 562–570. [Google Scholar] [CrossRef] [PubMed]
- Valan Arasu, M.; Jung, M.W.; Ilavenil, S.; Jane, M.; Kim, D.H.; Lee, K.D.; Park, H.S.; Hur, T.Y.; Choi, G.J.; Lim, Y.C.; et al. Isolation and characterization of antifungal compound from Lactobacillus plantarum KCC-10 from forage silage with potential beneficial properties. J. Appl. Microbiol. 2013, 115, 1172–1185. [Google Scholar] [CrossRef] [PubMed]
- Das, D.; Goyal, A. Antioxidant activity and γ-aminobutyric acid (GABA) producing ability of probiotic Lactobacillus plantarum DM5 isolated from Marcha of Sikkim. LWT-Food Sci. Technol. 2015, 61, 263–268. [Google Scholar] [CrossRef]
- Tang, W.; Xing, Z.; Li, C.; Wang, J.; Wang, Y. Molecular mechanisms and in vitro antioxidant effects of Lactobacillus plantarum MA2. Food Chem. 2017, 221, 1642–1649. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, M.; Yamamoto, Y.; Kamio, Y. Molecular biology of oxygen tolerance in lactic acid bacteria: Functions of NADH oxidases and Dpr in oxidative stress. J. Biosci. Bioeng. 2000, 90, 484–493. [Google Scholar] [CrossRef]
- Wang, X.; Shao, C.; Liu, L.; Guo, X.; Xu, Y.; Lü, X. Optimization, partial characterization and antioxidant activity of an exopolysaccharide from Lactobacillus plantarum KX041. Int. J. Biol. Macromol. 2017, 103, 1173–1184. [Google Scholar] [CrossRef]
- Nanjundaiah, Y.S.; Wright, D.A.; Baydoun, A.R.; O’Hare, W.T.; Ali, Z.; Khaled, Z.; Sarker, M.H. Lactobacillus rhamnosus GG conditioned media modulates acute reactive oxygen species and nitric oxide in J774 murine macrophages. Biochem. Biophys. Rep. 2016, 6, 68–75. [Google Scholar]
- Swanson, P.A., II; Kumar, A.; Samarin, S.; Vijay-Kumar, M.; Kundu, K.; Murthy, N.; Hansen, J.; Nusrat, A.; Neish, A.S. Enteric commensal bacteria potentiate epithelial restitution via reactive oxygen species-mediated inactivation of focal adhesion kinase phosphatases. Proc. Natl. Acad. Sci. USA 2011, 108, 8803–8808. [Google Scholar] [CrossRef]
- Matthews, J.D.; Reedy, A.R.; Wu, H.; Hinrichs, B.H.; Darby, T.M.; Addis, C.; Robinson, B.S.; Go, Y.-M.; Jones, D.P.; Jones, R.M. Proteomic analysis of microbial induced redox-dependent intestinal signaling. Redox Biol. 2019, 20, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of action of probiotics. Adv. Nutr. 2019, 10, S49–S66. [Google Scholar] [CrossRef] [PubMed]
- Monteagudo-Mera, A.; Rastall, R.A.; Gibson, G.R.; Charalampopoulos, D.; Chatzifragkou, A. Adhesion mechanisms mediated by probiotics and prebiotics and their potential impact on human health. Appl. Microbiol. Biotechnol. 2019, 103, 6463–6472. [Google Scholar] [CrossRef]
- Maldonado Galdeano, C.; Cazorla, S.I.; Lemme Dumit, J.M.; Vélez, E.; Perdigón, G. Beneficial effects of probiotic consumption on the immune system. Ann. Nutr. Metab. 2019, 74, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Tan, R.S.; Ho, B.; Leung, B.P.; Ding, J.L. TLR cross-talk confers specificity to innate immunity. Int. Rev. Immunol. 2014, 33, 443–453. [Google Scholar] [CrossRef]
- Hug, H.; Mohajeri, M.H.; La Fata, G. Toll-Like receptors: Regulators of the immune response in the human gut. Nutrients 2018, 10, 203. [Google Scholar] [CrossRef]
- Caruso, R.; Warner, N.; Inohara, N.; Núñez, G. NOD1 and NOD2: Signaling, host defense, and inflammatory disease. Immunity 2014, 41, 898–908. [Google Scholar] [CrossRef]
- Takeda, S.; Kawahara, S.; Hidaka, M.; Yoshida, H.; Watanabe, W.; Takeshita, M.; Kikuchi, Y.; Bumbein, D.; Muguruma, M.; Kurokawa, M. Effects of oral administration of probiotics from Mongolian dairy products on the Th1 immune response in mice. Biosci. Biotechnol. Biochem. 2013, 77, 1372–1378. [Google Scholar] [CrossRef]
- Kwon, E.K.; Kang, G.-D.; Kim, W.-K.; Han, M.J.; Kim, D.-H. Lactobacillus plantarum LC27 and Bifidobacterium longum LC67 simultaneously alleviate ethanol-induced gastritis and hepatic injury in mice. J. Funct. Foods 2017, 38, 389–398. [Google Scholar] [CrossRef]
- Vitali, B.; Minervini, G.; Rizzello, C.G.; Spisni, E.; Maccaferri, S.; Brigidi, P.; Gobbetti, M.; Di Cagno, R. Novel probiotic candidates for humans isolated from raw fruits and vegetables. Food Microbiol. 2012, 31, 116–125. [Google Scholar] [CrossRef]
- Carey, C.M.; Kostrzynska, M. Lactic acid bacteria and bifidobacteria attenuate the proinflammatory response in intestinal epithelial cells induced by Salmonella enterica serovar Typhimurium. Can. J. Microbiol. 2013, 59, 9–17. [Google Scholar] [CrossRef]
- Murosaki, S.; Yamamoto, Y.; Ito, K.; Inokuchi, T.; Kusaka, H.; Yoshikai, Y. Heat-killed Lactobacillus plantarum L-137 suppresses naturally fed antigen-specific IgE production by stimulation of IL-12 production in mice. J. Allergy Clin. Immunol. 1998, 102, 57–64. [Google Scholar] [CrossRef]
- Wang, J.; Wu, T.; Fang, X.; Min, W.; Yang, Z. Characterization and immunomodulatory activity of an exopolysaccharide produced by Lactobacillus plantarum JLK0142 isolated from fermented dairy tofu. Int. J. Biol. Macromol. 2018, 115, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Zago, M.; Fornasari, M.E.; Carminati, D.; Burns, P.; Suàrez, V.; Vinderola, G.; Reinheimer, J.; Giraffa, G. Characterization and probiotic potential of Lactobacillus plantarum strains isolated from cheeses. Food Microbiol. 2011, 28, 1033–1040. [Google Scholar] [CrossRef]
- Kawashima, T.; Hayashi, K.; Kosaka, A.; Kawashima, M.; Igarashi, T.; Tsutsui, H.; Tsuji, N.M.; Nishimura, I.; Hayashi, T.; Obata, A. Lactobacillus plantarum strain YU from fermented foods activates Th1 and protective immune responses. Int. Immunopharmacol. 2011, 11, 2017–2024. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.E.; Han, M.J.; Kim, S.Y.; Kim, D.H. Lactobacillus plantarum CLP-0611 ameliorates colitis in mice by polarizing M1 to M2-like macrophages. Int. Immunopharmacol. 2014, 21, 186–192. [Google Scholar] [CrossRef]
- Ren, C.; Zhang, Q.; de Haan, B.J.; Zhang, H.; Faas, M.M.; de Vos, P. Identification of TLR2/TLR6 signalling lactic acid bacteria for supporting immune regulation. Sci. Rep. 2016, 6, 34561. [Google Scholar] [CrossRef] [PubMed]
- Murofushi, Y.; Villena, J.; Morie, K.; Kanmani, P.; Tohno, M.; Shimazu, T.; Aso, H.; Suda, Y.; Hashiguchi, K.; Saito, T.; et al. The toll-like receptor family protein RP105/MD1 complex is involved in the immunoregulatory effect of exopolysaccharides from Lactobacillus plantarum N14. Mol. Immunol. 2015, 64, 63–75. [Google Scholar] [CrossRef]
- Kujawa-Szewieczek, A.; Adamczak, M.; Kwiecień, K.; Dudzicz, S.; Gazda, M.; Więcek, A. The effect of Lactobacillus plantarum 299v on the incidence of Clostridium difficile infection in high risk patients treated with antibiotics. Nutrients 2015, 7, 10179–10188. [Google Scholar] [CrossRef]
- Fedorak, R.N.; Feagan, B.G.; Hotte, N.; Leddin, D.; Dieleman, L.A.; Petrunia, D.M.; Enns, R.; Bitton, A.; Chiba, N.; Paré, P.; et al. The probiotic VSL#3 has anti-inflammatory effects and could reduce endoscopic recurrence after surgery for Crohn’s disease. Clin. Gastroenterol. Hepatol. 2015, 13, 928–935.e2. [Google Scholar] [CrossRef]
- Barraza-Ortiz, D.A.; Pérez-López, N.; Medina-López, V.M.; Minero-Alfaro, J.I.; Zamarripa-Dorsey, F.; Fernández-Martínez, N.D.C.; Llorente-Ramón, A.; Ramos-Aguilar, G.A. Combination of a probiotic and an antispasmodic increases quality of life and reduces symptoms in patients with irritable bowel syndrome: A pilot study. Dig. Dis. 2020. [Google Scholar] [CrossRef]
- Hütt, P.; Songisepp, E.; Rätsep, M.; Mahlapuu, R.; Kilk, K.; Mikelsaar, M. Impact of probiotic Lactobacillus plantarum TENSIA in different dairy products on anthropometric and blood biochemical indices of healthy adults. Benef. Microbes 2015, 6, 233–243. [Google Scholar] [CrossRef]
- Alisi, A.; Bedogni, G.; Baviera, G.; Giorgio, V.; Porro, E.; Paris, C.; Giammaria, P.; Reali, L.; Anania, F.; Nobili, V. Randomised clinical trial: The beneficial effects of VSL#3 in obese children with non-alcoholic steatohepatitis. Aliment. Pharmacol. Ther. 2014, 39, 1276–1285. [Google Scholar] [CrossRef] [PubMed]
- Bosch, M.; Fuentes, M.C.; Audivert, S.; Bonachera, M.A.; Peiró, S.; Cuñé, J. Lactobacillus plantarum CECT 7527, 7528 and 7529: Probiotic candidates to reduce cholesterol levels. J. Sci. Food Agric. 2014, 94, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Miraghajani, M.; Dehsoukhteh, S.S.; Rafie, N.; Hamedani, S.G.; Sabihi, S.; Ghiasvand, R. Potential mechanisms linking probiotics to diabetes: A narrative review of the literature. Sao Paulo Med. J. 2017, 135, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Vicariotto, F.; Mogna, L.; Del Piano, M. Effectiveness of the two microorganisms Lactobacillus fermentum LF15 and Lactobacillus plantarum LP01, formulated in slow-release vaginal tablets, in women affected by bacterial vaginosis: A pilot study. J. Clin. Gastroenterol. 2014, 48 (Suppl. 1), 106–112. [Google Scholar] [CrossRef] [PubMed]
- Kaźmierczak-Siedlecka, K.; Folwarski, M.; Skonieczna-Żydecka, K.; Ruszkowski, J.; Makarewicz, W. The use of Lactobacillus plantarum 299v (DSM 9843) in cancer patients receiving home enteral nutrition—Study protocol for a randomized, double-blind, and placebo-controlled trial. Nutr. J. 2020, 19, 98. [Google Scholar] [CrossRef] [PubMed]
- Rudzki, L.; Ostrowska, L.; Pawlak, D.; Małus, A.; Pawlak, K.; Waszkiewicz, N.; Szulc, A. Probiotic Lactobacillus Plantarum 299v decreases kynurenine concentration and improves cognitive functions in patients with major depression: A double-blind, randomized, placebo controlled study. Psychoneuroendocrinology 2019, 100, 213–222. [Google Scholar] [CrossRef]
- Ahn, H.Y.; Kim, M.; Ahn, Y.T.; Sim, J.H.; Choi, I.D.; Lee, S.H.; Lee, J.H. The triglyceride-lowering effect of supplementation with dual probiotic strains, Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032: Reduction of fasting plasma lysophosphatidylcholines in nondiabetic and hypertriglyceridemic subjects. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Ahn, H.Y.; Kim, M.; Chae, J.S.; Ahn, Y.T.; Sim, J.H.; Choi, I.D.; Lee, S.H.; Lee, J.H. Supplementation with two probiotic strains, Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032, reduces fasting triglycerides and enhances apolipoprotein A-V levels in non-diabetic subjects with hypertriglyceridemia. Atherosclerosis 2015, 241, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.; Lee, Y.J.; Kim, M.; Kim, M.; Kwak, J.H.; Lee, J.-W.; Ahn, Y.-T.; Sim, J.-H.; Lee, J.H. Supplementation with two probiotic strains, Lactobacillus curvatus HY7601 and Lactobacillus plantarum KY1032, reduced body adiposity and Lp-PLA2 activity in overweight subjects. J. Funct. Foods 2015, 19, 744–752. [Google Scholar] [CrossRef]
- Kim, M.; Kang, M.; Yoo, H.J.; Kim, M.S.; Ahn, Y.T.; Sim, J.H.; Jee, S.H.; Lee, J.H. Effects of weight loss using supplementation with Lactobacillus strains on body fat and medium-chain acylcarnitines in overweight individuals. Food Funct. 2017, 8, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Costabile, A.; Buttarazzi, I.; Kolida, S.; Quercia, S.; Baldini, J.; Swann, J.R.; Brigidi, P.; Gibson, G.R. An in vivo assessment of the cholesterol-lowering efficacy of Lactobacillus plantarum ECGC 13110402 in normal to mildly hypercholesterolaemic adults. PLoS ONE 2017, 12, e0187964. [Google Scholar] [CrossRef]
- Madempudi, R.S.; Ahire, J.J.; Neelamraju, J.; Tripathi, A.; Nanal, S. Efficacy of UB0316, a multi-strain probiotic formulation in patients with type 2 diabetes mellitus: A double blind, randomized, placebo controlled study. PLoS ONE 2019, 14, e0225168. [Google Scholar] [CrossRef]
- Sudha, M.R.; Ahire, J.J.; Jayanthi, N.; Tripathi, A.; Nanal, S. Effect of multi-strain probiotic (UB0316) in weight management in overweight/obese adults: A 12-week double blind, randomised, placebo-controlled study. Benef. Microbes 2019, 10, 855–866. [Google Scholar] [CrossRef]
- Surono, I.S.; Martono, P.D.; Kameo, S.; Suradji, E.W.; Koyama, H. Effect of probiotic L. plantarum IS-10506 and zinc supplementation on humoral immune response and zinc status of Indonesian pre-school children. J. Trace Elem. Med. Biol. 2014, 28, 465–469. [Google Scholar] [CrossRef]
- Kusumo, P.D.; Bela, B.; Wibowo, H.; Munasir, Z.; Surono, I.S. IS-10506 supplementation increases faecal sIgA and immune response in children younger than two years. Benef. Microbes 2019, 10, 245–252. [Google Scholar] [CrossRef]
- Prakoeswa, C.R.S.; Herwanto, N.; Prameswari, R.; Astari, L.; Sawitri, S.; Hidayati, A.N.; Indramaya, D.M.; Kusumowidagdo, E.R.; Surono, I.S. Lactobacillus plantarum IS-10506 supplementation reduced SCORAD in children with atopic dermatitis. Benef. Microbes 2017, 8, 833–840. [Google Scholar] [CrossRef]
- Han, Y.; Kim, B.; Ban, J.; Lee, J.; Kim, B.J.; Choi, B.S.; Hwang, S.; Ahn, K.; Kim, J. A randomized trial of Lactobacillus plantarum CJLP133 for the treatment of atopic dermatitis. Pediatr. Allergy Immunol. 2012, 23, 667–673. [Google Scholar] [CrossRef] [PubMed]
- Harima-Mizusawa, N.; Kano, M.; Nozaki, D.; Nonaka, C.; Miyazaki, K.; Enomoto, T. Citrus juice fermented with Lactobacillus plantarum YIT 0132 alleviates symptoms of perennial allergic rhinitis in a double-blind, placebo-controlled trial. Benef. Microbes 2016, 7, 649–658. [Google Scholar] [CrossRef] [PubMed]
- Harima-Mizusawa, N.; Kamachi, K.; Kano, M.; Nozaki, D.; Uetake, T.; Yokomizo, Y.; Nagino, T.; Tanaka, A.; Miyazaki, K.; Nakamura, S. Beneficial effects of citrus jiuce fermented with Lactobacillus plantarum YIT0132 on atopic dermatitis: Results of daily intake by adult patients in two open trials. Biosci. Microbiota Food Health 2016, 35, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Harima-Mizusawa, N.; Iino, T.; Onodera-Masuoka, N.; Kato-Nagaoka, N.; Kiyoshima-Shibata, J.; Gomi, A.; Shibahara-Sone, H.; Kano, M.; Shida, K.; Sakai, M.; et al. Beneficial effects of citrus juice fermented with Lactobacillus plantarum YIT 0132 on Japanese cedar pollinosis. Biosci. Microbiota Food Health 2014, 33, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.C.; Pan, C.H.; Wei, C.C.; Huang, H.Y. Lactobacillus plantarum PS128 improves physiological adaptation and performance in triathletes through gut microbiota modulation. Nutrients 2020, 12, 315. [Google Scholar] [CrossRef]
- Huang, H.Y.; Tsao, S.P. Long term supplementation of Lactobacillus plantarum PS128 attenuated high-intensity exercise induced acute-phase inflammation in triathletes. Curr. Dev. Nutr. 2020, 4 (Suppl. 2), 1564. [Google Scholar] [CrossRef]
- Huang, W.C.; Wei, C.C.; Huang, C.C.; Chen, W.L.; Huang, H.Y. The beneficial effects of Lactobacillus plantarum PS128 on high-intensity, exercise-induced oxidative stress, inflammation, and performance in triathletes. Nutrients 2019, 11, 353. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.W.; Liong, M.T.; Chung, Y.E.; Huang, H.Y.; Peng, W.S.; Cheng, Y.F.; Lin, Y.S.; Wu, Y.Y.; Tsai, Y.C. Effects of Lactobacillus plantarum PS128 on children with autism spectrum disorder in Taiwan: A randomized, double-blind, placebo-controlled trial. Nutrients 2019, 11, 820. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, J.; Guo, Z.; Kwok, L.; Ma, C.; Zhang, W.; Lv, Q.; Huang, W.; Zhang, H. Effect of oral consumption of probiotic Lactobacillus planatarum P-8 on fecal microbiota, SIgA, SCFAs, and TBAs of adults of different ages. Nutrition 2014, 30, 776–783. [Google Scholar] [CrossRef]
- Lew, L.C.; Hor, Y.Y.; Yusoff, N.A.A.; Choi, S.B.; Yusoff, M.S.B.; Roslan, N.S.; Ahmad, A.; Mohammad, J.A.M.; Abdullah, M.F.I.L.; Zakaria, N.; et al. Probiotic Lactobacillus plantarum P8 alleviated stress and anxiety while enhancing memory and cognition in stressed adults: A randomised, double-blind, placebo-controlled study. Clin. Nutr. 2019, 38, 2053–2064. [Google Scholar] [CrossRef]
- Huang, W.C.; Lee, M.C.; Lee, C.C.; Ng, K.S.; Hsu, Y.J.; Tsai, T.Y.; Young, S.L.; Lin, J.S.; Huang, C.C. Effect of Lactobacillus plantarum TWK10 on exercise physiological adaptation, performance, and body composition in healthy humans. Nutrients 2019, 11, 2836. [Google Scholar] [CrossRef]
- Huang, W.C.; Hsu, Y.J.; Li, H.; Kan, N.W.; Chen, Y.M.; Lin, J.S.; Hsu, T.K.; Tsai, T.Y.; Chiu, Y.S.; Huang, C.C. Effect of Lactobacillus plantarum TWK10 on improving endurance performance in humans. Chin. J. Physiol. 2018, 61, 163–170. [Google Scholar] [CrossRef]
- Chong, H.X.; Yusoff, N.A.A.; Hor, Y.Y.; Lew, L.C.; Jaafar, M.H.; Choi, S.B.; Yusoff, M.S.B.; Wahid, N.; Abdullah, M.F.I.L.; Zakaria, N.; et al. Lactobacillus plantarum DR7 improved upper respiratory tract infections via enhancing immune and inflammatory parameters: A randomized, double-blind, placebo-controlled study. J. Dairy Sci. 2019, 102, 4783–4797. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Chong, H.X.; Chung, F.Y.; Li, Y.; Liong, M.T. DR7 Modulated bowel movement and gut microbiota associated with dopamine and serotonin pathways in stressed adults. Int. J. Mol. Sci. 2020, 21, 4608. [Google Scholar] [CrossRef]
- Chong, H.X.; Yusoff, N.A.A.; Hor, Y.Y.; Lew, L.C.; Jaafar, M.H.; Choi, S.B.; Yusoff, M.S.B.; Wahid, N.; Abdullah, M.F.I.L.; Zakaria, N.; et al. DR7 alleviates stress and anxiety in adults: A randomised, double-blind, placebo-controlled study. Benef. Microbes 2019, 10, 355–373. [Google Scholar] [CrossRef]
- Higashikawa, F.; Noda, M.; Awaya, T.; Nomura, K.; Oku, H.; Sugiyama, M. Improvement of constipation and liver function by plant-derived lactic acid bacteria: A double-blind, randomized trial. Nutrition 2010, 26, 367–374. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.H.; Park, S.; Paik, J.W.; Chae, S.W.; Kim, D.H.; Jeong, D.G.; Ha, E.; Kim, M.; Hong, G.; Park, S.H.; et al. Efficacy and safety of Lactobacillus plantarum C29-fermented soybean (DW2009) in individuals with mild cognitive impairment: A 12-week, multi-center, randomized, double-blind, placebo-controlled clinical trial. Nutrients 2019, 11, 305. [Google Scholar] [CrossRef]
- Veiga, P.; Suez, J.; Derrien, M.; Elinav, E. Moving from probiotics to precision probiotics. Nat. Microbiol. 2020, 5, 878–880. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Strain Names | Isolation Source | Plantaricins Names | Sensitive Microbes | Reference |
|---|---|---|---|---|
| Lpb. plantarum ST28MS | Molasses | Plantaricin ST28 MS | Lacticaseibacillus casei, Staphylococcus aureus, Enterococcus faecalis, Pseudomonas aeruginosa, Escherichia coli and Acinetobacter baumanii | [45] |
| Lpb. plantarum LMG 2379 | Wine | Plantaricin W | Gram-positive bacteria including S. aureus, Listeria innocua and E. faecalis | [50] |
| Lpb. plantarum C19 | Fermented cucumber | Plantaricin C19 | Listeria grayi | [51] |
| Lpb. plantarum 423 | Sorghum beer | Plantaricin 423 | Bacillus cereus, Clostridium sporogenes, E. faecalis, Listeria spp. and Staphylococcus spp. | [52] |
| Lpb. plantarum ST8SH | Salami | Plantaricin ST8 SH | Enterococcus spp., Lactobacillus spp., Listeria spp., Streptococcus spp. and Klebsiella pneumonia | [53] |
| Lpb. plantarum UG1 | Dry sausage | Plantaricin UG1 | Listeria monocytogenes | [55] |
| Lpb. plantarum LTF 154 | Fermented sausage | Plantaricin 154 | E. faecalis, Bacillus spp., Staphylococcus spp. and Salmonella enterica serovar Typhimurium | [56] |
| Lpb. plantarum SA6 | Fermented sausage | Plantaricin SA6 | Lpb. plantarum, Levilactobacillus brevis, Leuconostoc spp. and L. grayi | [57] |
| Lpb. plantarum TMW 1.25 | Sausage fermentation | Plantaricin 1.25L | Lactobacillus spp. | [58] |
| Lpb. plantarum BF001 | Spoiled catfish fillets | Plantaricin F | S. aureus, S. Typhimurium, L. monocytogenes, and P. aeruginosa | [59] |
| Lpb. plantarum LL441 | Cabrales cheese | Plantaricin C | Gram-positive bacteria including Bacillus subtilis, E. faecalis, C. sporogenes, and Clostridium tyrobutyricum | [60,61] |
| Lpb. plantarum LC74 | Crude goat’s milk | Plantaricin LC74 | Lpb. plantarum, Lev. brevis, Lentilactobacillus buchneri, Leuc. paramesenteroides, Bacillus stearothermophilus | [62] |
| Lpb. plantarum K25 | Kimchi | Plantaricin K25 | B. cereus and L. monocytogenes | [63] |
| Lpb. plantarum ST31 | Sourdough | Plantaricin ST31 | Lactobacillus spp., Leuconostoc spp., Pediococcus spp., Streptococcus spp., Bacillus spp. and S. aureus | [64] |
| Lpb. plantarum 510 | Koshu vineyard | Plantaricin Y | L. monocytogenes, Weissella spp., Lactococcus lactis, Streptococcus salivarius and B. subtilis | [65] |
| Lpb. plantarum LPCO10 | Green olives fermentations | Plantaricin S Plantaricin T | Clostridium spp., Propionibacterium spp, E. faecalis | [66] |
| Lpb. plantarum 163 | Fermented vegetables | Plantaricin 163 | S. aureus, L. monocytogenes, Bacillus pumilus, B. cereus, Micrococcus luteus, Streptococcus thermophilus, Lacticaseibacillus rhamnosus, E. coli, P. aeruginosa and Pseudomonas fluorescens | [67] |
| Lpb. plantarum JLA-9 | Fermented cabbage | Plantaricin JLA-9 | Bacillus spp., Clostridium spp., S. aureus, M. luteus, P. fluorescens, Serratia marcescens, E. coli, Salmonella spp. and Proteus mirabilis | [68] |
| Lpb. plantarum C-11 | Fermented cucumber | Plantaricin A Plantaricin EF Plantaricin JK | LAB species such as Lactobacillus spp., Pediococcus spp., Leuconostoc spp. and Streptococcus spp. | [69,70] |
| Lpb. plantarum ZJ008 | Fresh milk | Plantaricin ZJ008 | Streptomyces citreus, M. luteus, S. aureus, E. coli, Shigella flexneri, Vibrio parahaemolyticus, L. monocytogenes and P. aeruginosa | [71] |
| Lpb. plantarum ZJ5 | Fermented mustard | Plantaricin ZJ5 | S. aureus, L. monocytogenes, S. flexneri, P. aeruginosa, Shigella dysenteriae, E. coli and Salmonella spp. | [72] |
| Lpb. plantarum NRIC 149 | Pineapple | Plantaricin 149 | Enterococcus hirae, Pediococcus acidilactici, Pediococcus cerevisiae, Lactobacillus spp. | [73] |
| Lpb. plantarum BFE 905 | Ready-to-eat salad | Plantaricin D | Latilactobacillus sakei and L. monocytogenes | [74] |
| Lpb. plantarum OL15 | Algerian fermented olives | Plantaricin OL15 | Lactobacillus spp., Lactococcus spp. and Propionibacterium spp. | [75] |
| Lpb. plantarum DL3 | Chinese pickled cabbage | Plantaricin DL3 | P. aeruginosa, P. fluorescens, Shewanella putrefaciens, Psychrobacter spp., L. monocytogenes, B. cereus, Bacillus licheniformis | [76] |
| Lpb. plantarum Q7 | Yak yogurt | Plantaricin Q7 | L. monocytogenes, S. aureus, E. coli, P. fluorescens, P. putida, P. aeruginosa, Shigella flexneri, Shigella sonnei, S. Typhimurium | [77] |
| Lpb. plantarum KLDS1 | Chinese fermented cream | Plantaricin MG | L. monocytogenes, S. aureus, S. Typhimurium and E. coli | [78] |
| Lpb. plantarum LPL-1 | Fermented fish | Plantaricin LPL-1 | L. monocytogenes, S. aureus, Bacillus amyloliquefaciens, B. pumilus, E. faecalis, Lactobacillus spp., Lact. lactis | [79,80,81] |
| Lpb. plantarum GZ1-27 | Traditional kipper | Plantaricin GZ1-27 | Brochothrix thermosphacta, P. fluorescens, A. baumannii, B. cereus, S. aureus, S. Typhimurium, L. monocytogenes and E. coli. | [82] |
| Lpb. plantarum SLG1 | Yak cheese | Plantaricin SLG1 | B. subtilis, B. cereus, Bacillus megaterium, M. luteus, B. thermosphacta, Clostridium butyricum, S. aureus, L. innocua, L. monocytogenes, E. coli, P. aeruginosa, Enterobacter cloacae, Salmonella paratyphi, Saccharomyces cerevisiae and Candida albicans | [83] |
| Lpb. plantarum BM1 | Fermented Chinese meat product | Plantaricin BM1 | E. faecalis, L. monocytogenes, Lpb. pentosus, Lpb. plantarum, Shigelladysenteriae, E. coli, S. aureus, S. Enteritidis | [84] |
| Lpb. plantarum MBSa4 | Brazilian salami | Plantaricin MBSa4 | S. aureus, L. innocua, Listeria welshimeri, L. monocytogenes, E. hirae, E. faecium, Limosilactobacillus fermentum, Lat. sakei, Penicillium roqueforti, Penicillium expansum, Fusarium sp., Mucor plumbeus, Cladosporium sp., Debariomyces hansenii | [85] |
| Bacteria | Origin | Dose | Health Condition | Subjects and Timeline | Main Impact | Main Outcomes | Reference |
|---|---|---|---|---|---|---|---|
| L. curvatus HY7601 Lpb. plantarum KY1032 | Korean traditional fermented cabbage | 5 × 109 CFU/d in powder | Hypertriglyceridemia | 92 adults 12 weeks | Cholesterol-lowering effect | Triglyceride-lowering effects through reductions in plasma metabolites, fatty acid primary amides and lysoPC | [185] |
| 0.5 × 1010 CFU/d in powder | Hypertriglyceridemia | 128 adults 12 weeks | Cholesterol-lowering effect | reduction of triglycerides and increase of apo A-V and LDL particle size | [186] | ||
| 5 × 109 CFU/d in powder | Overweight subjects | 120 adults 12 weeks | Weight loss | Reductions in body weight, body fat percentage and body fat mass | [187] | ||
| 5 × 109 CFU/d in powder | Overweight subjects | 66 adults 12 weeks | Weight loss | Weight loss and adiposity reduction associated with an increase in medium-chain acylcarnitines | [188] | ||
| Lpb. plantarum ECGC 13110402 | Dairy isolate | 4 × 109 CFU/day in capsules | Hypercholesterolaemia | 49 adults 12 weeks | Cholesterol-lowering effect | Reduction in LDL and Total cholesterol, reduction of triacylgycerides and increase of HDL in over 60′s | [189] |
| UB0316 containing Lpb. plantarum UBLP-40 | Fermented foods | 5 × 109 CFU/day in capsules | Type 2 diabetes mellitus | 79 adults 12 weeks | Weight loss | Weight loss, improvements in glycemic control via reduction in HbA1c levels | [190] |
| 5 × 109 CFU/day in capsules | Overweight/obesity conditions | 71 adults 12 weeks | Weight loss | Reduction of BMI, body weight and WHR in overweight/obese adults | [191] | ||
| 2 × 1011 CFU/day Microencapsulated in powder | Healthy preschool children | 48 one-two years old children 90 days | Immunomodulation | Increased humoral immune response, as well as improved zinc status | [192] | ||
| Lpb. plantarum IS-10506 | dadih Indonesian traditional fermented buffalo milk | 2.3 × 1010 CFU/day Microencapsulated in powder | Immature intestinal immune system | 38 one-two years old children 90 days | Immunomodulation | Stimulation of TGF-β1, increased sIgA production, with a significant correlation between TGF-β1/TNF-α and fecal sIgA | [193] |
| 1010 CFU/day Microencapsulated in powder | Atopic dermatitis | 22 children 12 weeks | Immunomodulation Anti-inflammatory activity | SCORAD and levels of IL-4, IFN-γ, and IL-17 were significantly lower decrease in clinical Symptoms through down regulation of Th2 adaptive immune response | [194] | ||
| Lpb.plantarum CJLP133 | Kimchi | 1010 CFU/day powder in airtight alu-bags | Atopic dermatitis | 83 children 16 weeks | Immunomodulation Anti-inflammatory activity | Reduced SCORAD score, eosinophil count and cytokines levels (IL-4 and IFN-γ) | [195] |
| 8 × 1010 cells/day in fermented citrus juice | Perennial allergic rhinitis | 33 adults 8 weeks | Immunomodulation with antillaergic effects | Reduction of nasal symptoms. attenuation of Th2 cells, total IgE and ECP | [196] | ||
| Lpb.plantarum YIT 0132 | Fermented foods | 8 × 1010 cells/day in fermented citrus juice | Atopic dermatitis | 32 + 18 adults 8 + 8 weeks | Immunomodulation with antillaergic effects | Reduced symptoms with immunomodulatory effect via attenuation of IgE and ECP | [197] |
| 8 × 1010 cells/day in fermented citrus juice | Japanese Cedar Pollinosis | 42 adults 8 weeks | Antiallergic effects | Reduction of allergy symptoms | [198] | ||
| 3 ×1010 CFU/day in capsules | Triathlete’s microbiota | 20 Triathletes 4 weeks | Endurance performance amelioration via gut microbiota | GI health and physiological homeostasis maintenancenduring endurance exercise through functional microbiota modulation | [199] | ||
| Lpb. plantarum PS128 | Fu-tsai Taiwan fermented mustard greens | 3 × 1010 CFU/day in capsules | Exercise-induced inflammation | N.R. | Antioxidative and anti-inflammatory activities | Oxidative stress and inflammation alleviation | [200] |
| 3 × 1010 CFU/day in capsules | Exercise-Induced oxidative stress and inflammation | 18 Triathletes 8 weeks | Antioxidative and anti-inflammatory activities | Oxidative stress alleviation, decreased pro-inflammatory parameters, enhanced exercise performance | [201] | ||
| 3 × 1010 CFU/day in capsules | Autism spectrum disorder | 80 children 4 weeks | Mental health | Age-dependent amelioration of autism symptoms | [202] | ||
| Lpb. plantarum P8 | Traditionally Mongolian fermented sour milk | 6 × 1010 CFU/day in tablets | Healthy adults | 33 adults 4 weeks | Gut microbiota and Immunomodulation | Time- and age- related changes in fecal sIgA, TBAs, and SCFAs levels Beneficial alteration of gut microbiota | [203] |
| 2 × 1010 CFU/day powder in sachet | Mental stress conditions | 103 adults 12 weeks | Antistress and anti-inflammatory activities | Alleviation of selected stress, anxiety, memory and cognitive symptoms in stressed adults with reduction of pro-inflammatory markers and enhanced memory and cognitive traits | [204] | ||
| Lpb. plantarum TWK10 | Taiwan pickled cabbage | 3 × 1010 CFU/day 9 × 1010 CFU/day in capsules | Exercise physiology | 54 adults 6 weeks | Endurance performance amelioration | Enhanced exercise performance in a dose-dependent manner correlated with better physiological adaptation (body fat significantly decreased and muscle mass significantly increased) | [205] |
| 1 × 1011 CFU/day in capsules | Endurance performance | 16 adults 6 weeks | Endurance performance amelioration | Significantly higher endurance performance and glucose content | [206] | ||
| Lpb. plantarum DR7 | Cow milk | 109 CFU/day powder in aluminium sachet | Upper respiratory tract infections | 109 adults 12 weeks | Immunomodulation with anti-inflammatory activities | Improvements of nasal symptoms and URTI frequency by improving inflammatory parameters and enhancing immunomodulatory properties | [207] |
| 109 CFU/day powder in aluminium sachet | Mental stress conditions | 124 adults 12 weeks | Antistress and anti-inflammatory activities | Modulation of stress-induced bowel movement and gut microbiota in association with dopamine and serotonin pathways | [208] | ||
| 109 CFU/day powder in aluminium sachet | Mental stress conditions | 111 adults 12 weeks | Antistress and anti-inflammatory activities | Reduced plasma cortisol and pro-inflammatory cytokines. Reduction of stress and anxiety, improved cognitive and memory functions with stimulation of serotonin and dopamine-norepinephrine pathway | [209] | ||
| Lpb. plantarum SN13T | Plant-derived | 2 × 108 CFU/g in yogurt | Constipation | 68 adults 6 weeks | Gut functions improvement | Constipation restoring effects with improved serum lipid contents and liver functionality | [210] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Gonzalez, N.; Battista, N.; Prete, R.; Corsetti, A. Health-Promoting Role of Lactiplantibacillus plantarum Isolated from Fermented Foods. Microorganisms 2021, 9, 349. https://doi.org/10.3390/microorganisms9020349
Garcia-Gonzalez N, Battista N, Prete R, Corsetti A. Health-Promoting Role of Lactiplantibacillus plantarum Isolated from Fermented Foods. Microorganisms. 2021; 9(2):349. https://doi.org/10.3390/microorganisms9020349
Chicago/Turabian StyleGarcia-Gonzalez, Natalia, Natalia Battista, Roberta Prete, and Aldo Corsetti. 2021. "Health-Promoting Role of Lactiplantibacillus plantarum Isolated from Fermented Foods" Microorganisms 9, no. 2: 349. https://doi.org/10.3390/microorganisms9020349
APA StyleGarcia-Gonzalez, N., Battista, N., Prete, R., & Corsetti, A. (2021). Health-Promoting Role of Lactiplantibacillus plantarum Isolated from Fermented Foods. Microorganisms, 9(2), 349. https://doi.org/10.3390/microorganisms9020349