
The Use of Raman Spectroscopy to Monitor Metabolic Changes in Stressed Metschnikowia sp. Yeasts


 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Revitalization of Yeast Strains
2.3. Screening of C/N Ratio
2.4. Cell Preparation
2.5. Gas Chromatography Analysis
2.6. Raman Analysis
2.7. Raman Data Processing
2.8. Statistical Analysis
3. Results and Discussion
3.1. Monitoring of Production of Pulcherrimin
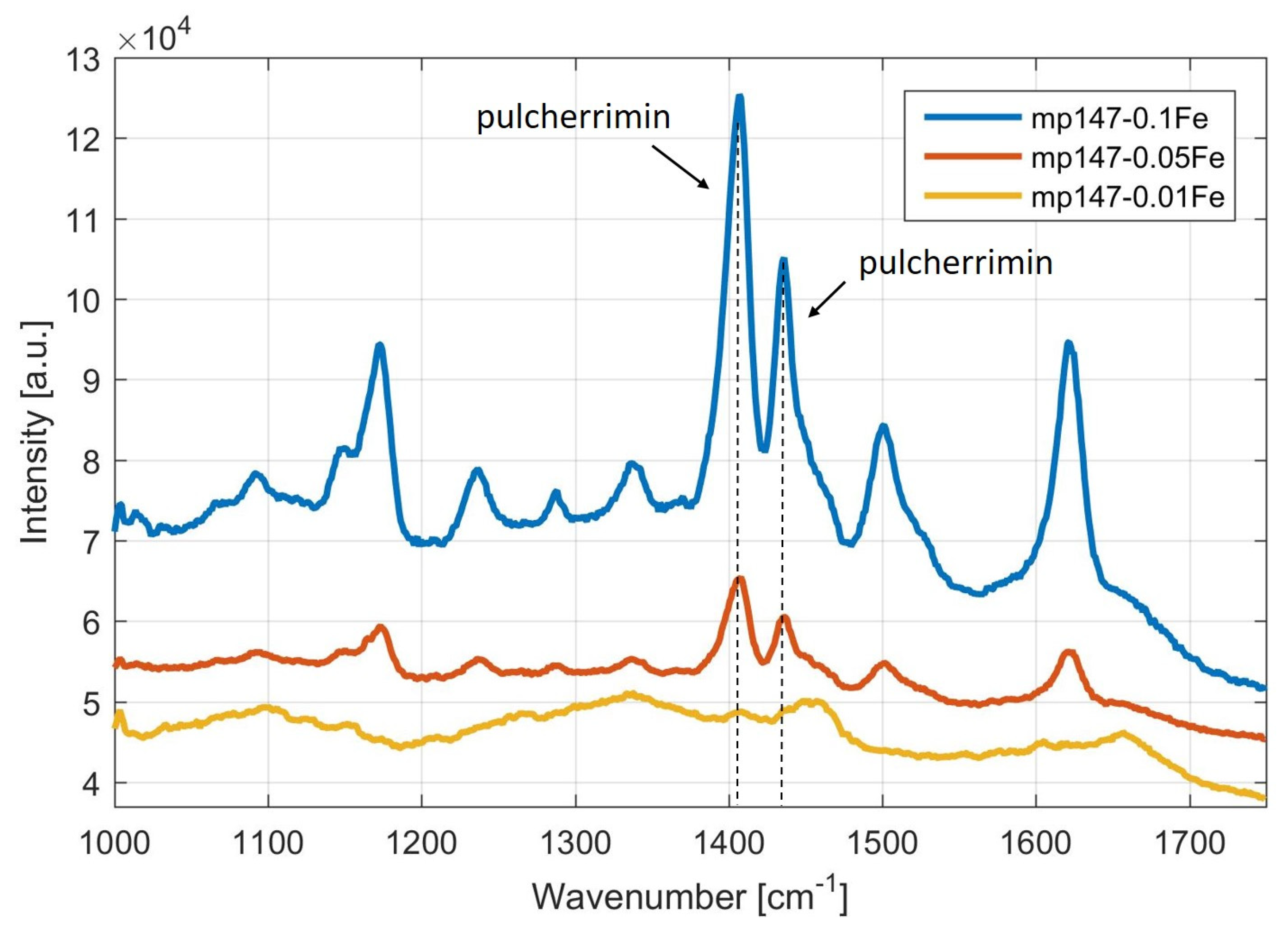
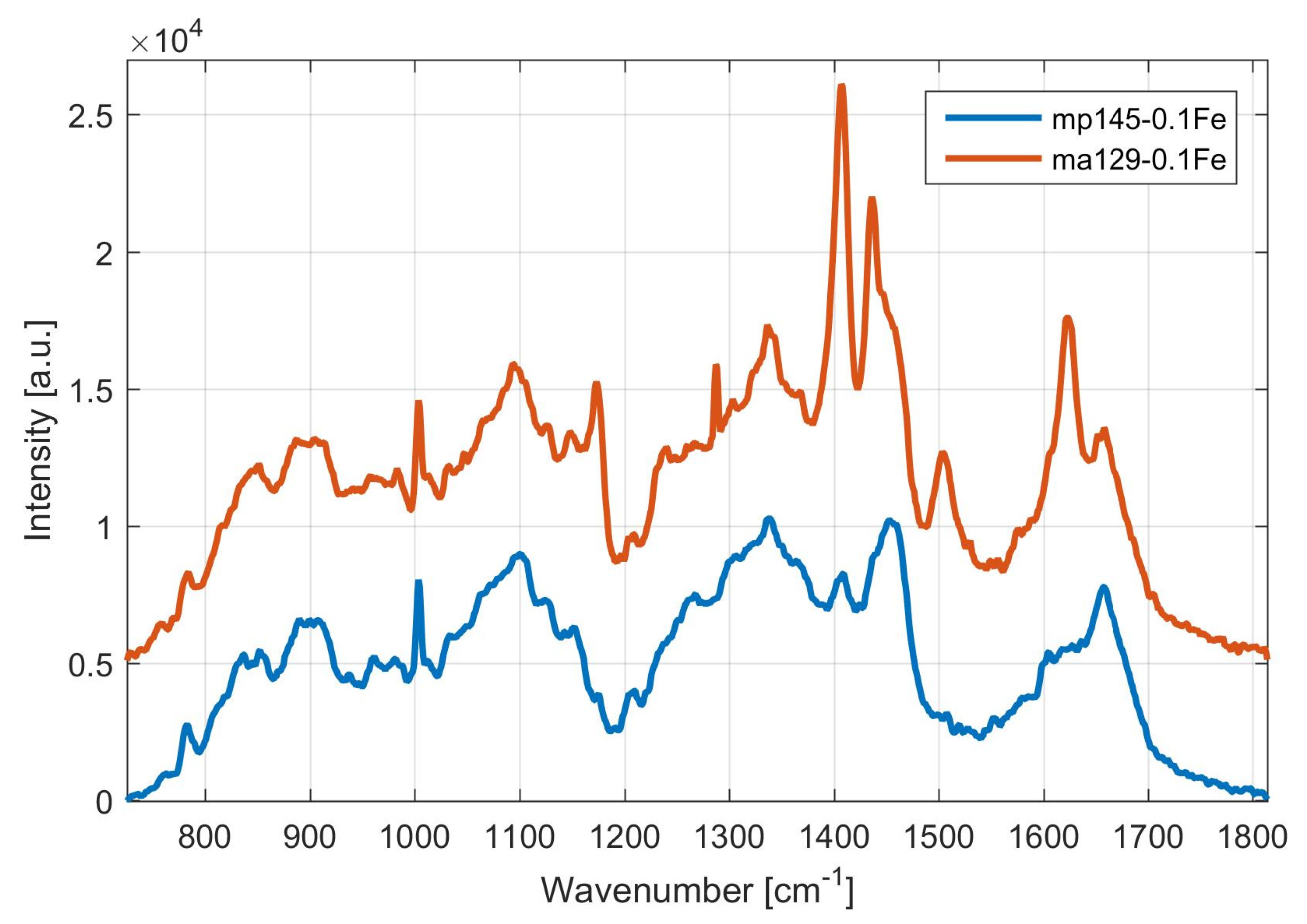
3.1.1. Screening and Identification of Pulcherrimin Production in the Presence of Iron Ions by Visual Observation and by Raman Spectroscopy
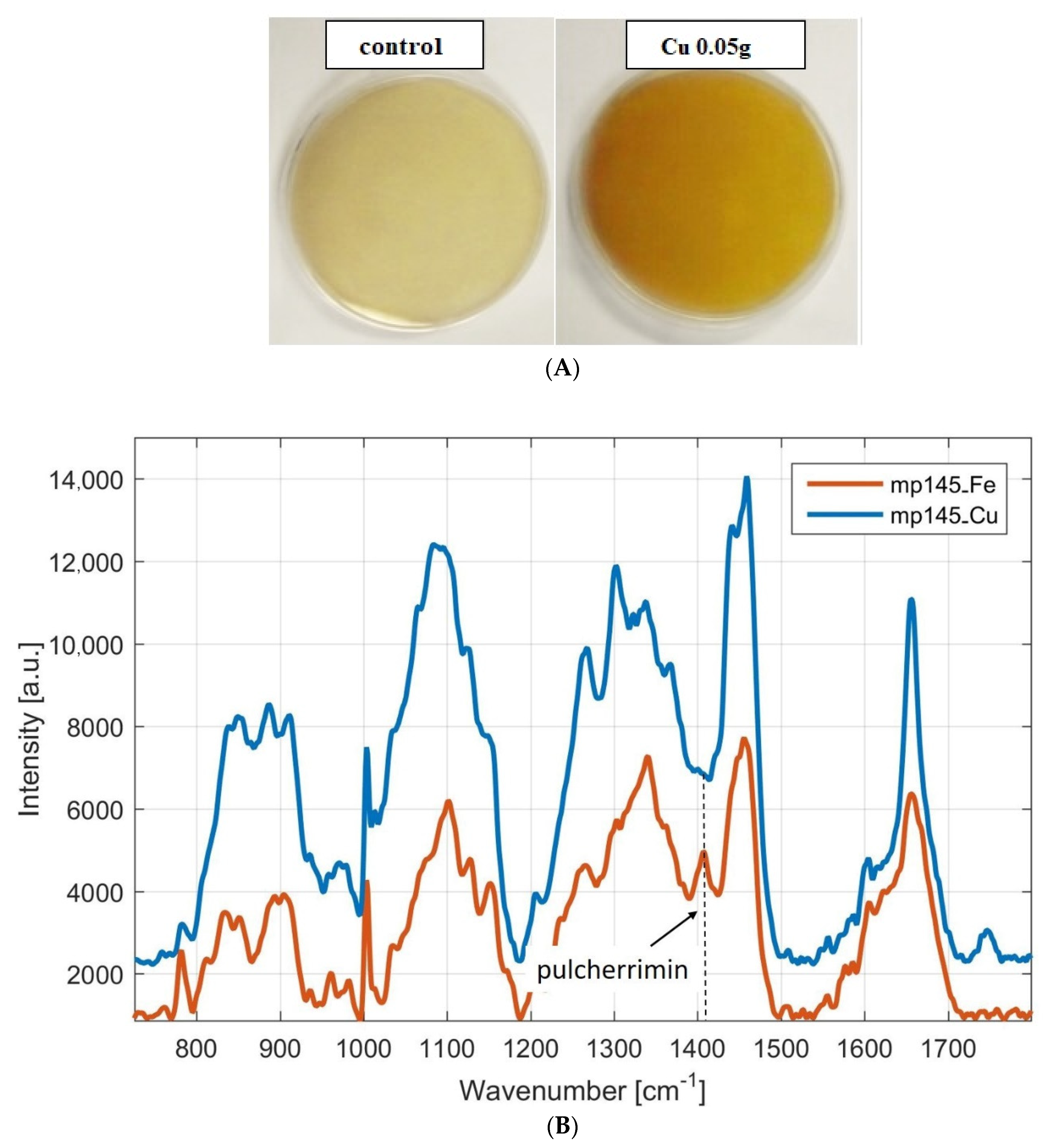
3.1.2. Monitoring of Pulcherrimin Production Depending on the Presence of Other Metal Ions in the Medium Visually and by Raman Spectroscopy
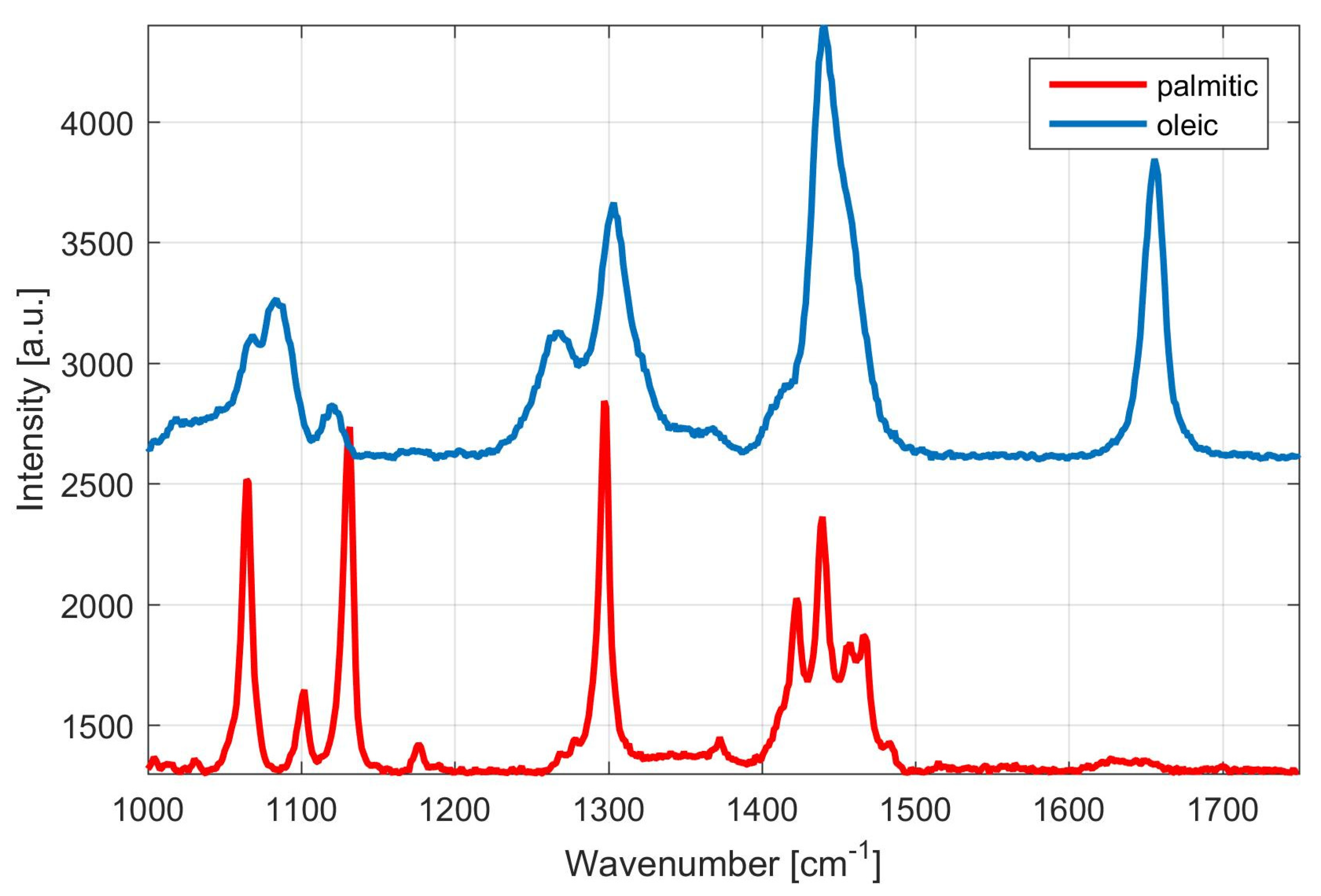
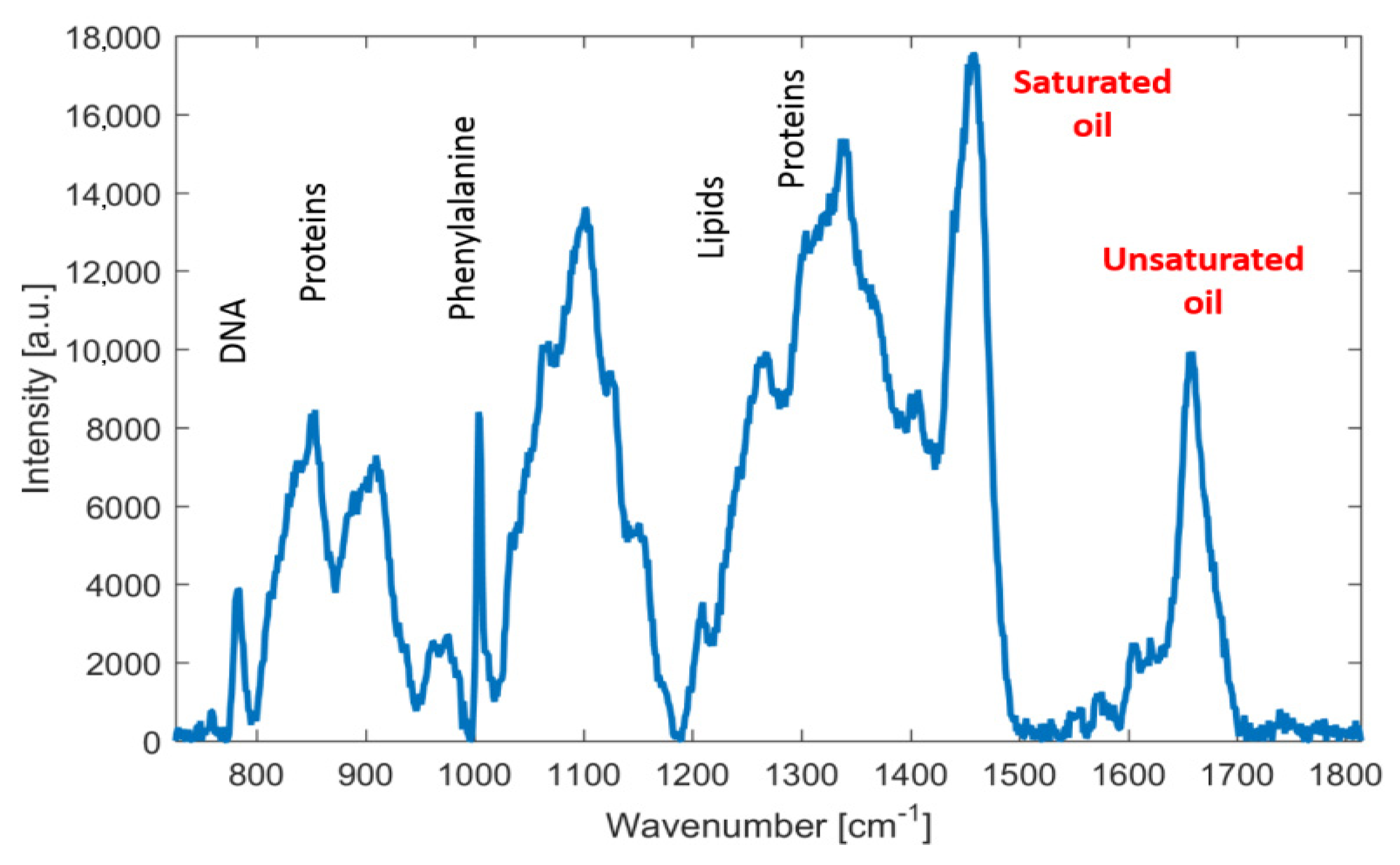
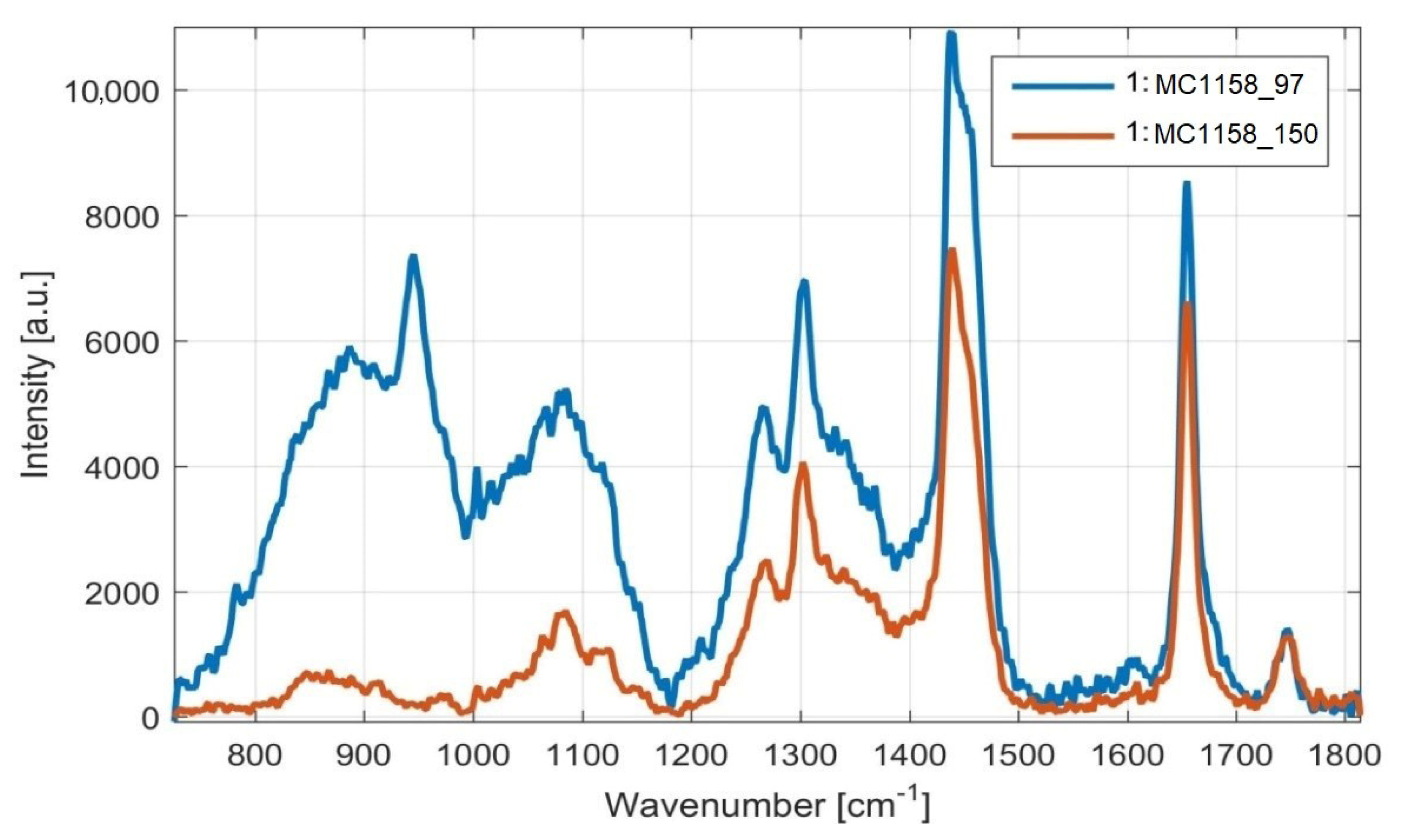
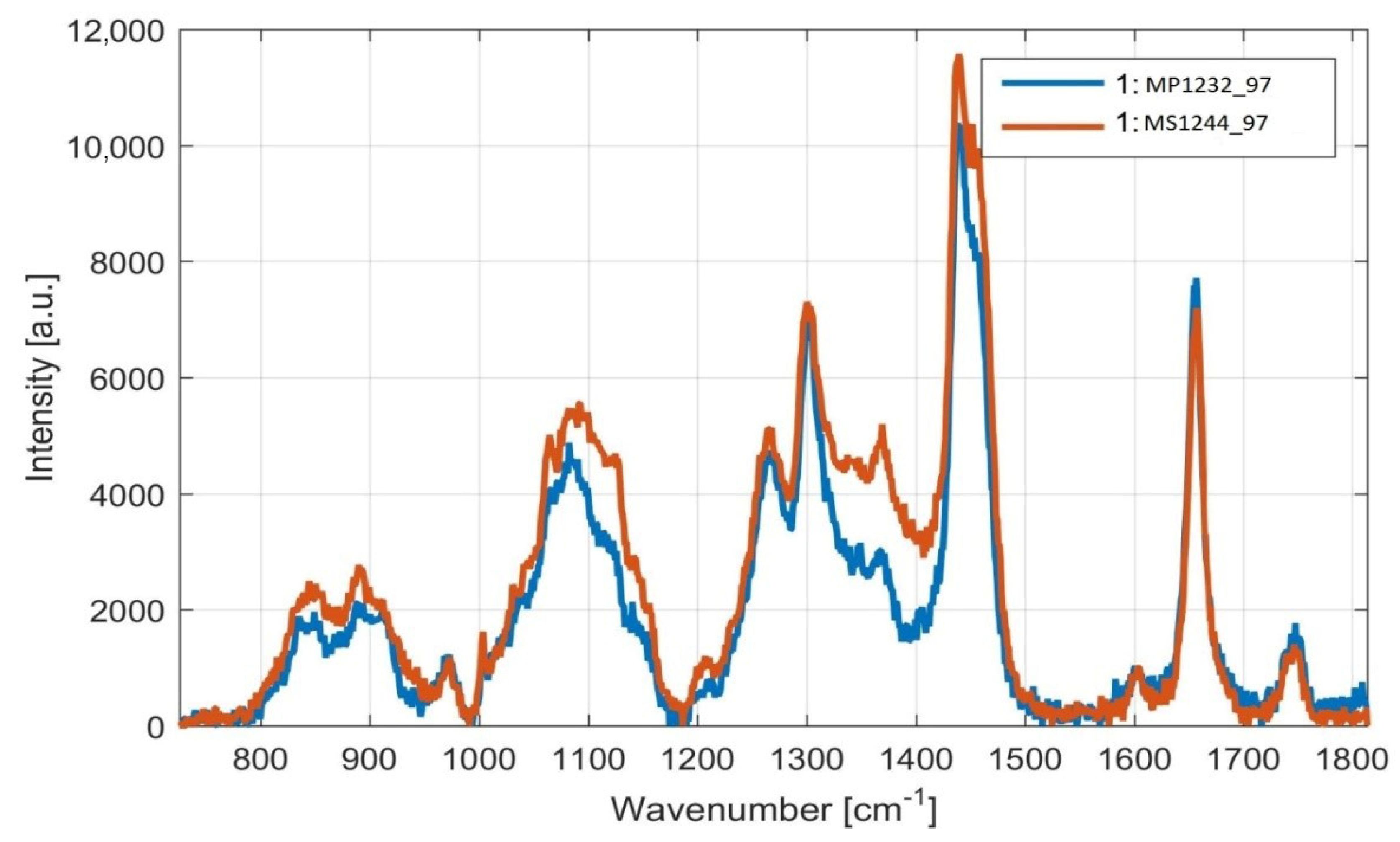
3.2. Rapid Monitoring of Lipid Production by Metschnikowia Yeasts, Using Raman Spectroscopy
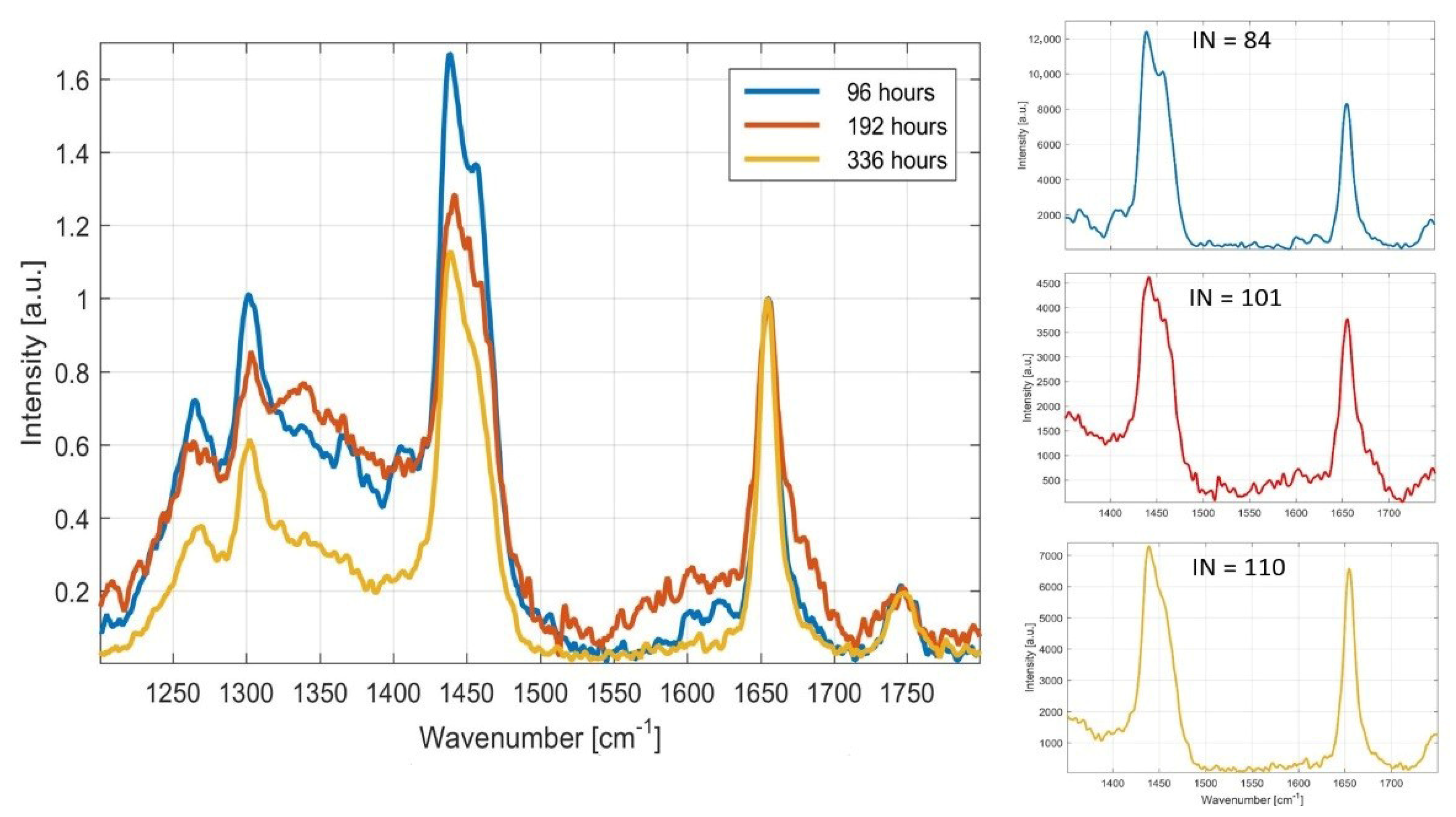
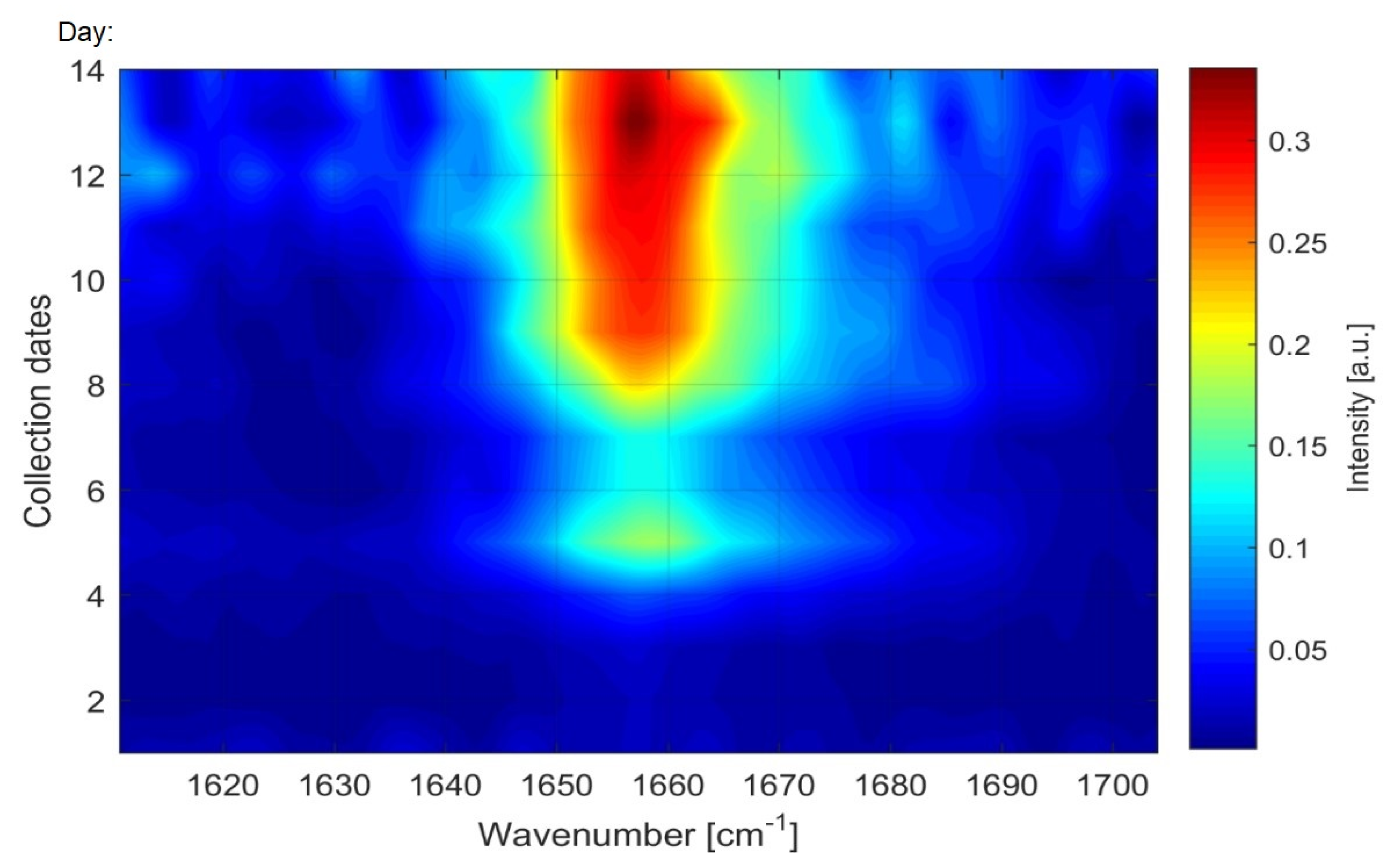
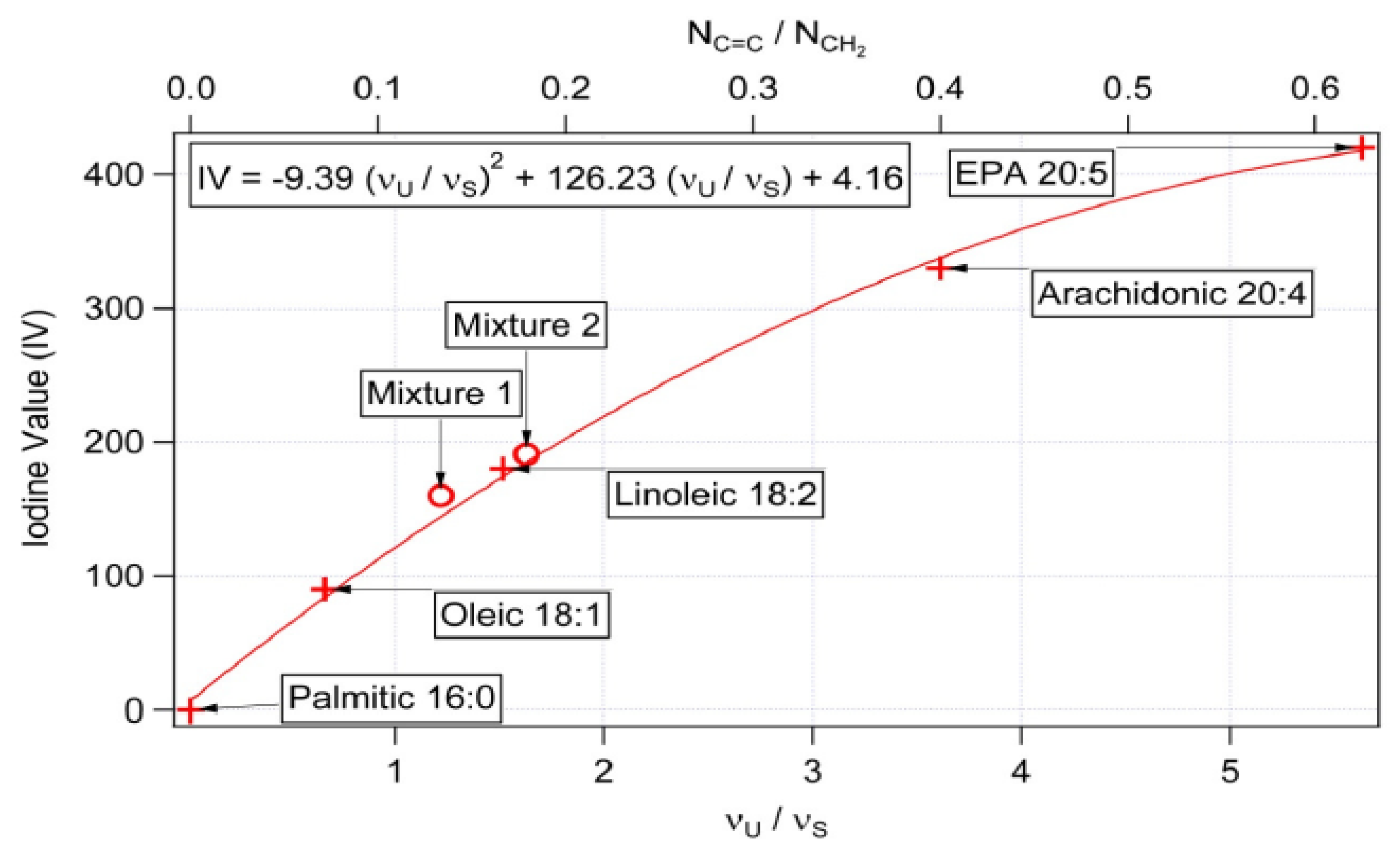
Monitoring of Fatty Acid Production Changes during Cultivation, Using Raman Spectroscopy
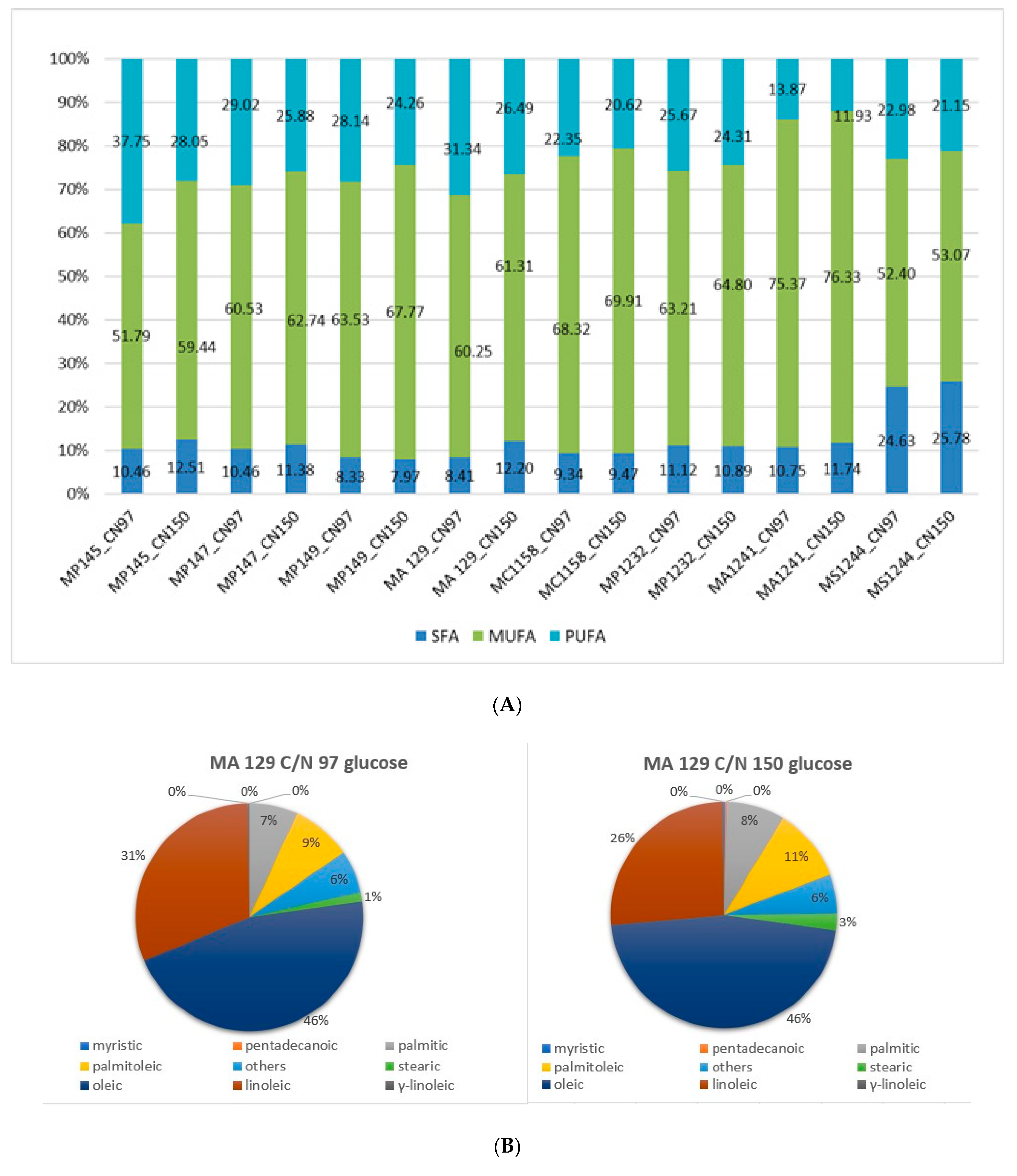
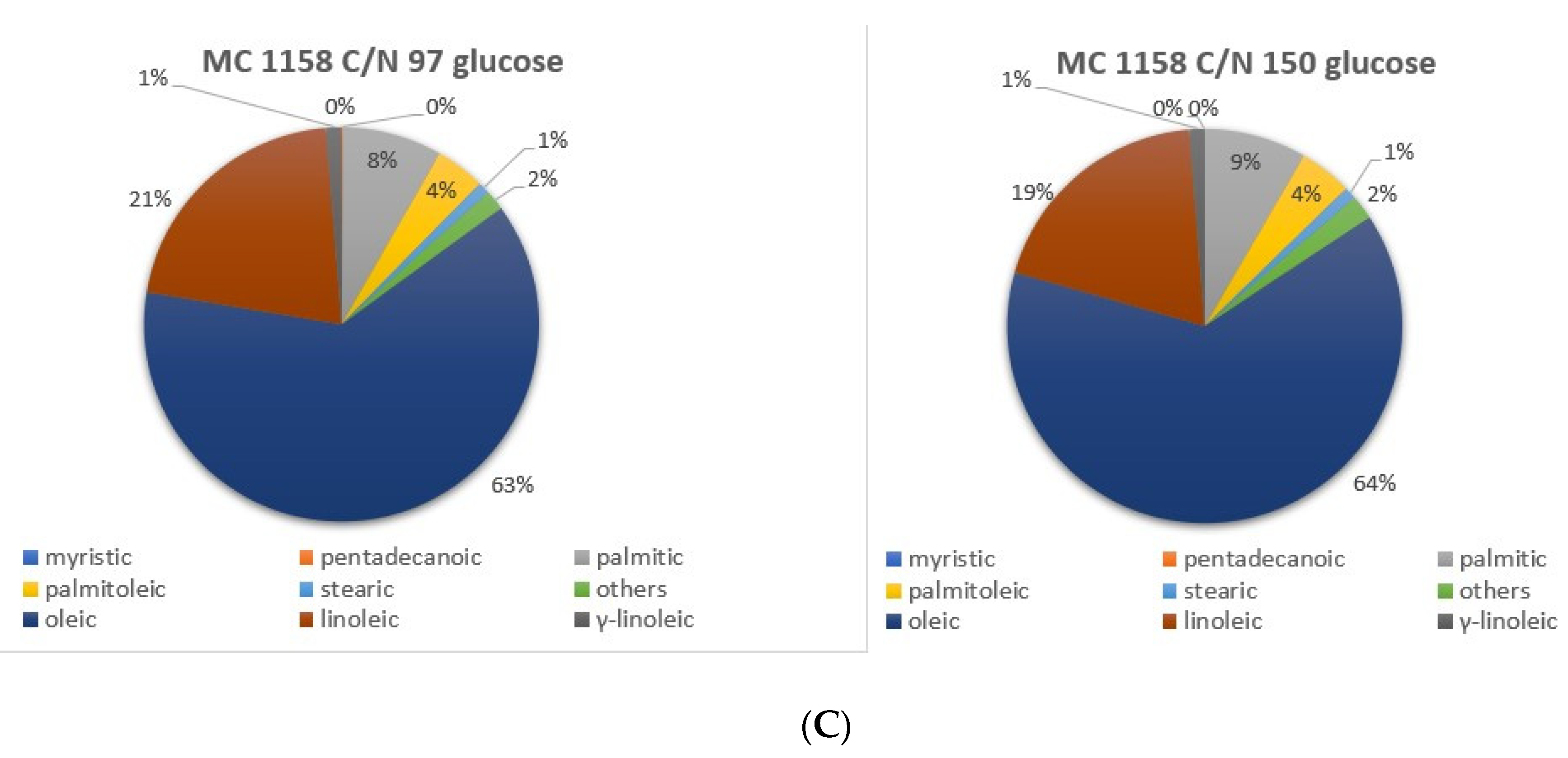
3.3. Analysis of Lipid Production and Fatty Acid Distribution at Different C/N Ratio
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Schie, I.W.; Huser, T. Methods and applications of Raman microspectroscopy to single-cell analysis. Appl. Spectrosc. 2013, 67, 813–828. [Google Scholar] [CrossRef] [PubMed]
- Notingher, I. Raman Spectroscopy Cell-based Biosensors. Sensors 2007, 7, 1343–1358. [Google Scholar] [CrossRef]
- Almarashi, J.; Kapel, N.; Wilkinson, T.S.; Telle, H. Raman Spectroscopy of Bacterial Species and Strains Cultivated under Reproducible Conditions. Spectrosc. Int. J. 2012, 27, 361–365. [Google Scholar] [CrossRef]
- De Gelder, J.; De Gussem, K.; Vandenabeele, P.; Moens, L. Reference database of Raman spectra of biological molecules. J. Raman Spectrosc. 2007, 38, 1133–1147. [Google Scholar] [CrossRef]
- Brauchle, E.; Schenke-Layland, K. Raman spectroscopy in biomedicine—Non-invasive in vitro analysis of cells and extracellular matrix components in tissues. Biotechnol. J. 2012, 8, 288–297. [Google Scholar] [CrossRef]
- Samek, O.; Al-Marashi, J.; Telle, H. The potential of Raman spectroscopy for the identification of biofilm formation by Staphylococcus epidermidis. Laser Phys. Lett. 2010, 7, 378–383. [Google Scholar] [CrossRef]
- Stuart, B. Gas Chromatography; Royal Society of Chemistry: Cambridge, UK, 2003; ISBN 08-540-4478-7. [Google Scholar]
- Littlewood, A.B. Gas Chromatography: Principles, Techniques, and Applications, 2nd ed.; Academic Press: New York, NY, USA, 1970. [Google Scholar]
- Li-Chan, E. The applications of Raman spectroscopy in food science. Trends Food Sci. Technol. 1996, 7, 361–370. [Google Scholar] [CrossRef]
- Bumbrah, G.S.; Sharma, R.M. Raman spectroscopy—Basic principle, instrumentation and selected applications for the characterization of drugs of abuse. Egypt. J. Forensic. Sci. 2016, 6, 209–215. [Google Scholar] [CrossRef]
- Kang, Y.; Choi, J.E.; Komakech, R.; Park, J.H.; Kim, D.W.; Cho, K.M.; Kang, S.M.; Choi, S.H.; Song, K.C.; Ryu, C.-M.; et al. Characterization of a novel yeast species Metschnikowia persimmonesis KCTC 12991BP (KIOM G15050 type strain) isolated from a medicinal plant, Korean persimmon calyx (Diospyros kaki Thumb). AMB Express 2017, 7, 199. [Google Scholar] [CrossRef]
- Lachance, M.-A. Metschnikowia: Half tetrads, a regicide and the fountain of youth. Yeast 2016, 33, 563–574. [Google Scholar] [CrossRef]
- Manso, T.; Nunes, C. Metschnikowia andauensis as a new biocontrol agent of fruit postharvest diseases. Postharvest Biol. Technol. 2011, 61, 64–71. [Google Scholar] [CrossRef]
- Kluyver, A.J.; van der Walt, J.P.; van Triet, A.J. Pulcherrimin, the Pigment of Candida Pulcherrima. Proc. Natl. Acad. Sci. USA 1953, 39, 583–593. [Google Scholar] [CrossRef]
- Macdonald, J. Biosynthesis of pulcherriminic acid. Biochem. J. 1965, 96, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.H.; Slater, C.A. Metabolism of “Wild” Yeasts I. The Chemical Nature of Pulcherrimin. J. Inst. Brew. 1954, 60, 213–217. [Google Scholar] [CrossRef]
- Cook, A.H.; Slater, C.A. Pulcherrimin: A synthesis of 1:4-dihydroxy-2:5-dioxopiperazines. J. Chem. Soc. 1956, 797, 4130–4133. [Google Scholar] [CrossRef]
- Cook, A.H.; Slater, C.A. The structure of pulcherrimin. J. Chem. Soc. 1956, 798, 4133. [Google Scholar] [CrossRef]
- Van der Walt, J.P. On the Yeast Candida Pulcherrima and Its Pigment; Excelsiors Foto-Offset: Delft, The Netherlands; The Hague, The Netherlands, 1952. [Google Scholar]
- Kupfer, D.G.; Uffen, R.L.; Canale-Parola, E. The role of iron and molecular oxygen in pulcherrimin synthesis by bacteria. Arch. Microbiol. 1967, 56, 9–21. [Google Scholar] [CrossRef]
- Bonnefond, L.; Arai, T.; Sakaguchi, Y.; Suzuki, T.; Ishitani, R.; Nureki, O. Structural basis for nonribosomal peptide synthesis by an aminoacyl-tRNA synthetase paralog. Proc. Natl. Acad. Sci. USA 2011, 108, 3912–3917. [Google Scholar] [CrossRef]
- Sipiczki, M. Metschnikowia Strains Isolated from Botrytized Grapes Antagonize Fungal and Bacterial Growth by Iron Depletion. Appl. Environ. Microbiol. 2006, 72, 6716–6724. [Google Scholar] [CrossRef]
- Certik, M.; Shimizu, S. Biosynthesis and regulation of microbial poly-unsaturated fatty acid production. J. Biosci. Bioeng. 1999, 87, 1–14. [Google Scholar] [CrossRef]
- Ochsenreither, K.; Glück, C.; Stressler, T.; Fischer, L.; Syldatk, C. Production Strategies and Applications of Microbial Single Cell Oils. Front. Microbiol. 2016, 7, 1539. [Google Scholar] [CrossRef]
- Garay, L.A.; Boundy-Mills, K.L.; German, J.B. Accumulation of High-Value Lipids in Single-Cell Microorganisms: A Mechanistic Approach and Future Perspectives. J. Agric. Food Chem. 2014, 62, 2709–2727. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Yarrowia lipolytica: A model microorganism used for the production of tailor-made lipids. Eur. J. Lipid. Sci. Technol. 2010, 112, 639–654. [Google Scholar] [CrossRef]
- Maina, S.; Pateraki, C.; Kopsahelis, N.; Paramithiotis, S.; Drosinos, E.H.; Papanikolaou, S.; Koutinas, A. Microbial oil production from various carbon sources by newly isolated oleaginous yeasts. Eng. Life Sci. 2017, 17, 333–344. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Stanley, G.S. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Szotkowski, M.; Byrtusová, D.; Haronikova, A.; Vysoka, M.; Rapta, M.; Shapaval, V.; Marova, I. Study of Metabolic Adaptation of Red Yeasts to Waste Animal Fat Substrate. Microorganisms 2019, 7, 578. [Google Scholar] [CrossRef]
- Brandt, N.N.; Brovko, O.O.; Chikishev, A.Y.; Paraschuk, O.D. Optimization of the Rolling-Circle Filter for Raman Background Subtraction. Appl. Spectrosc. 2006, 60, 288–293. [Google Scholar] [CrossRef]
- Samek, O.; Jonáš, A.; Pilát, Z.; Zemánek, P.; Nedbal, L.; Triska, J.; Kotas, P.; Trtilek, M. Raman Microspectroscopy of Individual Algal Cells: Sensing Unsaturation of Storage Lipids in vivo. Sensors 2010, 10, 8635–8651. [Google Scholar] [CrossRef]
- TÜRKEL, S.; Ener, B. Isolation and Characterization of New Metschnikowia pulcherrima Strains as Producers of the Antimicrobial Pigment Pulcherrimin. Z. Naturforsch. C 2009, 64, 405–410. [Google Scholar] [CrossRef]
- Macdonald, J.C. The Structure of Pulcherriminic Acid. Can. J. Chem. 1963, 41, 165–172. [Google Scholar] [CrossRef]
- Zdaniauskienėa, A.; Charkovaa, T.; Ignatjeva, I.; Melvydasb, V.; Garjonytėa, R.; Matulaitienėa, I.; Talaikisc, M.; Niaura, G. Shell-isolated nanoparticle-enhanced Raman spectroscopy for characterization of living yeast cells. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2020, 240, 118560. [Google Scholar] [CrossRef]
- Vicente, J.; Ruiz, J.; Belda, I.; Benito-Vázquez, I.; Marquina, D.; Calderón, F.; Santos, A.; Benito, S. The Genus Metschnikowia in Enology. Microorganisms 2020, 8, 1038. [Google Scholar] [CrossRef]
- Sipiczki, M. Metschnikowia pulcherrima and Related Pulcherrimin-Producing Yeasts: Fuzzy Species Boundaries and Complex Antimicrobial Antagonism. Microorganisms 2020, 8, 1029. [Google Scholar] [CrossRef]
- Santamauro, F.; Whiffin, F.M.; Scott, R.J.; Chuck, C.J.; Antonucci, M.; Truzzi, C.; Scarponi, G.; Ciani, M. Low-cost lipid production by an oleaginous yeast cultured in non-sterile conditions using model waste resources. Biotechnol. Biofuels 2014, 7, 34. [Google Scholar] [CrossRef]
- Canonico, L.; Ashoor, S.; Taccari, M.; Comitini, F.; Antonucci, M.; Truzzi, C.; Scarponi, G.; Ciani, M. Conversion of raw glycerol to microbial lipids by new Metschnikowia and Yarrowia lipolytica strains. Ann. Microbiol. 2016, 66, 1409–1418. [Google Scholar] [CrossRef]
- Hilal, N.; Ismail, A.F.; Matsuura, T.; Oatley-Radcliffe, D. Raman Spectroscopy. Membrane Characterization; Elsevier: Amsterdam, The Netherlands, 2017; pp. 31–46. ISBN 9780444637765. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Day of Cultivation | ||
|---|---|---|---|
| 4 | 8 | 14 | |
| C15:0 | - | - | 0.14 |
| C16:0 | 14.1 | 13.26 | 10 |
| C16:1n7 | 4.45 | 4.51 | 4.69 |
| C18:0 | 12.4 | 11.48 | 6.4 |
| C18:1n9 | 45.51 | 43.97 | 39 |
| C18:2n6 | 19.07 | 22.73 | 31.93 |
| C18:3n6 | - | 0.06 | 0.14 |
| C18:3n3 | 0.91 | 1.28 | 2.65 |
| SFA * | 27.43 | 25.34 | 18.66 |
| MUFA * | 53 | 50.48 | 46.55 |
| PUFA * | 19.98 | 24.17 | 34.8 |
| Strain | C/N Ratio | Biomass (g/L) | Lipid Content (%) |
|---|---|---|---|
| M. pulcherrima 149 | 24 | 10.4 ± 0.6 | 5.6 ± 0.5 |
| 97 | 7.4 ± 0.5 | 4.4 ± 0.6 | |
| 150 | 7.2 ± 0.5 | 8.4 ± 0.4 | |
| M. andauensis 129 | 24 | 12.4 ± 0.5 | 2.2 ± 0.2 |
| 97 | 6.6 ± 0.5 | 4.0 ± 0.3 | |
| 150 | 6.2 ± 0.4 | 3.5 ± 0.2 | |
| M. chrysoperlae 1158 | 97 | 6.9 ± 0.7 | 12.3 ± 2.4 |
| 150 | 7.1 ± 0.5 | 13.6 ± 0.9 | |
| M. pulcherrima 1232 | 97 | 7.0 ± 0.5 | 14.6 ± 1.9 |
| 150 | 6.5 ± 0.4 | 15.4 ± 1.0 | |
| M. sinensis 1244 | 97 | 10.3 ± 1.2 | 14.4 ± 2.5 |
| 150 | 9.9 ± 0.6 | 16.9 ± 1.5 | |
| M. fructicola 1235 | 97 | 15.2 ± 1.4 | 2.7 ± 0.4 |
| 150 | 5.6 ± 0.6 | 4.9 ± 1.3 | |
| M. andauensis 1241 | 97 | 9.3 ± 0.8 | 17.0 ± 2.2 |
| 150 | 8.2 ± 0.5 | 19.6 ± 2.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Němcová, A.; Gonová, D.; Samek, O.; Sipiczki, M.; Breierová, E.; Márová, I. The Use of Raman Spectroscopy to Monitor Metabolic Changes in Stressed Metschnikowia sp. Yeasts. Microorganisms 2021, 9, 277. https://doi.org/10.3390/microorganisms9020277
Němcová A, Gonová D, Samek O, Sipiczki M, Breierová E, Márová I. The Use of Raman Spectroscopy to Monitor Metabolic Changes in Stressed Metschnikowia sp. Yeasts. Microorganisms. 2021; 9(2):277. https://doi.org/10.3390/microorganisms9020277
Chicago/Turabian StyleNěmcová, Andrea, Dominika Gonová, Ota Samek, Matthias Sipiczki, Emilia Breierová, and Ivana Márová. 2021. "The Use of Raman Spectroscopy to Monitor Metabolic Changes in Stressed Metschnikowia sp. Yeasts" Microorganisms 9, no. 2: 277. https://doi.org/10.3390/microorganisms9020277
APA StyleNěmcová, A., Gonová, D., Samek, O., Sipiczki, M., Breierová, E., & Márová, I. (2021). The Use of Raman Spectroscopy to Monitor Metabolic Changes in Stressed Metschnikowia sp. Yeasts. Microorganisms, 9(2), 277. https://doi.org/10.3390/microorganisms9020277