Abstract
Although known as causes of community-acquired pneumonia and Pontiac fever, the global burden of infection caused by Legionella species other than Legionella pneumophila is under-recognised. Non-L. pneumophila legionellae have a worldwide distribution, although common testing strategies for legionellosis favour detection of L. pneumophila over other Legionella species, leading to an inherent diagnostic bias and under-detection of cases. When systematically tested for in Australia and New Zealand, L. longbeachae was shown to be a leading cause of community-acquired pneumonia. Exposure to potting soils and compost is a particular risk for infection from L. longbeachae, and L. longbeachae may be better adapted to soil and composting plant material than other Legionella species. It is possible that the high rate of L. longbeachae reported in Australia and New Zealand is related to the composition of commercial potting soils which, unlike European products, contain pine bark and sawdust. Genetic studies have demonstrated that the Legionella genomes are highly plastic, with areas of the chromosome showing high levels of recombination as well as horizontal gene transfer both within and between species via plasmids. This, combined with various secretion systems and extensive effector repertoires that enable the bacterium to hijack host cell functions and resources, is instrumental in shaping its pathogenesis, survival and growth. Prevention of legionellosis is hampered by surveillance systems that are compromised by ascertainment bias, which limits commitment to an effective public health response. Current prevention strategies in Australia and New Zealand are directed at individual gardeners who use potting soils and compost. This consists of advice to avoid aerosols generated by the use of potting soils and use masks and gloves, but there is little evidence that this is effective. There is a need to better understand the epidemiology of L. longbeachae and other Legionella species in order to develop effective treatment and preventative strategies globally.
1. Introduction
Following the discovery of Legionella pneumophila as the cause of the Legionnaires’ disease outbreak in Philadelphia in 1976 [1], it was quickly realised that there were other species within the genus Legionella, some of which could also cause human disease. Nearly 90 different Legionella species have now been reported, of which more than 20 different species are known to be pathogenic in humans [2,3]. A list of species currently known to cause human disease is available online [4]
Despite recognition that a variety of Legionella species and serogroups can cause human disease, L. pneumophila is overwhelmingly reported as the most common cause of legionellosis globally. While the predominance of L. pneumophila may be a true observation, there is inherent diagnostic bias in current testing strategies for legionellosis that favours detection of L. pneumophila over other Legionella species. In particular, many countries rely on the urinary antigen test in cases of suspected Legionnaires’ disease. This test only reliably detects L. pneumophila serogroup 1, so other Legionella species and serogroups may not be tested for at all if this is used as the sole, or predominant testing method. As a consequence, detection of infections caused by Legionella species other than L. pneumophila are usually made by chance, and the true burden and epidemiology of disease is largely unknown. This situation is further complicated by the lack of studies on the microbial aetiology of community-acquired pneumonia that systematically test for all species of Legionella using PCR and/or culture-based methods.
The one exception is Legionella longbeachae, which was first isolated from patients with pneumonia in the USA 40 years ago [5]. In some countries, such as Australia and New Zealand, where the aetiology of community-acquired pneumonia has been systematically studied, L. longbeachae disease has emerged as the predominant cause of legionellosis [6,7]. Cases are increasingly being reported from other regions as well. The countries that have reported at least one case are shown in Figure 1, demonstrating that the conditions for infection may occur in countries with widely varying climates and socioeconomic conditions. However, it is still unclear whether the relatively high incidence of Legionnaires’ disease caused by species other than L. pneumophila in Australia and New Zealand is a true geographic variation or simply reflects that these species are more rigorously tested for in this region.
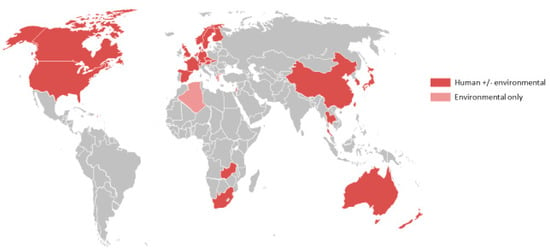
Figure 1.
Reported global distribution of L. longbeachae isolated from human and/or environmental sources [5,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29]. Data from Austria, Norway and Slovakia provided by Julien Beauté, European Centre for Disease Prevention and Control (ECDC); and for the Czech Republic by Vladimir Drašar, Head Public Health Institute Ostrava, National Legionella Reference Laboratory, Czech Republic.
This paper reviews legionellosis caused by Legionella species other than L. pneumophila. The primary focus is on legionellosis caused by L. longbeachae, simply because considerably more data are available on this species. Indeed, apart from L. pneumophila, L. longbeachae is the only species causing legionellosis for which there are sufficient data to make detailed descriptions about clinical features and epidemiology.
2. Clinical Features
The clinical spectrum of legionellosis caused by Legionella species other than L. pneumophila is similar to that caused by L. pneumophila, with pneumonia (Legionnaires’ disease) being the predominant clinical presentation. There are no clinical features that distinguish Legionnaires’ disease caused by L. pneumophila from L. longbeachae, nor can Legionnaires’ disease be it distinguished clinically from other causes of community-acquired pneumonia [30,31]. Increasing age, immune suppression and pre-existing lung disease have been regarded as important predisposing conditions for legionellosis. Recognition of these factors has led some researchers to develop scoring systems to increase recognition of Legionnaires’ disease, but none of these have been developed for non-L. pneumophila species or gained widespread acceptance as a clinical tool [32]. A case series of L. longbeachae disease from New Zealand found that a higher proportion of cases were males (63%), most cases (59%) were of mild to moderate severity (CURB-65 score 0–1), 25% required admission to the intensive care unit, and the mortality rate within 30 days of admission was 5% [31].
Pontiac fever, is transient flu-like illness with a high attack rate, [33] and has been associated with exposure to a variety of Legionella species, including L. pneumophila, Legionella feeleii, Legionella micdadei, Legionella anisa, and L. longbeachae [34,35,36,37,38,39]. Outbreaks of Pontiac fever caused by species other than L. pneumophila have occurred in a variety of occupational and recreational settings, including workers in an automobile manufacturing plant (L. feeleii) [34], users of whirlpool spas (L. micdadei) [35,36,40], workers at a compost manufacturing site (L. longbeachae) [37] and conference delegates and restaurant patrons exposed to decorative fountains (L. anisa) [38,39]. Reports of Pontiac fever are almost certainly underestimated because sporadic cases are unlikely to come to attention and be diagnosed.
Non-L. pneumophila has also been identified in cases of skin and soft tissue infection, septic arthritis and bacterial endocarditis [41,42,43,44,45,46,47,48,49,50]. These cases may be severe and difficult to diagnose as legionellae are unexpected as the infecting organism. A recent example is that of a 78-year-old gardener who pricked her hand on a rose thorn. L. longbeachae was cultured on selective media from surgically debrided tissue that would have been missed with standard investigations [41]. Likewise, diagnosis and treatment of recurrent upper limb abscesses of L. bozemanii was delayed for weeks in a patient on steroid therapy and methotrexate for seronegative arthritis; the diagnosis was eventually made by 16S rRNA PCR and seroconversion [51].
3. Epidemiology
The epidemiology of infection with L. longbeachae and other non-L. pneumophila species of Legionella is poorly described worldwide, although cases have been reported in many countries (Figure 1). A major reason for this is the difficulty with diagnosis, with the likelihood that cases are markedly under-diagnosed and under-recognised. In many centres, testing is primarily based around urinary antigen testing that recognises only L. pneumophila serogroup 1. Culture of respiratory samples is discouraged by some authorities for nonsevere disease, including in hospitalised patients [52]. The results may also be compromised if specimens are taken after empiric therapy (including antilegionella therapy) has been administered. Where other techniques are employed, such as serology, specific culture techniques and PCR, L. longbeachae has emerged as an important cause of Legionnaires’ disease in community-acquired pneumonia. Other non-L. pneumophila species are recognised particularly in severely immunocompromised patients [20,53,54]. The non-L. pneumophila species almost certainly cause many more cases of mild to moderate disease that do not come to medical attention and remain an ongoing burden of morbidity on the population.
3.1. Incidence
Several high-income countries and regions have well established surveillance systems but almost all data on legionellosis relies on various forms of passive surveillance (Table 1). Most of these data are derived from cohort studies or case reports of patients who required admission to hospital. These reports demonstrate striking differences in the number of cases identified in Australia and New Zealand compared with Europe and North America [6,55,56,57,58]. Most cases are sporadic, although clusters of Legionnaires’ disease caused by L. longbeachae have been recognised in Scotland, where six cases of Legionnaires’ disease caused by L. longbeachae were identified over a four-week period in 2013 [59], and in Sweden, where 30 cases were identified during spring/summer 2018 [60].

Table 1.
Number of cases of Legionellosis reported from countries with national surveillance systems.
The highest reported incidence rates of Legionnaires’ disease from any cause and from L. longbeachae have been reported from New Zealand. Since 1999 in the Canterbury region, routine passive surveillance, based on the culture of respiratory samples, demonstrated that L. longbeachae was a major cause of Legionnaires’ disease in New Zealand. To increase the sensitivity of the testing methods, PCR was adopted as the primary diagnostic tool for all species of Legionella in respiratory samples submitted to the diagnostic laboratory in 2010. This technique demonstrated there was a large number of cases that were previously unsuspected, and there was a marked seasonal variation in cases with the peak occurring in spring and early summer [53]. Because there were differences in diagnostic practices around New Zealand, a nationwide study was conducted in 2015–6 using the same method of recruitment to properly assess the nationwide burden [7]. This gave an annualized rate of 5.4 per 100,000 people per year. L. longbeachae was the predominant species identified and was implicated in 150 (63%) of 238 Legionnaires’ disease cases. This is the highest rate of Legionnaires’ disease reported caused by both L. longbeachae and L. pneumophila worldwide.
A major limitation of this study was the reliance on testing only those samples submitted to the laboratory. A small study using enhanced sputum collection of hospitalised patients with community-acquired pneumonia found >40% of pneumonia patients in our study were unable to spontaneously expectorate sputum, but this could be significantly increased to 67% by extra assistance or sputum induction [62]. This increased the case detection of Legionnaires’ disease by 36%. Some cases were almost certainly missed in this study as a dry cough is more common in Legionnaires’ disease than other causes of community-acquired pneumonia [31].
In Australia over the period of 2009 to 2015, the annual notification rate of Legionellosis using passive surveillance was about 1.5 per 100,000 people [6]. L. longbeachae was the more commonly identified (144 to 215 notified cases) compared with L. pneumophila (114 to 228 cases). L. longbeachae was most common (>50%) in the Mediterranean climate of Western Australia and the tropical Northern Territories.
In contrast, L. longbeachae was reported in low numbers elsewhere, making up 0.3–3.6% of cases in the US, 0.7% in Japan, and 1% in Europe (Table 1). There is some preliminary evidence that cases of L. longbeachae are increasing in both Scotland [63] and the Netherlands [64]. There is very little information on the epidemiology of Legionnaires’ disease from low and middle-income countries, but a year-long population-based study of the microbiological causes of clinically defined community-acquired pneumonia requiring hospitalisation in a rural province of Thailand has been reported [27]. Infection with L. longbeachae was identified by seroconversion using an indirect immunofluorescence assay of combined IgG, IgA and IgM titres. These cases were not confirmed by culture or PCR testing. The incidence rate was 5–29 cases per 100,000 of the population, incidence increased with age, and it was more common among males (rate ratio 1.6 95% CI 1.1–2.3). There was also a single case of Legionnaires’ disease caused by L longbeachae reported in a child in Zambia [29]. This was the only case found as part of a year-long study conducted in five locations in Africa and two in Asia of 2757 cases of childhood community-acquired pneumonia.
The epidemiologic evidence of human infection of other non-L. pneumophila species is limited. These infections are rarely reported due, at least in part, to diagnostic bias that precludes cases being identified. There appears to be little difference in the distribution or reported incidence rates of these species across the world. Of these, L. micdadei is the most consistently reported across regions (Table 1). Clusters of cases related to a common source have been described, including 12 cases of Legionnaires’ disease caused by L. micdadei among 38 renal and cardiac transplant patients in one centre over a three-month period, and further cases were detected 16 months later [65]. Subsequently, 16 cases of L. micdadei were reported in another tertiary care centre over a 12 year period, predominantly among immunocompromised hospitalised patients [66]. Another cluster of five cases of nosocomial pneumonia caused by L. bozemanii in a community hospital among immune compromised patients has been identified [67]. Case series have demonstrated that Legionnaires’ disease may be caused by a wide variety of non-L. pneumophila species in immune-compromised patients. The MD Anderson Centre in Houston Texas reported consecutive cases of Legionella infections in cancer patients from 2002 to 2014. Most of the 33 strains identified were on specimens obtained at bronchoscopy, and 18 were non-L. pneumophila species [54]. Twenty-seven patients had underlying hematologic malignancies and 23 were leukopenic. Six patients were recipients of allogeneic hematopoietic stem cell transplant, with their infections caused by five Legionella species. Legionnaires’ disease was the immediate cause of death in six [54]. Some cases may be related to travel but the numbers are difficult to obtain as the species of infecting Legionella species may not be reported [68,69].
3.2. Environmental Studies
3.2.1. Potting Soils and Gardens
An important epidemiological link has been made between exposure to compost or potting soils and L. longbeachae infection. An investigation during 1988–1989, into 22 cases of L. longbeachae infection in South Australia, found cases were among regular gardeners. L. longbeachae was subsequently isolated from compost and potting soils found at the homes of four cases, providing a possible link to a natural habitat for this bacterium [70,71]. Further studies have shown that L. longbeachae infection was linked to the use of potting soils or gardens in other states in Australia, as well as Japan, the United States, Canada, the Netherlands and Scotland through case-series and laboratory evidence [59,64,72,73,74,75,76]. In the Scottish cluster all cases were associated with amateur gardeners and required admission to the intensive care unit [59]. A case control study in South Australia found recent use of potting mix was associated with illness (OR 4·74, 95% CI 1·65–13·55, p = 0·004) [72]. Similarly, exposure to purchased compost was a risk factor for Legionnaires’ disease from L. longbeachae in New Zealand (OR 6.2, 95% CI 2.2–17.3), unlike for homemade compost [77].
Seminal studies by Steele et al. further implicated potting soils as a source of human L. longbeachae infection. L. longbeachae was recovered on selective media from soil samples and potting mixes from four patients who suffered Legionnaires’ disease with this organism. The bacteria were able to persist for seven months in potting soil held at room temperature but did not grow at 43°C or above in soil [70]. Subsequently L. longbeachae was isolated from 26 (56%) of 45 potting mixes manufactured in Australia but not from imported potting soils [78]. L. longbeachae was found in one of four samples of fresh pine sawdust and in a soil sample close to the site, but not hammer milled bark. L. longbeachae was also found at two of six large scale manufacturing plants and in samples from three of 20 home compost heaps [78]. L. Longbeachae has since been detected in potting soils and gardens from many other countries in Europe, North America and Japan, but not as frequently as in Australia [16,79,80,81,82,83,84]. There is a highly diverse pool of L. longbeachae genotypes within the samples from potting soils that makes it difficult to identify the source of a clinical infection [85].
Steele and colleagues also demonstrated that the ecology of Legionella species in potting soils was extremely complex, as multiple species of pathogenic Legionella were usually present in Australia-made potting soils [86]. Among 20 Australian potting soils, L. anisa, L. bozemanii, L. micdadei, L. gormani, L. dumoffi and L. pneumophila serogroups 1–14 were found, in addition to L. longbeachae [78]. A similar range of Legionella species were found in 20 home composts, but with a lower proportion of L. longbeachae present among the 29 isolated (L. pneumophila serogroup 2–14, 11 strains (55%); L. pneumophila SG, 1 strain (5%); L. anisa, 5 strains (25%); L. longbeachae and L. bozemani, three strains each (15%); unidentified, six (30%)) [86]. UK studies of 24 compost products found 15 (62.5%) samples tested contained Legionella species either on first testing or after enrichment by coculture with amoeba when retested 8–10 weeks later. The most common species identified before enrichment was L. sainthelensis (5/24; 21%), while L. longbeachae (4/24; 17%) was the most common after amoebal enrichment. Other identified species known to cause human disease were also isolated (L. anisa, L. bozemanii, L. micdadei, L. gormani, L. dumoffi, L. birminghamensis and L. pneumophila) [73,83]. A similar range of Legionella species have been identified in potting soils in Greece, Switzerland and Japan [16,79,82]. These results suggest that potting soils could be a source of other Legionella infections, including with L. pneumophila species.
L. longbeachae and other non-L. pneumophila species have been found in water contaminated by soil, including rain water puddles in Europe, and to be widely distributed among fresh water and marine waters in Puerto Rico [87,88]. Legionella species were also found in water collected in rain forests from epiphytes 30 feet above ground [87]. L. longbeachae was not found in soil or other environmental samples obtained from around houses and work places of people in Southern Thailand where a high prevalence of L. longbeachae infection had been found by serological testing [89]. Many other Legionella species were found, suggesting one or more of these may have been responsible for the infections.
One possible difference of European potting mixes compared to and Australian and New Zealand products is the range and proportions of the constituents used in the manufacturing process. Australian and New Zealand potting soils contain a significant amount of partially or fully composted pine bark and sawdust in addition to peat, and composted green waste. Recent studies in New Zealand using qPCR identified L. longbeachae DNA in pine bark, pine sawdust and potting soil products containing pine products, but did not identify it in peat or commercially composted green waste [90]. Studies from Europe indicate that Legionella species, including L. pneumophila and L. bozemanii are common in composted green waste, but not fresh green waste. L. longbeachae is rarely found in either fresh or composted green waste [80]. Australian, New Zealand and European studies have failed to find Legionella species in peat. These studies raise the possibility that composting facilities are contaminated with Legionella species from other sites [78,83,90]. Studies of bark from living pine trees in New Zealand found L. longbeachae DNA is common in spring, but very uncommon on bark from other tree species [91]. L. longbeachae could be brought into composting facilities with bark, and the load may contribute to the seasonal variation in clinical cases. In Switzerland, it has been suggested that Legionella species could be brought into the storage facilities by wind and rain [79].
All of these studies indicate that composted material is an important reservoir of pathogenic species of Legionella and are a potential source of human infection, but we are unaware of any substantial data linking cases of Legionnaires’ disease to the manufacture of compost. There is some evidence that compost facilities can release bioaerosols containing Legionella species, and transmission of L. longbeachae aerosol has been suggested as a cause of a nosocomial infection [92,93].
3.2.2. Water Bodies
There have been occasional reports of nosocomial L. longbeachae Legionnaires’ disease associated with detection of this species in hospital and residential water supplies and unpasteurised water tanks [94,95]. L. longbeachae has been identified in samples taken from a cooling tower, but infection from this source was not proven [96]. It has also been isolated from a hot spring water in Algeria by culture and molecular methods [8].
In contrast to L. longbeachae, infection with other non-L. pneumophila species have typically been associated with water bodies, and the epidemiology of water related non-L. pneumophila species is likely to be similar to L. pneumophila. The environmental sources of community acquired Legionnaires’ disease has been extensively reviewed [97]. Nosocomial Legionnaires’ disease with L. micdadei, L. bosemani, L. feelii, and L. anisa has been attributed to infected water distribution systems, including those in hospitals [58,98,99,100]. L. micdadei has also been identified in ultrasonic nebulisers [101]. L. micdadei has been found in whirlpool spas in Norway, and L. anisa associated with a decorative fountain [35].
4. Pathogenesis
The pathogenesis of Legionella infection has been most extensively studied for L. pneumophila, and this has been comprehensively reviewed [102]. Relatively recent advances in high throughput next generation sequencing technologies has allowed the genomes of non-L. pneumophila species to be sequenced and examined in greater depth. As such, comparative genomic analysis has proven to be instrumental in elucidating the factors that are important for the pathogenicity of Legionella species. This has revealed both important parallels, and differences between the various pathogenic factors of Legionella species, which remain under continued investigation.
Analysis of the genomes of up to 58 different Legionella species revealed they are highly dynamic and diverse, with lengths ranging in size from the smallest at 2.37 Mb in Legionella adelaidensis to the largest at 4.82 Mb in Legionella santicrucis [103,104]. Indeed, across the Legionella pan genome only 6% of the genes (1008) constitute the core genome [103,104], illustrating the high genomic diversity and the importance of horizontal gene transfer and recombination in shaping the genome of the genus. At around 4.1 Mbp, the single circular chromosome of L. longbeachae is approximately 500 kb larger than L. pneumophila, but has a similar GC content at around 37% [104,105,106]. The gene organization is also markedly different to L. pneumophila, with limited gene synteny, although there are small clusters where the gene order is highly conserved [105,106].
There are around 3500 predicted coding sequences in the L. longbeachae genome, and around 2000 of these have been assigned putative functions [105,106,107,108,109]. Only 65% of the genes in L. longbeachae are orthologous to those in L. pneumophila [105,110]. The L. longbeachae genome, as a result of its soil and plant habitat, contains numerous genes that reflect its ecological niche, with several genes that encode proteins that can degrade components of plant cell walls, such as cellulases, hemicellulases and pectin lyases [105]. In addition, the genome is highly methylated with m4 C and m6 A epigenetic modifications throughout the genome, where m6 A is strongly associated with particular sequence motifs [109]. Although the L. longbeachae genome appears to be more stable than that of L. pneumophila [105], large-scale rearrangements have occurred in some regions of the chromosome with evidence of extensive recombination [85].
In the accessory genome, L. longbeachae strains NSW150 and D4968 [106] were both found to contain a similar plasmid of around 70 kb. It was first thought that L. longbeachae plasmids only circulated within this species and were distinct from those found in L. pneumophila, which range in size from around 59 kb to 131 kb [110]. More recently, sequencing data from multiple clinical and environmental L. longbeachae isolates has shown that most contain at least one plasmid ranging in size from around 70 kb to 150 kb [85,109]. Despite still relatively limited data, the plasmids have been shown to have a common backbone including conjugational genes separated by variable regions. Importantly, there is evidence of extensive recombination and horizontal gene transfer between the plasmids from different species, indicating both intra and interspecies exchange of genetic material across the genus [85,107,108,109].
Transcriptome analysis of L. pneumophila and L. longbeachae has shown that both have a biphasic lifestyle (a replicative/avirulent phase and a transmissive/virulent phase), although this is less pronounced in L. longbeachae [105]. Both also typically exist in biofilm forms or intracellularly with amoeba or macrophages. Conditions which disrupt the biofilm may release large numbers of organisms into the environment [111]. Likewise, the ability to multiply within free-living amoeba may facilitate transmission, as a single amoebal cyst may contain thousands of Legionella bacteria [93]. The major route of infection is probably by inhalation of aerosols derived from soil or water, but may also reach the lung by microaspiration [112]. The infectious dose has not been precisely defined and may be higher than 1000 organisms [113]. This is almost certainly lower in people with pre-existing lung disease that disrupts the mucociliary escalator, allowing the bacterium to reach the alveolar space. Immune deficiencies, particularly cell mediated deficiencies, such as haematological malignancy, as well as immunosuppressive therapy such as dose steroid therapy, tumor necrosis factor inhibitors and anti CD-52 agents, are also important risk factors for developing disease from Legionella species [114,115,116]. Mucosal associated invariant T (MAIT) cells that are resident in the lung, also appear to contribute to protection against fatal infection with Legionella. The mechanism is dependent on the major histocompatibility complex (MHC) class I-related molecule MR1, interferon-γ (IFN-γ) and granulocyte macrophage-colony stimulating factor (GM-CSF) [117].
On reaching the alveolar space L. pneumophila is known to attach to alveolar macrophages using flagella, pili and outer membrane porins [118,119,120,121,122,123]. The attachment enables phagocytosis to follow, mediated by macrophage infectivity potentiator protein (MIP) and human complement C3. In contrast L. longbeachae lack flagella but have other effector proteins, possibly derived from organisms that typically live in soil and plant derived material. The lack of flagella has been cited as one of the reasons for the different responses to infection in different strains of mice, where A/J mice are susceptible to L. pneumophila infection and replication, but C57BL/6 and BALB/c mice are not [110,124]. In contrast, all three mouse strains are susceptible to L. longbeachae infection and replication. It is thought that the flagellin of L. pneumophila triggers pyroptosis and cell death in the C57BL/6 and BALB/c mice, leading to a failure to establish infection [110,124].
Following phagocytosis, Legionella infection, survival and replication within a host cell is dependent on successfully transforming the phagosome into a distinct membrane bound vacuole, termed the Legionella containing vacuole (LCV) [102,125]. To do this Legionella employ various secretion systems to translocate effector proteins into the vacuole, which, in turn, inhibit phagocyte lysosome fusion [102]. The effector proteins also facilitate the transformation of the vacuole into a nutrient rich environment, as they enable the host resources to be hijacked and transported back to the LCV to support legionella replication. As the nutrients are depleted by bacterial replication, the organism enters stationary phase, triggers apoptosis of the host cell releasing the organisms to be phagocytosed and enters a new replicative cycle.
The various secretion systems and the effector proteins of Legionella are the predominant drivers of their pathogenicity. Comparative genomic analysis has shown that all species analysed to date contain the genes for both a type 2 secretion system (T2SS) and a type 4B secretion system (T4BSS), also known as the Icm/Dot secretion system [102,103,126]. In addition to these, and similarly to L. pneumophila, L. longbeachae contains the genes for a type 4A secretion system (T4ASS) but, like all other non-L. pneumophila species, it lacks a type 1 secretion system [126]. The Icm/Dot T4BSS is crucial for LCV biosynthesis and intracellular replication. It is also the primary and most well-studied pathogenic secretion system of Legionella [104,125,126,127], particularly in L. pneumophila. The components are encoded by 25 genes that occur within two distinct pathogenicity regions on the chromosome, and the gene order and orientation is highly conserved genus-wide [104,126,127]. The Icm/Dot T4BSS translocates effector proteins into the host cell that interact with the host to subvert resources and cell function for the benefit of the bacterium and is indispensable for pathogenicity. Despite the highly conserved nature of the Icm/Dot T4BSS, the effector repertoire that it translocates into the host has been shown to be highly variable across the various species examined to date [103,104]. From species to species, the effectors differ in number and, although they share functional similarities, there is little DNA sequence homology [128]. Of the over 300 effectors identified in L. pneumophila, only 35% are found to occur in L. longbeachae [125]. Genus-wide, comparative analysis of 38 species has revealed even greater variability with only seven putative core effectors identified [104], i.e., effectors that were found to have orthologs in each of the species examined out of a total of 5885. More recently, Gomez-Valero et al., 2018, after examining 80 strains from 58 different Legionella species showed that the pan-genus pool of putative Icm/Dot T4BSS effectors could be well over 18,000 proteins, again illustrating the extremely high diversity of the effector proteins in Legionella. Furthermore, analysis of these putative effector genes revealed that they encode secreted proteins that contain a large number and variety of eukaryotic domain-encoding proteins and eukaryotic-like proteins [102,103]. The genes encoding these proteins are thought to have been acquired via horizontal gene transfer from their protozoan hosts in a type of coevolutionary arms race that allows Legionella to mimic the host to establish infection and successfully replicate.
The survival tactics and pathogenic trajectory of Legionella is significantly shaped by the interactions and exchanges that it has with its protozoan hosts, but not by interactions it has with its human host [129,130,131]. This is because human infection by Legionella is an evolutionary dead-end; either the bacterium dies because of successful response/treatment or the human host dies because of the infection, which consequently causes the bacterium to die as it does not have recourse to the external environment. Therefore, infectivity and pathogenicity to humans is not an evolutionary advantage to Legionella. While L. pneumophila has been found to exhibit a broad host range that spans several protozoan phyla [129], little is known about the protozoan hosts of other Legionella species. Nonetheless, it has been shown in various coculture studies that the protozoan host species has a major influence on L. pneumophila’s ability to infect human cells [132], and it is reasonable to expect that the same is true for other Legionella species. For instance, L. pneumophila cocultured in Acathamoeba polyphaga is more tolerant to antibiotics, while resistance to chlorination is higher in those grown in Vermamoeba vermiformis when compared to those from Acathoamoeba castillanii [132].
5. Prevention and Public Health
The public health aspects most specific to legionellosis caused by Legionella species other than L. pneumophila relate to L. longbeachae and exposure to gardening activities. The public health response to the threat posed by potting soils in New Zealand and Australia has been to place responsibility onto the individual gardener. Gardeners are alerted to the risk with warnings printed on potting soil containers and information leaflets available at the point of sale. Information on Legionnaires’ disease can be found on the Ministry of Health or State Health authorities’ websites. Gardeners, particularly those with other health conditions or advanced age, are advised to protect themselves by using gloves and masks and to dampen the potting soil when the bag is opened to minimise aerosol formation [31,70].
This advice makes intuitive sense as inhalation of aerosols is probably the most important means of transmission and is the major route of infection of L. pneumophila. This is supported by results of a case control study among gardeners that found that tipping or trowelling potting soils was a major risk factor for Legionnaires’ disease (population attributable fraction 65%) [77]. However, behaviours that involve touching the face, such as eating or drinking without handwashing, or smoking, may also place gardeners at risk of infection (population attributable fraction 35%) [77]. These results suggest that eliminating aerosolization activities and effective hand hygiene could reduce the risk of Legionnaires’ disease but, disappointingly, the study did not find any protective effect of wearing a mask or gloves [77].
There has been little research on the acceptability or effectiveness on the use of personal protective equipment for preventing Legionnaires’ disease while gardening. Multiple barriers to the use of face masks have been identified in other settings with serious health risks, such as tuberculosis and influenza, and contribute to poor acceptability among gardeners [133,134,135]. These include lack of perceived risk, discomfort and poor fit. Discomfort may be compounded when masks become dampened, as this increases the work of breathing and the masks subsequently lose their protective effect. Alongside discomfort, masks may also may give a false sense of security and interfere with communication, and may also accumulate bacteria and become hazardous prior to disposal [136]. It may be possible to change the behaviour of gardeners as the COVID-19 pandemic has changed public perception of the value of masks [135].
Other avenues to change gardeners’ behaviour to reduce Legionnaires’ disease risk are still to be explored. These could include warning about the risks through major media outlets during the high seasons, and physicians providing personalised information for immune compromised patients to help them prevent exposure to all Legionella species.
A more proactive approach in Australia and New Zealand may be to engage with the potting soil industry to develop low-risk manufactured potting soils. The apparent differences in epidemiology of Legionnaires’ disease between Europe and Australia and New Zealand suggests that there is less exposure to L. longbeachae in Europe. Pasteurising potting soils, or reducing or eliminating pine products from potting soils, and using peat and well composted green waste, which appear to be inhospitable to L. longbeachae, may reduce these infections. As peat is a limited resource, supply may become restricted for environmental reasons so that manufactures may turn to other products such as pine bark to fill the gap.
The prevention of Legionnaires’ disease in hospitals has been widely discussed, and water standards have been set by the World Health Organization and many national agencies [137,138]. These have been developed appropriately with particular reference to L. pneumophila, but the measures employed to minimise infection with this organism would be expected to be effective for other waterborne Legionella species. These include well designed hospital water systems, and maintenance of hot water systems, to ensure inhibitory temperatures are maintained. Hot water pipes need to be properly insulated to prevent the warming of adjacent cold-water pipes, stagnation sites such as blind pipe sections and holding tanks eliminated, and material that supports legionella growth minimised [139,140,141,142]. A metanalysis has shown that temperatures of 55–59°C are associated with a low probability of Legionella species being detectable in water samples, and in cold water kept below 20 °C [143,144].
There is a lack of consensus on the need for monitoring water systems in hospitals, with some organisations recommending against monitoring, others monitoring only those areas housing immune-compromised patients, and others all hospital water supplies [145,146]. This is in part because it has been difficult to define specific and sensitive target concentrations for remediation although recent studies have suggested methods to improve monitoring techniques [100,147,148].
If Legionella species are detected, methods such as the use of monochloramine, heat, ultraviolet light, copper-silver ionization and chlorination have been used to reduce the risk to patients [67,149,150,151,152,153]. Improvement of chlorination of water supplies may reduce the burden of travel-related legionellosis from non-L pneumophila species [151]. It is important that detection systems are not solely focused on L. pneumophila, as molecular techniques may help identify the source of infection. These techniques should also be used for monitoring water distribution systems that may harbour non-L. pneumophila species [154].
6. Conclusions
The number and distribution of infections caused by L. longbeachae and other non-L. pneumophila legionellae are almost certainly markedly underestimated globally. Humans are accidental hosts, where infections are most likely to occur following interactions with the natural environment. It is the relationship between the environmental conditions and the interactions Legionella has with is protozoan hosts that shape their pathogenesis, enabling their survival and growth. Favourable conditions are needed to allow Legionella numbers to increase in the environment, thereby accounting for the seasonal pattern of human infections.
Legionnaires’ disease is a preventable pneumonia. Resourcing of the public health response will inevitably be governed by the number of cases identified and how political figures assign responsibility for controlling Legionnaires’ disease. Multiple factors currently combine to mitigate against case ascertainment. These include the passive design of surveillance systems with testing of only respiratory samples submitted for analysis. Many patients with community-acquired pneumonia are not asked to produce sputum samples, and up to 40% of patients with community-acquired pneumonia are unable to expectorate sputum when asked [62]. Investigation of nonsevere cases of community-acquired pneumonia is discouraged including for hospitalised cases in many centres. This is exacerbated by a perception among some clinicians that Legionnaires’ disease is necessarily a severe infection, and only these cases are investigated, and then testing is limited to only the urinary antigen test for L. pneumophila serogroup 1. Compounding these issues, investigations may only be done after antimicrobial therapy has been initiated, including agents that are active against Legionella species, thereby reducing the sensitivity of any testing that is conducted. There may be significant numbers of cases who present to health practitioners in the community and are not investigated or are not sufficiently unwell to seek medical attention. Lastly, financial constraints discourage testing, or at least minimise the resources allocated to investigating community-acquired pneumonia.
Practice guidelines make pragmatic sense but should not override the need for well conducted studies that adequately define the causes of community-acquired pneumonia in both the general public and immune compromised patients. These studies should avoid the tyranny of urine testing only, to identify pinch points where public health interventions can minimise infection rates of non-L. pneumophila species.
Author Contributions
Conceptualization, S.T.C., S.S., A.S.-T. and D.R.M.; Methodology, S.S.; Project administration, A.S.-T.; Supervision, S.T.C.; Validation, D.R.M.; Writing—original draft, S.T.C.; Writing—review & editing, S.S., A.S.-T. and D.R.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
No new data presented.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McDade, J.E.; Shepard, C.C.; Fraser, D.W.; Tsai, T.R.; Redus, M.A.; Dowdle, W.R. Legionnaires’ disease: Isolation of a bacterium and demonstration of its role in other respiratory disease. N. Engl. J. Med. 1977, 297, 1197–1203. [Google Scholar] [CrossRef] [PubMed]
- Honigsbaum, M. Legionnaires’ disease: Revisiting the puzzle of the century. Lancet 2016, 30, 456–457. [Google Scholar] [CrossRef]
- Cunha, B.; Burillo, A.; Bouza, E. Legionnaires’ disease. Lancet 2016, 23, 376–385. [Google Scholar] [CrossRef]
- Specials Pathogens Laboratory. Available online: https://specialpathogenslab.com/legionella-species/ (accessed on 21 January 2021).
- McKinney, R.M.; Porschen, R.K.; Edelstein, P.H.; Bissett, M.L.; Harris, P.P.; Bondell, S.P.; Steigerwalt, A.G.; Weaver, R.E.; Ein, M.E.; Lindquist, D.S.; et al. Legionella longbeachae species nova, another etiologic agent of human pneumonia. Ann. Intern. Med. 1981, 94, 739–743. [Google Scholar] [CrossRef]
- Australia’s notifiable disease status, 2015: Annual report of the National Notifiable Diseases Surveillance System. Commun. Dis. Intell. 2019, 43. [CrossRef]
- Priest, P.C.; Slow, S.; Chambers, S.T.; Cameron, C.M.; Balm, M.N.; Beale, M.W.; Blackmore, T.K.; Burns, A.D.; Drinković, D.; Elvy, J.A.; et al. The burden of Legionnaires’ disease in New Zealand (LegiNZ): A national surveillance study. Lancet Infect. Dis. 2019, 19, 770–777. [Google Scholar] [CrossRef]
- Boilattabi, N.; Barrassi, L.; Bouanane-Darenfed, A.; La Scola, B. Isolation and identification of Legionella spp. from hot spring water in Algeria by culture and molecular methods. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef]
- Steele, T.W. Legionnaires’ disease in South Australia, 1979–1988. Med. J. Aust. 1989, 151, 5–6. [Google Scholar] [CrossRef]
- Wright, A.J.; Humar, A.; Gourishankar, S.; Bernard, K.; Kumar, D. Severe Legionnaire’s disease caused by Legionella longbeachae in a long-term renal transplant patient: The importance of safe living strategies after transplantation. Transpl. Infect. Dis. 2012, 14, E30–E33. [Google Scholar] [CrossRef]
- Wang, J.Y.; Li, X.; Chen, J.Y.; Tong, B. Epileptic Seizure after Use of Moxifloxacin in Man with Legionella longbeachae Pneumonia. Emerg. Infect. Dis. 2020, 26, 2725–2727. [Google Scholar] [CrossRef]
- Svendsen, J.H.; Jønsson, V.; Niebuhr, U. Combined pericarditis and pneumonia caused by Legionella infection. Br. Heart J. 1987, 58, 663–664. [Google Scholar] [CrossRef] [PubMed]
- Mentula, S.; Pentikäinen, J.; Perola, O.; Ruotsalainen, E. Legionella longbeachae infection in a persistent hand-wound after a gardening accident. JMM Case Rep. 2014, 1, e004374. [Google Scholar] [CrossRef] [PubMed]
- Desplaces, N.; Nahapetian, N.; Bure, A.; Dournon, E. Legionella longbeachae serogroup 2 in Europe. Lancet 1982, 2, 1400. [Google Scholar] [CrossRef]
- Kümpers, P.; Tiede, A.; Kirschner, P.; Girke, J.; Ganser, A.; Peest, D. Legionnaires’ disease in immunocompromised patients: A case report of Legionella longbeachae pneumonia and review of the literature. J. Med. Microbiol. 2008, 57, 384–387. [Google Scholar] [CrossRef] [PubMed]
- Velonakis, E.N.; Kiousi, I.M.; Koutis, C.; Papadogiannakis, E.; Babatsikou, F.; Vatopoulos, A. First isolation of Legionella species, including L. pneumophila serogroup 1, in Greek potting soils: Possible importance for public health. Clin. Microbiol. Infect. 2010, 16, 763–766. [Google Scholar] [CrossRef] [PubMed]
- Lang, R.; Wiler, Z.; Manor, J.; Kazak, R.; Boldur, I. Legionella longbeachae pneumonia in a patient splenectomized for hairy-cell leukemia. Infection 1990, 18, 31–32. [Google Scholar] [CrossRef]
- Irie, S.; Akagi, K.; Hiraga, T.; Tada, S.; Kimura, I.; Saito, A. The first case of Legionella longbeachae pneumonia in Japan. Nihon Kyobu Shikkan Gakkai Zasshi 1984, 22, 518–522. [Google Scholar]
- van’t Hullenaar, N.G.; van Ketel, R.J.; Kuijper, E.J.; Bakker, P.J.; Dankert, J. Relapse of Legionella longbeachae infection in an immunocompromised patient. Neth. J. Med. 1996, 49, 202–204. [Google Scholar] [CrossRef]
- Yu, V.L.; Plouffe, J.F.; Pastoris, M.C.; Stout, J.E.; Schousboe, M.; Widmer, A.; Summersgill, J.; File, T.; Heath, C.M.; Paterson, D.L.; et al. Distribution of Legionella species and serogroups isolated by culture in patients with sporadic community-acquired legionellosis: An international collaborative survey. J. Infect. Dis. 2002, 186, 127–128. [Google Scholar] [CrossRef]
- Wolter, N.; Carrim, M.; Cohen, C.; Tempia, S.; Walaza, S.; Sahr, P.; de Gouveia, L.; Treurnicht, F.; Hellferscee, O.; Cohen, A.L.; et al. Legionnaires’ Disease in South Africa, 2012–2014. Emerg. Infect. Dis. 2016, 22, 131–133. [Google Scholar] [CrossRef]
- Lee, H.K.; Woo, M.K.; Ju, Y.I.; Baek, S.J.; Song, H.J.; Choi, J.S.; Kweon, S.S.; Jeon, D.Y.; Kang, Y.H. Prevalence of antibodies in response to Legionella species, analysis of a healthy population from Jeollanam-do Province, Korea. J. Microbiol. 2008, 46, 160–164. [Google Scholar] [CrossRef] [PubMed]
- García, C.; Ugalde, E.; Campo, A.B.; Miñambres, E.; Kovács, N. Fatal case of community-acquired pneumonia caused by Legionella longbeachae in a patient with systemic lupus erythematosus. Eur. J. Clin. Microbiol. Infect. Dis. 2004, 23, 116–118. [Google Scholar] [CrossRef] [PubMed]
- Eitrem, R.; Forsgren, A.; Nilsson, C. Pneumonia and acute pancreatitis most probably caused by a Legionella longbeachae infection. Scand. J. Infect. Dis. 1987, 19, 381–382. [Google Scholar] [CrossRef] [PubMed]
- Reynaldos Canton Migotto, T.; Györik Lora, S.; Gaia, V.; Pagnamenta, A. ARDS with septic shock due to Legionella longbeachae pneumonia in a patient with polymyalgia rheumatica. HeartLung Vessel. 2014, 6, 114–118. [Google Scholar]
- Wei, S.H.; Tseng, L.R.; Tan, J.K.; Cheng, C.Y.; Hsu, Y.T.; Cheng, E.T.; Lu, C.S.; Hsiao, Y.C.; Wu, T.H.; Hsu, J.F.; et al. Legionnaires’ disease caused by Legionella longbeachae in Taiwan, 2006–2010. Int. J. Infect. Dis. 2014, 19, 95–97. [Google Scholar] [CrossRef][Green Version]
- Phares, C.R.; Wangroongsarb, P.; Chantra, S.; Paveenkitiporn, W.; Tondella, M.L.; Benson, R.F.; Thacker, W.L.; Fields, B.S.; Moore, M.R.; Fischer, J.; et al. Epidemiology of severe pneumonia caused by Legionella longbeachae, Mycoplasma pneumoniae, and Chlamydia pneumoniae: 1-year, population-based surveillance for severe pneumonia in Thailand. Clin. Infect. Dis. 2007, 45, e147–e155. [Google Scholar] [CrossRef]
- Pravinkumar, S.J.; Edwards, G.; Lindsay, D.; Redmond, S.; Stirling, J.; House, R.; Kerr, J.; Anderson, E.; Breen, D.; Blatchford, O.; et al. A cluster of Legionnaires’ disease caused by Legionella longbeachae linked to potting compost in Scotland, 2008–2009. Eurosurveillance 2010, 15, 19496. [Google Scholar] [CrossRef]
- The Pneumonia Etiology Research for Child Health (PERCH) Study Group. Causes of severe pneumonia requiring hospital admission in children without HIV infection from Africa and Asia: The PERCH multi-country case-control study. Lancet 2019, 394, 757–779. [Google Scholar] [CrossRef]
- Puri, S.; Boudreaux-Kelly, M.; Walker, J.D.; Clancy, C.J.; Decker, B.K. Clinical Presentation of Community-Acquired Legionella Pneumonia Identified by Universal Testing in an Endemic Area. Int. J. Envion. Res. Public Health 2020, 17, 533. [Google Scholar] [CrossRef]
- Isenman, H.; Chambers, S.; Pithie, A.; MacDonald, S.; Hegarty, J.; Fenwick, J.; Maze, M.; Metcalf, S.; Murdoch, D. Legionnaires’ disease caused by Legionella longbeachae: Clinical features and outcomes of 107 cases from an endemic area. Respirology 2016, 21, 1292–1299. [Google Scholar] [CrossRef]
- Ito, A.; Ishida, T.; Washio, Y.; Yamazaki, A.; Tachibana, H. Legionella pneumonia due to non-Legionella pneumophila serogroup 1: Usefulness of the six-point scoring system. BMC Pulm. Med. 2017, 17, 211. [Google Scholar] [CrossRef] [PubMed]
- Glick, T.H.; Gregg, M.B.; Berman, B.; Mallison, G.; Rhodes, W.W., Jr.; Kassanoff, I. Pontiac fever. An epidemic of unknown etiology in a health department: I. Clinical and epidemiologic aspects. Am. J. Epidemiol. 1978, 107, 149–160. [Google Scholar] [CrossRef] [PubMed]
- Herwaldt, L.A.; Gorman, G.W.; McGrath, T.; Toma, S.; Brake, B.; Hightower, A.W.; Jones, J.; Reingold, A.L.; Boxer, P.A.; Tang, P.W.; et al. A new Legionella species, Legionella feeleii species nova, causes Pontiac fever in an automobile plant. Ann. Intern. Med. 1984, 100, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Fallon, R.J.; Rowbotham, T.J. Microbiological investigations into an outbreak of Pontiac fever due to Legionella micdadei associated with use of a whirlpool. J. Clin. Pathol. 1990, 43, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Götz, H.M.; Tegnell, A.; De Jong, B.; Broholm, K.A.; Kuusi, M.; Kallings, I.; Ekdahl, K. A whirlpool associated outbreak of Pontiac fever at a hotel in Northern Sweden. Epidemiol. Infect. 2001, 126, 241–247. [Google Scholar] [CrossRef]
- Cramp, G.J.; Harte, D.; Douglas, N.M.; Graham, F.; Schousboe, M.; Sykes, K. An outbreak of Pontiac fever due to Legionella longbeachae serogroup 2 found in potting mix in a horticultural nursery in New Zealand. Epidemiol. Infect. 2010, 138, 15–20. [Google Scholar] [CrossRef]
- Fenstersheib, M.D.; Miller, M.; Diggins, C.; Liska, S.; Detwiler, L.; Werner, S.B.; Lindquist, D.; Thacker, W.L.; Benson, R.F. Outbreak of Pontiac fever due to Legionella anisa. Lancet 1990, 336, 35–37. [Google Scholar] [CrossRef]
- Jones, T.F.; Benson, R.F.; Brown, E.W.; Rowland, J.R.; Crosier, S.C.; Schaffner, W. Epidemiologic investigation of a restaurant-associated outbreak of Pontiac fever. Clin. Infect. Dis. 2003, 37, 1292–1297. [Google Scholar] [CrossRef]
- Fields, B.S.; Haupt, T.; Davis, J.P.; Arduino, M.J.; Miller, P.H.; Butler, J.C. Pontiac fever due to Legionella micdadei from a whirlpool spa: possible role of bacterial endotoxin. J. Infect. Dis. 2001, 184, 1289–1292. [Google Scholar] [CrossRef]
- Desgranges, F.; Coste, A.T.; Wernly, D.; Dufour, J.; Opota, O.; Meylan, S. Immunosuppressed gardener pricked by roses grows Legionella longbeachae. Lancet 2020, 395, 604. [Google Scholar] [CrossRef]
- Vaidya, T.; Schmidt, E.; Papanicolaou, G.; Hauser, J.; Lezcano, C.; Tang, Y.W.; Markova, A. Cutaneous Legionella infections in allogeneic hematopoietic cell transplantation recipients. Derm. Online J. 2020, 26, 4. [Google Scholar]
- Ibranosyan, M.; Beraud, L.; Lemaire, H.; Ranc, A.G.; Ginevra, C.; Jarraud, S.; Descours, G. The clinical presentation of Legionella arthritis reveals the mode of infection and the bacterial species: Case report and literature review. BMC Infect. Dis. 2019, 19, 864. [Google Scholar] [CrossRef] [PubMed]
- Banderet, F.; Blaich, A.; Soleman, E.; Gaia, V.; Osthoff, M. Septic arthritis due to Legionella cincinnatiensis: Case report and review of the literature. Infection 2017, 45, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Just, S.A.; Knudsen, J.B.; Uldum, S.A.; Holt, H.M. Detection of Legionella bozemanae, a new cause of septic arthritis, by PCR followed by specific culture. J. Clin. Microbiol. 2012, 50, 4180–4182. [Google Scholar] [CrossRef]
- Compain, F.; Bruneval, P.; Jarraud, S.; Perrot, S.; Aubert, S.; Napoly, V.; Ramahefasolo, A.; Mainardi, J.L.; Podglajen, I. Chronic endocarditis due to Legionella anisa: A first case difficult to diagnose. Infection 2015, 8, 113–115. [Google Scholar] [CrossRef] [PubMed]
- Leggieri, N.; Gouriet, F.; Thuny, F.; Habib, G.; Raoult, D.; Casalta, J.P. Legionella longbeachae and endocarditis. Emerg. Infect. Dis. 2012, 18, 95–97. [Google Scholar] [CrossRef]
- Fukuta, Y.; Yildiz-Aktas, I.Z.; William Pasculle, A.; Veldkamp, P.J. Legionella micdadei prosthetic valve endocarditis complicated by brain abscess: Case report and review of the literature. Scand. J. Infect. Dis. 2012, 44, 414–418. [Google Scholar] [CrossRef]
- Pearce, M.M.; Theodoropoulos, N.; Noskin, G.A.; Flaherty, J.P.; Stemper, M.E.; Aspeslet, T.; Cianciotto, N.P.; Reed, K.D. Native valve endocarditis due to a novel strain of Legionella. J. Clin. Microbiol. 2011, 49, 3340–3342. [Google Scholar] [CrossRef]
- Heriot, W.J.; Mack, H.G.; Stawell, R. Ocular involvement in a patient with Legionella longbeachae 1 infection. Clin. Exp. Ophthalmol. 2014, 42, 497–499. [Google Scholar] [CrossRef]
- Neiderud, C.J.; Vidh, A.L.; Salaneck, E. Soft tissue infection caused by Legionella bozemanii in a patient with ongoing immunosuppressive treatment. Infect. Ecol. Epidemiol. 2013, 3. [Google Scholar] [CrossRef]
- Metlay, J.P.; Waterer, G.W.; Long, A.C.; Anzueto, A.; Brozek, J.; Crothers, K.; Cooley, L.A.; Dean, N.C.; Fine, M.J.; Flanders, S.A.; et al. Diagnosis and Treatment of Adults with Community-acquired Pneumonia. An Official Clinical Practice Guideline of the American Thoracic Society and Infectious Diseases Society of America. Am. J. Respir. Crit. Care Med. 2019, 200, e45–e67. [Google Scholar] [CrossRef] [PubMed]
- Murdoch, D.R.; Podmore, R.G.; Anderson, T.P.; Barratt, K.; Maze, M.J.; French, K.E.; Young, S.A.; Chambers, S.T.; Werno, A.M. Impact of routine systematic polymerase chain reaction testing on case finding for Legionnaires’ disease: A pre-post comparison study. Clin. Infect. Dis. 2013, 57, 1275–1281. [Google Scholar] [CrossRef] [PubMed]
- Han, X.Y.; Ihegword, A.; Evans, S.E.; Zhang, J.; Li, L.; Cao, H.; Tarrand, J.J.; El-Kweifi, O. Microbiological and Clinical Studies of Legionellosis in 33 Patients with Cancer. J. Clin. Microbiol. 2015, 53, 2180–2187. [Google Scholar] [CrossRef] [PubMed]
- ESR. Annual Notifiable Disease Tables. Available online: https://surv.esr.cri.nz/surveillance/annual_diseasetables.php (accessed on 10 December 2020).
- Miyashita, N.; Higa, F.; Aoki, Y.; Kikuchi, T.; Seki, M.; Tateda, K.; Maki, N.; Uchino, K.; Ogasawara, K.; Kiyota, H.; et al. Distribution of Legionella species and serogroups in patients with culture-confirmed Legionella pneumonia. J. Infect. Chemother. 2020, 26, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Benin, A.L.; Benson, R.F.; Besser, R.E. Trends in legionnaires disease, 1980–1998: Declining mortality and new patterns of diagnosis. Clin. Infect. Dis. 2002, 35, 1039–1046. [Google Scholar] [CrossRef]
- Beauté, J.; Plachouras, D.; Sandin, S.; Giesecke, J.; Sparén, P. Healthcare-Associated Legionnaires’ Disease, Europe, 2008–2017. Emerg. Infect. Dis. 2020, 26, 2309–2318. [Google Scholar] [CrossRef]
- Potts, A.; Donaghy, M.; Marley, M.; Othieno, R.; Stevenson, J.; Hyland, J.; Pollock, K.G.; Lindsay, D.; Edwards, G.; Hanson, M.F.; et al. Cluster of Legionnaires disease cases caused by Legionella longbeachae serogroup 1, Scotland, August to September 2013. Eurosurveillance 2013, 18, 20656. [Google Scholar] [CrossRef]
- Löf, E. Fellowship Report. Summary of Work Activities. 2019. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/EPIET%20report%20-%20Summary%20of%20work%20activities%2C%20Emma%20L%C3%B6f.pdf (accessed on 31 December 2020).
- Legionella (Legionnaires’ Disease and Pontiac Fever). Available online: https://www.cdc.gov/legionella/health-depts/surv-reporting/index.html (accessed on 31 December 2020).
- Maze, M.J.; Slow, S.; Cumins, A.M.; Boon, K.; Goulter, P.; Podmore, R.G.; Anderson, T.P.; Barratt, K.; Young, S.A.; Pithie, A.D.; et al. Enhanced detection of Legionnaires’ disease by PCR testing of induced sputum and throat swabs. Eur. Respir. J. 2014, 43, 644–646. [Google Scholar] [CrossRef]
- Cameron, R.L.; Pollock, K.G.J.; Lindsay, D.S.J.; Anderson, E. Comparison of Legionella longbeachae and Legionella pneumophila cases in Scotland; implications for diagnosis, treatment and public health response. J. Med. Microbiol. 2016, 65, 142–146. [Google Scholar] [CrossRef]
- de Bruin, L.; Timmerman, C.P.; Huisman, P.M.; Heidt, J. Legionella longbeachae; don’t miss it! Neth. J. Med. 2018, 76, 294–297. [Google Scholar]
- Grove, D.I.; Lawson, P.J.; Burgess, J.S.; Moran, J.L.; O’Fathartaigh, M.S.; Winslow, W.E. An outbreak of Legionella longbeachae infection in an intensive care unit? J. Hosp. Infect. 2002, 52, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Doebbeling, B.N.; Ishak, M.A.; Wade, B.H.; Pasquale, M.A.; Gerszten, R.E.; Gröschel, D.H.; Kadner, R.J.; Wenzel, R.P. Nosocomial Legionella micdadei pneumonia: 10 years experience and a case-control study. J. Hosp. Infect. 1989, 13, 289–298. [Google Scholar] [CrossRef]
- Parry, M.F.; Stampleman, L.; Hutchinson, J.H.; Folta, D.; Steinberg, M.G.; Krasnogor, L.J. Waterborne Legionella bozemanii and nosocomial pneumonia in immunosuppressed patients. Ann. Intern. Med. 1985, 103, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Wolff, C.; Lange, H.; Feruglio, S.; Vold, L.; MacDonald, E. Evaluation of the national surveillance of Legionnaires’ disease in Norway, 2008-2017. BMC Public Health 2019, 19, 1624. [Google Scholar] [CrossRef] [PubMed]
- Dabrera, G.; Brandsema, P.; Lofdahl, M.; Naik, F.; Cameron, R.; McMenamin, J.; Pebody, R.; Phin, N. Increase in Legionnaires’ disease cases associated with travel to Dubai among travellers from the United Kingdom, Sweden and the Netherlands, October 2016 to end August 2017. Eurosurveillance 2017, 22. [Google Scholar] [CrossRef] [PubMed]
- Steele, T.W.; Lanser, J.; Sangster, N. Isolation of Legionella longbeachae serogroup 1 from potting mixes. Appl. Environ. Microbiol. 1990, 56, 49–53. [Google Scholar] [CrossRef]
- Cameron, S.; Roder, D.; Walker, C.; Feldheim, J. Epidemiological characteristics of Legionella infection in South Australia: Implications for disease control. Aust. N. Z. J. Med. 1991, 21, 65–70. [Google Scholar] [CrossRef]
- O’Connor, B.A.; Carman, J.; Eckert, K.; Tucker, G.; Givney, R.; Cameron, S. Does using potting mix make you sick? Results from a Legionella longbeachae case-control study in South Australia. Epidemiol. Infect. 2007, 135, 34–39. [Google Scholar] [CrossRef]
- Currie, S.L.; Beattie, T.K.; Knapp, C.W.; Lindsay, D.S. Legionella spp. in UK composts—A potential public health issue? Clin. Microbiol. Infect. 2014, 20, O224–O229. [Google Scholar] [CrossRef]
- Li, J.S.; O’Brien, E.D.; Guest, C. A review of national legionellosis surveillance in Australia, 1991 to 2000. Commun. Dis. Intell. Q Rep. 2002, 26, 461–468. [Google Scholar]
- Kubota, M.; Tomii, K.; Tachikawa, R.; Harada, Y.; Seo, R.; Kaji, R.; Takeshima, Y.; Hayashi, M.; Nishimura, T.; Ishihara, K. Legionella longbeachae pneumonia infection from home garden soil. Nihon Kokyuki Gakkai Zasshi 2007, 45, 698–703. [Google Scholar] [PubMed]
- Legionnaires’ Disease associated with potting soil--California, Oregon, and Washington, May–June 2000. MMWR Morb. Mortal. Wkly Rep. 2000, 49, 777–778.
- Kenagy, E.; Priest, P.C.; Cameron, C.M.; Smith, D.; Scott, P.; Cho, V.; Mitchell, P.; Murdoch, D.R. Risk Factors for Legionella longbeachae Legionnaires’ Disease, New Zealand. Emerg. Infect. Dis. 2017, 23, 1148–1154. [Google Scholar] [CrossRef] [PubMed]
- Steele, T.W.; Moore, C.V.; Sangster, N. Distribution of Legionella longbeachae serogroup 1 and other legionellae in potting soils in Australia. Appl. Environ. Microbiol. 1990, 56, 2984–2988. [Google Scholar] [CrossRef] [PubMed]
- Casati, S.; Gioria-Martinoni, A.; Gaia, V. Commercial potting soils as an alternative infection source of Legionella pneumophila and other Legionella species in Switzerland. Clin Microbiol. Infect. 2009, 15, 571–575. [Google Scholar] [CrossRef]
- Huss, A.; Derks, L.A.N.; Heederik, D.J.J.; Wouters, I.M. Green waste compost as potential reservoirs of Legionella in the Netherlands. Clin. Microbiol. Infect. 2020, 26, 1259.e1–1259.e3. [Google Scholar] [CrossRef] [PubMed]
- Koide, M.; Saito, A.; Okazaki, M.; Umeda, B.; Benson, R.F. Isolation of Legionella longbeachae serogroup 1 from potting soils in Japan. Clin. Infect. Dis. 1999, 29, 943–944. [Google Scholar] [CrossRef][Green Version]
- Koide, M.; Arakaki, N.; Saito, A. Distribution of Legionella longbeachae and other legionellae in Japanese potting soils. J. Infect. Chemother. 2001, 7, 224–227. [Google Scholar] [CrossRef]
- Lindsay, D.S.J.; Brown, A.W.; Brown, D.J.; Pravinkumar, S.J.; Anderson, E.; Edwards, G.F.S. Legionella longbeachae serogroup 1 infections linked to potting compost. J. Med. Microbiol. 2012, 61, 218–222. [Google Scholar] [CrossRef]
- van Heijnsbergen, E.; van Deursen, A.; Bouwknegt, M.; Bruin, J.P.; de Roda Husman, A.M.; Schalk, J.A. Presence and Persistence of Viable, Clinically Relevant Legionella pneumophila Bacteria in Garden Soil in the Netherlands. Appl. Environ. Microbiol. 2016, 82, 5125–5131. [Google Scholar] [CrossRef]
- Bacigalupe, R.; Lindsay, D.; Edwards, G.; Fitzgerald, J.R. Population Genomics of Legionella longbeachae and Hidden Complexities of Infection Source Attribution. Emerg. Infect. Dis. 2017, 23, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.S.; Steele, T.W. Occurrence and distribution of Legionella species in composted plant materials. Appl. Environ. Microbiol. 1994, 60, 2003–2005. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Roque, C.M.; Hazen, T.C. Abundance and distribution of Legionellaceae in Puerto Rican waters. Appl. Environ. Microbiol. 1987, 53, 2231–2236. [Google Scholar] [CrossRef]
- van Heijnsbergen, E.; de Roda Husman, A.M.; Lodder, W.J.; Bouwknegt, M.; Docters van Leeuwen, A.E.; Bruin, J.P.; Euser, S.M.; den Boer, J.W.; Schalk, J.A. Viable Legionella pneumophila bacteria in natural soil and rainwater puddles. J. Appl. Microbiol. 2014, 117, 882–890. [Google Scholar] [CrossRef] [PubMed]
- Travis, T.C.; Brown, E.W.; Peruski, L.F.; Siludjai, D.; Jorakate, P.; Salika, P.; Yang, G.; Kozak, N.A.; Kodani, M.; Warner, A.K.; et al. Survey of Legionella species found in thai soil. Int. J. Microbiol. 2012, 2012, 218791. [Google Scholar] [CrossRef]
- Mohammadi, A. Development of Detection Methods for Legionella longbeachae in Clinical and Environmental Samples. Ph.D. Thesis, University of Otago, Christchurch, New Zealand, 2020. [Google Scholar]
- Chambers, S.; Slow, S.; Withers, A.; Chim, M.; Dawson, K.; Clemens, J.; Anderson, T.; Williman, J.; Murdoch, D.; Scott-Thomas, A. Pine Species Provide a Niche for Legionella longbeachae. J. Appl. Environ. Microbiol. 2020, 8, 46–52. [Google Scholar] [CrossRef]
- Loh, C.H.; Soni, R. Exposure to potting soils and compost material as potential sources of Legionella pneumophilia in Australia. Respir. Med. Case Rep. 2020, 31, 101156. [Google Scholar] [CrossRef]
- Conza, L.; Pagani, S.C.; Gaia, V. Presence of Legionella and free-living Amoebae in composts and bioaerosols from composting facilities. PLOS ONE 2013, 8, e68244. [Google Scholar] [CrossRef]
- Marrie, T.; Green, P.; Burbridge, S.; Bezanson, G.; Neale, S.; Hoffman, P.S.; Haldane, D. Legionellaceae in the potable water of Nova Scotia hospitals and Halifax residences. Epidemiol. Infect. 1994, 112, 143–150. [Google Scholar] [CrossRef]
- Dobrowsky, P.H.; Khan, S.; Cloete, T.E.; Khan, W. Molecular detection of Acanthamoeba spp., Naegleria fowleri and Vermamoeba (Hartmannella) vermiformis as vectors for Legionella spp. in untreated and solar pasteurized harvested rainwater. Parasit. Vectors 2016, 9, 539. [Google Scholar] [CrossRef]
- Thornley, C.N.; Harte, D.J.; Weir, R.P.; Allen, L.J.; Knightbridge, K.J.; Wood, P.R.T. Legionella longbeachae detected in an industrial cooling tower linked to a legionellosis outbreak, New Zealand, 2015; possible waterborne transmission? Epidemiol. Infect. 2017, 145, 2382–2389. [Google Scholar] [CrossRef] [PubMed]
- Orkis, L.T.; Harrison, L.H.; Mertz, K.J.; Brooks, M.M.; Bibby, K.J.; Stout, J.E. Environmental sources of community-acquired legionnaires’ disease: A review. Int. J. Hygen Environ. Health 2018, 221, 764–774. [Google Scholar] [CrossRef] [PubMed]
- Buse, H.Y.; Morris, B.J.; Gomez-Alvarez, V.; Szabo, J.G.; Hall, J.S. Legionella Diversity and Spatiotemporal Variation in The Occurrence of Opportunistic Pathogens within a Large Building Water System. Pathogen 2020, 9, 567. [Google Scholar] [CrossRef] [PubMed]
- Knirsch, C.A.; Jakob, K.; Schoonmaker, D.; Kiehlbauch, J.A.; Wong, S.J.; Della-Latta, P.; Whittier, S.; Layton, M.; Scully, B. An outbreak of Legionella micdadei pneumonia in transplant patients: Evaluation, molecular epidemiology, and control. Am. J. Med. 2000, 108, 290–295. [Google Scholar] [CrossRef]
- Girolamini, L.; Salaris, S.; Lizzadro, J.; Mazzotta, M.; Pascale, M.R.; Pellati, T.; Cristino, S. How Molecular Typing Can Support Legionella Environmental Surveillance in Hot Water Distribution Systems: A Hospital Experience. Int. J. Environ. Res. Public Health 2020, 17, 8662. [Google Scholar] [CrossRef]
- Leoni, E.; Zanetti, F.; Cristino, S.; Legnani, P.P. Monitoring and control of opportunistic bacteria in a spa water used for aerosol hydrotherapy. Ann. Ig 2005, 17, 377–384. [Google Scholar]
- Mondino, S.; Schmidt, S.; Rolando, M.; Escoll, P.; Gomez-Valero, L.; Buchrieser, C. Legionnaires’ Disease: State of the Art Knowledge of Pathogenesis Mechanisms of Legionella. Annu. Rev. Pathol. 2020, 15, 439–466. [Google Scholar] [CrossRef]
- Gomez-Valero, L.; Rusniok, C.; Carson, D.; Mondino, S.; Perez-Cobas, A.E.; Rolando, M.; Pasricha, S.; Reuter, S.; Demirtas, J.; Crumbach, J.; et al. More than 18,000 effectors in the Legionella genus genome provide multiple, independent combinations for replication in human cells. Proc. Natl. Acad. Sci. USA 2019, 116, 2265–2273. [Google Scholar] [CrossRef]
- Burstein, D.; Amaro, F.; Zusman, T.; Lifshitz, Z.; Cohen, O.; Gilbert, J.A.; Pupko, T.; Shuman, H.A.; Segal, G. Genomic analysis of 38 Legionella species identifies large and diverse effector repertoires. Nat. Genet. 2016, 48, 167–175. [Google Scholar] [CrossRef]
- Cazalet, C.; Gomez-Valero, L.; Rusniok, C.; Lomma, M.; Dervins-Ravault, D.; Newton, H.J.; Sansom, F.M.; Jarraud, S.; Zidane, N.; Ma, L.; et al. Analysis of the Legionella longbeachae genome and transcriptome uncovers unique strategies to cause Legionnaires’ disease. PLOS Genet. 2010, 6, e1000851. [Google Scholar] [CrossRef]
- Kozak, N.A.; Buss, M.; Lucas, C.E.; Frace, M.; Govil, D.; Travis, T.; Olsen-Rasmussen, M.; Benson, R.F.; Fields, B.S. Virulence factors encoded by Legionella longbeachae identified on the basis of the genome sequence analysis of clinical isolate D-4968. J. Bacteriol. 2010, 192, 1030–1044. [Google Scholar] [CrossRef] [PubMed]
- Slow, S.; Anderson, T.; Miller, J.; Singh, S.; Murdoch, D.; Biggs, P.J. Complete Genome Sequence of a Legionella longbeachae Serogroup 1 Strain Isolated from a Patient with Legionnaires’ Disease. Genome Announc. 2017, 5. [Google Scholar] [CrossRef] [PubMed]
- Haviernik, J.; Dawson, K.; Anderson, T.; Murdoch, D.; Chambers, S.; Biggs, P.; Cree, S.; Slow, S. Complete Genome Sequence of a Legionella longbeachae Serogroup 2 Isolate Derived from a Patient with Legionnaires’ Disease. Microbiol. Resour. Announc. 2020, 9. [Google Scholar] [CrossRef]
- Slow, S.; Anderson, T.; Murdoch, D.; Bloomfield, S.; Winter, D.; Biggs, P. Extensive epigenetic modification with large-scale chromosomal and plasmid recombination characterise the Legionella longbeachae serogroup 1 genome. bioRxiv 2020. [Google Scholar] [CrossRef]
- Gomez-Valero, L.; Rusniok, C.; Cazalet, C.; Buchrieser, C. Comparative and functional genomics of Legionella identified eukaryotic like proteins as key players in host-pathogen interactions. Front. Microbiol. 2011, 2, 208. [Google Scholar] [CrossRef] [PubMed]
- Edelstein, P.H.; Roy, C.R. Legionnaires’ Disease and Pontiac Fever. In Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 8th ed.; Bennett, J.E., Dolin, R., Blaser, M.J., Eds.; Elsevier: Philadelphia, PA, USA, 2015. [Google Scholar]
- Johnson, J.T.; Yu, V.L.; Best, M.G.; Vickers, R.M.; Goetz, A.; Wagner, R.; Wicker, H.; Woo, A. Nosocomial legionellosis in surgical patients with head-and-neck cancer: Implications for epidemiological reservoir and mode of transmission. Lancet 1985, 2, 298–300. [Google Scholar] [CrossRef]
- Berendt, R.F.; Young, H.W.; Allen, R.G.; Knutsen, G.L. Dose-response of guinea pigs experimentally infected with aerosols of Legionella pneumophila. J. Infect. Dis. 1980, 141, 186–192. [Google Scholar] [CrossRef]
- Sivagnanam, S.; Podczervinski, S.; Butler-Wu, S.M.; Hawkins, V.; Stednick, Z.; Helbert, L.A.; Glover, W.A.; Whimbey, E.; Duchin, J.; Cheng, G.S.; et al. Legionnaires’ disease in transplant recipients: A 15-year retrospective study in a tertiary referral center. Transpl. Infect. Dis. 2017, 19. [Google Scholar] [CrossRef]
- Marston, B.J.; Lipman, H.B.; Breiman, R.F. Surveillance for Legionnaires’ disease. Risk factors for morbidity and mortality. Arch. Intern. Med. 1994, 154, 2417–2422. [Google Scholar] [CrossRef]
- Lanternier, F.; Tubach, F.; Ravaud, P.; Salmon, D.; Dellamonica, P.; Bretagne, S.; Couret, M.; Bouvard, B.; Debandt, M.; Gueit, I.; et al. Incidence and risk factors of Legionella pneumophila pneumonia during anti-tumor necrosis factor therapy: A prospective French study. Chest 2013, 144, 990–998. [Google Scholar] [CrossRef]
- Wang, H.; D’Souza, C.; Lim, X.Y.; Kostenko, L.; Pediongco, T.J.; Eckle, S.B.G.; Meehan, B.S.; Shi, M.; Wang, N.; Li, S.; et al. MAIT cells protect against pulmonary Legionella longbeachae infection. Nat. Commun. 2018, 9, 3350. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, P.S.; Ripley, M.; Weeratna, R. Cloning and nucleotide sequence of a gene (ompS) encoding the major outer membrane protein of Legionella pneumophila. J. Bacteriol. 1992, 174, 914–920. [Google Scholar] [CrossRef] [PubMed]
- Payne, N.R.; Horwitz, M.A. Phagocytosis of Legionella pneumophila is mediated by human monocyte complement receptors. J. Exp. Med. 1987, 166, 1377–1389. [Google Scholar] [CrossRef]
- Stone, B.J.; Abu Kwaik, Y. Expression of multiple pili by Legionella pneumophila: Identification and characterization of a type IV pilin gene and its role in adherence to mammalian and protozoan cells. Infect. Immun. 1998, 66, 1768–1775. [Google Scholar] [CrossRef] [PubMed]
- Cianciotto, N.P.; Stamos, J.K.; Kamp, D.W. Infectivity of Legionella pneumophila mip mutant for alveolar epithelial cells. Curr. Microbiol. 1995, 30, 247–250. [Google Scholar] [CrossRef]
- Cianciotto, N.P.; Bangsborg, J.M.; Eisenstein, B.I.; Engleberg, N.C. Identification of mip-like genes in the genus Legionella. Infect. Immun. 1990, 58, 2912–2918. [Google Scholar] [CrossRef]
- Mody, C.H.; Paine, R., 3rd; Shahrabadi, M.S.; Simon, R.H.; Pearlman, E.; Eisenstein, B.I.; Toews, G.B. Legionella pneumophila replicates within rat alveolar epithelial cells. J. Infect. Dis. 1993, 167, 1138–1145. [Google Scholar] [CrossRef]
- Massis, L.M.; Assis-Marques, M.A.; Castanheira, F.V.; Capobianco, Y.J.; Balestra, A.C.; Escoll, P.; Wood, R.E.; Manin, G.Z.; Correa, V.M.; Alves-Filho, J.C.; et al. Legionella longbeachae Is Immunologically Silent and Highly Virulent In Vivo. J. Infect. Dis. 2017, 215, 440–451. [Google Scholar] [CrossRef]
- Wood, R.E.; Newton, P.; Latomanski, E.A.; Newton, H.J. Dot/Icm Effector Translocation by Legionella longbeachae Creates a Replicative Vacuole Similar to That of Legionella pneumophila despite Translocation of Distinct Effector Repertoires. Infect. Immun. 2015, 83, 4081–4092. [Google Scholar] [CrossRef]
- Qin, T.; Zhou, H.; Ren, H.; Liu, W. Distribution of Secretion Systems in the Genus Legionella and Its Correlation with Pathogenicity. Front. Microbiol. 2017, 8, 388. [Google Scholar] [CrossRef][Green Version]
- Gomez-Valero, L.; Rusniok, C.; Rolando, M.; Neou, M.; Dervins-Ravault, D.; Demirtas, J.; Rouy, Z.; Moore, R.; Chen, H.; Petty, N.; et al. Comparative analyses of Legionella species identifies genetic features of strains causing Legionnaires’ disease. Genome Bio. 2014, 15, 505. [Google Scholar]
- Feldman, M.; Segal, G. A specific genomic location within the icm/dot pathogenesis region of different Legionella species encodes functionally similar but nonhomologous virulence proteins. Infect. Immun. 2004, 72, 4503–4511. [Google Scholar] [CrossRef] [PubMed]
- Boamah, D.K.; Zhou, G.; Ensminger, A.W.; O’Connor, T.J. From Many Hosts, One Accidental Pathogen: The Diverse Protozoan Hosts of Legionella. Front. Cell Infect. Microbiol. 2017, 7, 477. [Google Scholar] [CrossRef] [PubMed]
- Amaro, F.; Wang, W.; Gilbert, J.A.; Anderson, O.R.; Shuman, H.A. Diverse protist grazers select for virulence-related traits in Legionella. ISME J. 2015, 9, 1607–1618. [Google Scholar] [CrossRef] [PubMed]
- Hilbi, H.; Weber, S.S.; Ragaz, C.; Nyfeler, Y.; Urwyler, S. Environmental predators as models for bacterial pathogenesis. Environ. Microbiol. 2007, 9, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Swart, A.L.; Harrison, C.F.; Eichinger, L.; Steinert, M.; Hilbi, H. Acanthamoeba and Dictyostelium as Cellular Models for Legionella Infection. Front. Cell. Infect. Microbiol. 2018, 8. [Google Scholar] [CrossRef]
- Houghton, C.; Meskell, P.; Delaney, H.; Smalle, M.; Glenton, C.; Booth, A.; Chan, X.H.S.; Devane, D.; Biesty, L.M. Barriers and facilitators to healthcare workers’ adherence with infection prevention and control (IPC) guidelines for respiratory infectious diseases: A rapid qualitative evidence synthesis. Cochrane Database Syst. Rev. 2020, 4, Cd013582. [Google Scholar] [CrossRef]
- Matuschek, C.; Moll, F.; Fangerau, H.; Fischer, J.C.; Zänker, K.; van Griensven, M.; Schneider, M.; Kindgen-Milles, D.; Knoefel, W.T.; Lichtenberg, A.; et al. Face masks: Benefits and risks during the COVID-19 crisis. Eur. J. Med. Res. 2020, 25, 32. [Google Scholar] [CrossRef]
- Chu, D.K.; Akl, E.A.; Duda, S.; Solo, K.; Yaacoub, S.; Schünemann, H.J. Physical distancing, face masks, and eye protection to prevent person-to-person transmission of SARS-CoV-2 and COVID-19: A systematic review and meta-analysis. Lancet 2020, 395, 1973–1987. [Google Scholar] [CrossRef]
- Angen, Ø.; Skade, L.; Urth, T.R.; Andersson, M.; Bækbo, P.; Larsen, A.R. Controlling Transmission of MRSA to Humans During Short-Term Visits to Swine Farms Using Dust Masks. Front. Microbiol. 2018, 9, 3361. [Google Scholar] [CrossRef]
- WHO. WHO Guidelines Approved by the Guidelines Review Committee. In Guidelines for Drinking-Water Quality: Fourth Edition Incorporating the First Addendum; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- ESGLI. ESGLI European Technical Guidelines for the Prevention, Control and Investigation, of Infections Caused by Legionella species. In Diseases ESoCMaI; European Centre for Disease Prevention and Control (ECDC): Stockholm, Sweden, 2017; p. 125. [Google Scholar]
- National Academies of Sciences, Medicine, Health. Management of Legionella in Water Systems; National Academies Press: Washington, DC, USA, 2019. [Google Scholar]
- Cullom, A.C.; Martin, R.L.; Song, Y.; Williams, K.; Williams, A.; Pruden, A.; Edwards, M.A. Critical Review: Propensity of Premise Plumbing Pipe Materials to Enhance or Diminish Growth of Legionella and Other Opportunistic Pathogens. Pathogen 2020, 9, 957. [Google Scholar] [CrossRef] [PubMed]
- Hozalski, R.M.; LaPara, T.M.; Zhao, X.; Kim, T.; Waak, M.B.; Burch, T.; McCarty, M. Flushing of Stagnant Premise Water Systems after the COVID-19 Shutdown Can Reduce Infection Risk by Legionella and Mycobacterium spp. Environ. Sci. Technol. 2020, 54, 15914–15924. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.L.; Strom, O.R.; Pruden, A.; Edwards, M.A. Interactive Effects of Copper Pipe, Stagnation, Corrosion Control, and Disinfectant Residual Influenced Reduction of Legionella pneumophila during Simulations of the Flint Water Crisis. Pathogen 2020, 9, 730. [Google Scholar] [CrossRef]
- Rasheduzzaman, M.; Singh, R.; Haas, C.N.; Gurian, P.L. Required water temperature in hotel plumbing to control Legionella growth. Water Res. 2020, 182, 115943. [Google Scholar] [CrossRef] [PubMed]
- Bédard, E.; Fey, S.; Charron, D.; Lalancette, C.; Cantin, P.; Dolcé, P.; Laferrière, C.; Déziel, E.; Prévost, M. Temperature diagnostic to identify high risk areas and optimize Legionella pneumophila surveillance in hot water distribution systems. Water Res. 2015, 71, 244–256. [Google Scholar] [CrossRef]
- Veterans Health Administration. Prevention of Healthcare-Associated Legionella Disease and Scald Injury from Potable Water Distribution Systems; Department of Veteran Affairs: Washington, DC, USA, 2014; pp. 1–35.
- The Control of Legionella Bacteria in Water Systems, 4th ed.; Health and Safety Executive: London, UK, 2013.
- Hirsh, M.B.; Baron, J.L.; Mietzner, S.M.; Rihs, J.D.; Yassin, M.H.; Stout, J.E. Evaluation of Recommended Water Sample Collection Methods and the Impact of Holding Time on Legionella Recovery and Variability from Healthcare Building Water Systems. Microorganisms 2020, 8, 1770. [Google Scholar] [CrossRef] [PubMed]
- Nagy, D.J.; Dziewulski, D.M.; Codru, N.; Lauper, U.L. Understanding the distribution of positive Legionella samples in healthcare-premise water systems: Using statistical analysis to determine a distribution for Legionella and to support sample size recommendations. Infect. Control. Hosp. Epidemiol. 2021, 42, 63–68. [Google Scholar] [CrossRef]
- Duda, S.; Kandiah, S.; Stout, J.E.; Baron, J.L.; Yassin, M.; Fabrizio, M.; Ferrelli, J.; Hariri, R.; Wagener, M.M.; Goepfert, J.; et al. Evaluation of a new monochloramine generation system for controlling Legionella in building hot water systems. Infect. Control Hosp. Epidemiol. 2014, 35, 1356–1363. [Google Scholar] [CrossRef]
- Coniglio, M.A.; Ferrante, M.; Yassin, M.H. Preventing Healthcare-Associated Legionellosis: Results after 3 Years of Continuous Disinfection of Hot Water with Monochloramine and an Effective Water Safety Plan. Int. J. Environ. Res. Public Health 2018, 15, 1594. [Google Scholar] [CrossRef]
- Papadakis, A.; Chochlakis, D.; Sandalakis, V.; Keramarou, M.; Tselentis, Y.; Psaroulaki, A. Legionella spp. Risk Assessment in Recreational and Garden Areas of Hotels. Int. J. Environ. Res. Public Health 2018, 15, 598. [Google Scholar] [CrossRef]
- Muzzi, A.; Cutti, S.; Bonadeo, E.; Lodola, L.; Monzillo, V.; Corbella, M.; Scudeller, L.; Novelli, V.; Marena, C. Prevention of nosocomial legionellosis by best water management: Comparison of three decontamination methods. J. Hosp. Infect. 2020, 105, 766–772. [Google Scholar] [CrossRef] [PubMed]
- Marchesi, I.; Ferranti, G.; Mansi, A.; Marcelloni, A.M.; Proietto, A.R.; Saini, N.; Borella, P.; Bargellini, A. Control of Legionella Contamination and Risk of Corrosion in Hospital Water Networks following Various Disinfection Procedures. Appl. Environ. Microbiol. 2016, 82, 2959–2965. [Google Scholar] [CrossRef] [PubMed]
- Napoli, C.; De Giglio, O.; Bertamino, E.; Montagna, M.T. Legionellosis in health care facilities: State of the art in control and prevention in Italy. Ann. Ig 2019, 31, 474–481. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).