
Long-Term Sinonasal Carriage of Staphylococcus aureus and Anti-Staphylococcal Humoral Immune Response in Patients with Chronic Rhinosinusitis

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Serum and Bacterial Samples from Patients with CRS
2.2. Antibiotic Susceptibility Testing
2.3. DNA Sequencing and Single-Nucleotide Polymorphism (SNP) Analysis
2.4. Microarray-Based Immunoglobulin Analysis
2.5. Statistical Analysis
2.6. Ethical Approval
3. Results
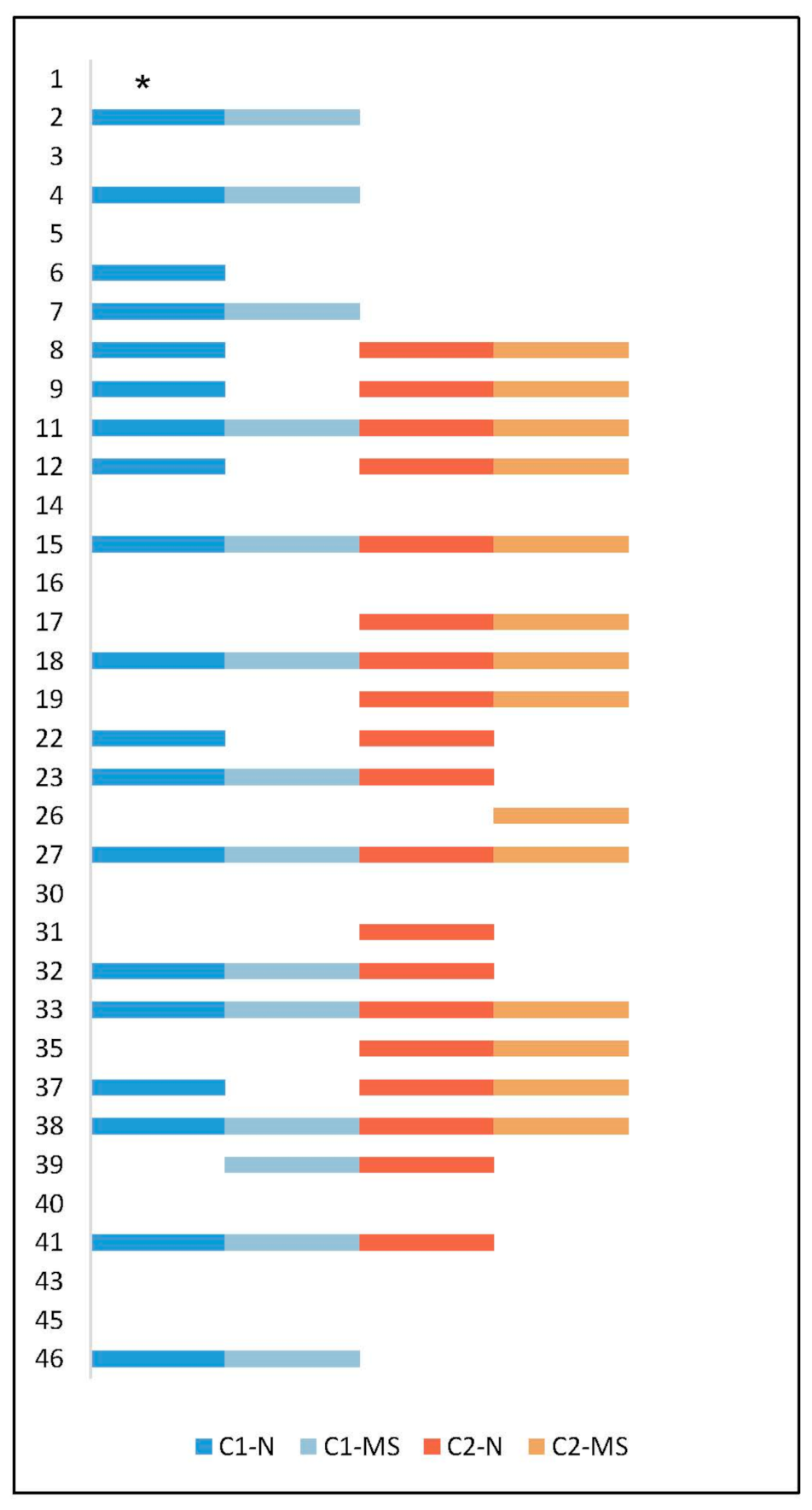
3.1. Presence of S. aureus in CRS Patients and Antibiotic Susceptibility
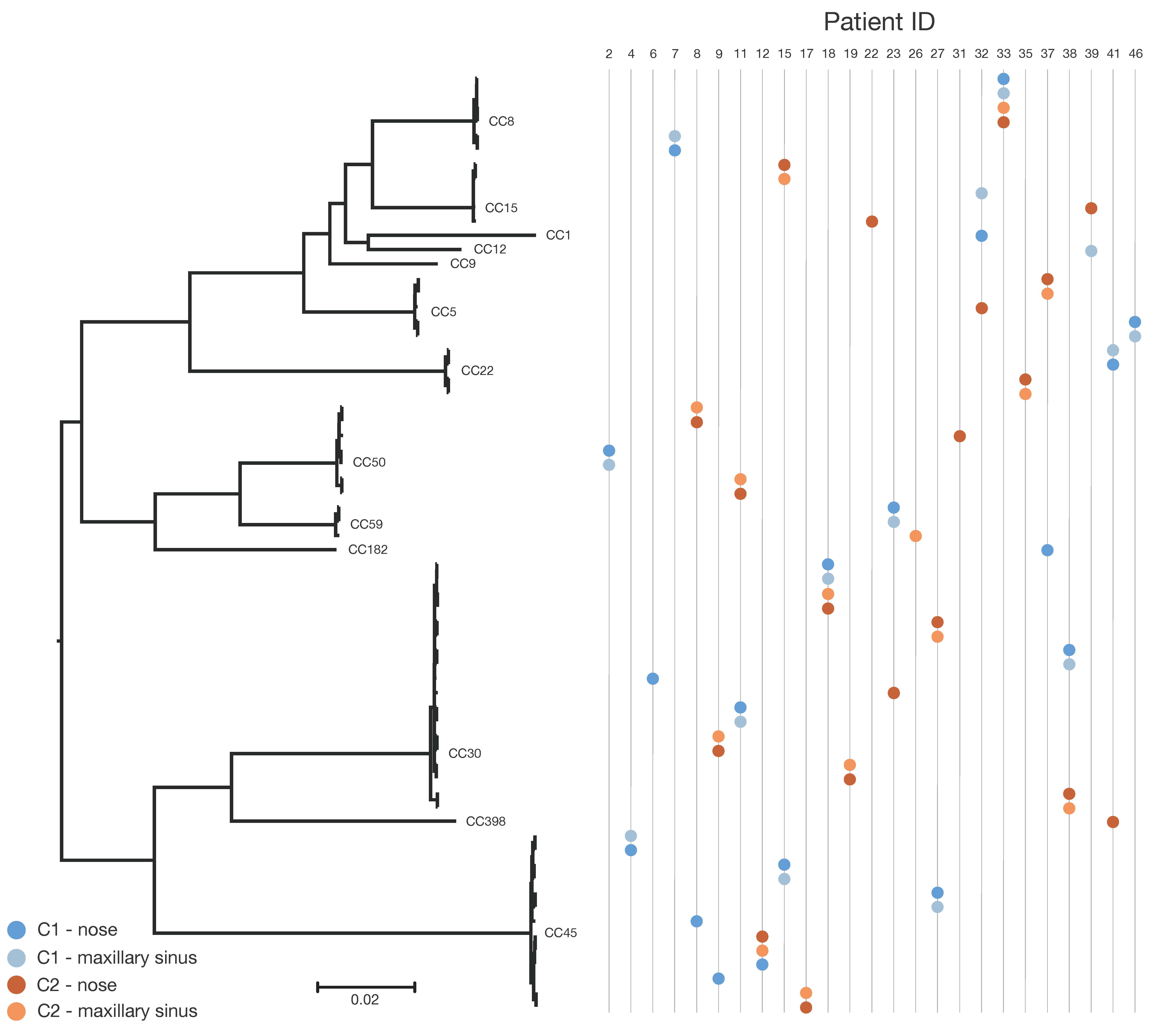
3.2. SNP-Based Phylogenetic Analysis
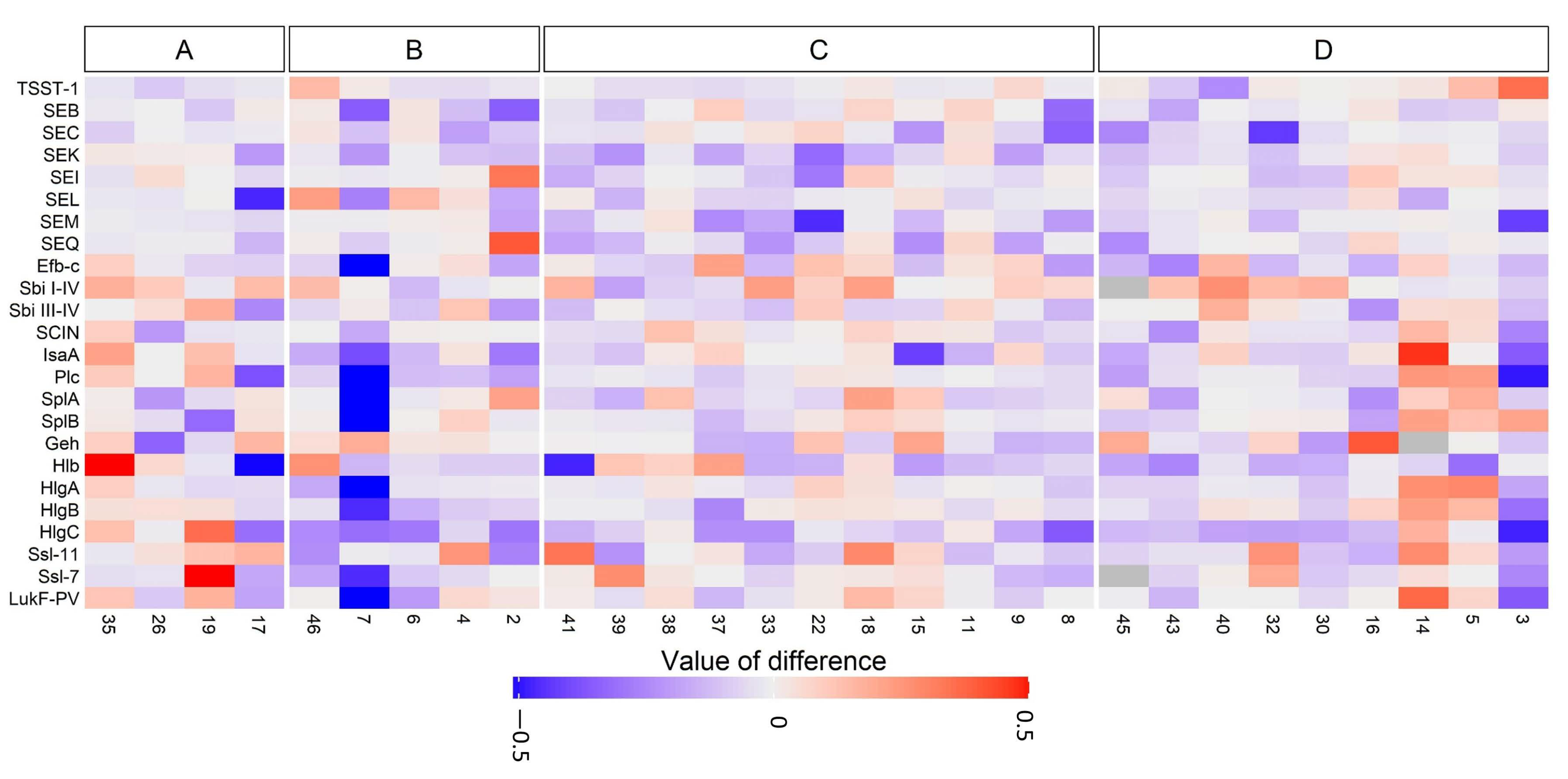
3.3. Anti-Staphylococcal Antibodies in CRS Patients
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Derycke, L.; Perez-Novo, C.; Van Crombruggen, K.; Corriveau, M.N.; Bachert, C. Staphylococcus aureus and chronic airway disease. World Allergy Organ. J. 2010, 3, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Abou-Hamad, W.; Matar, N.; Elias, M.; Nasr, M.; Sarkis-Karam, D.; Hokayem, N.; Haddad, A. Bacterial flora in normal adult maxillary sinuses. Am. J. Rhinol. Allergy 2009, 23, 261–263. [Google Scholar] [CrossRef] [PubMed]
- Thanasumpun, T.; Batra, P.S. Endoscopically-derived bacterial cultures in chronic rhinosinusitis: A systematic review. Am. J. Otolaryngol. 2015, 36, 686–691. [Google Scholar] [CrossRef]
- Thunberg, U.; Soderquist, B.; Hugosson, S. Bacterial findings in optimised sampling and characterisation of S. aureus in chronic rhinosinusitis. Eur. Arch. Otorhinolaryngol. 2017, 274, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Biel, M.A.; Brown, C.A.; Levinson, R.M.; Garvis, G.E.; Paisner, H.M.; Sigel, M.E.; Tedford, T.M. Evaluation of the microbiology of chronic maxillary sinusitis. Ann. Otol. Rhinol. Laryngol. 1998, 107 Pt 1, 942–945. [Google Scholar] [CrossRef]
- Kluytmans, J.; van Belkum, A.; Verbrugh, H. Nasal carriage of Staphylococcus aureus: Epidemiology, underlying mechanisms, and associated risks. Clin. Microbiol. Rev. 1997, 10, 505–520. [Google Scholar] [CrossRef]
- Wertheim, H.F.; Melles, D.C.; Vos, M.C.; van Leeuwen, W.; van Belkum, A.; Verbrugh, H.A.; Nouwen, J.L. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect. Dis. 2005, 5, 751–762. [Google Scholar] [CrossRef]
- Liu, C.M.; Price, L.B.; Hungate, B.A.; Abraham, A.G.; Larsen, L.A.; Christensen, K.; Stegger, M.; Skov, R.; Andersen, P.S. Staphylococcus aureus and the ecology of the nasal microbiome. Sci. Adv. 2015, 1, e1400216. [Google Scholar] [CrossRef]
- Lowy, F.D. Staphylococcus aureus infections. N. Engl. J. Med. 1998, 339, 520–532. [Google Scholar] [CrossRef]
- Laudien, M.; Gadola, S.D.; Podschun, R.; Hedderich, J.; Paulsen, J.; Reinhold-Keller, E.; Csernok, E.; Ambrosch, P.; Hellmich, B.; Moosig, F.; et al. Nasal carriage of Staphylococcus aureus and endonasal activity in Wegener’s granulomatosis as compared to rheumatoid arthritis and chronic rhinosinusitis with nasal polyps. Clin. Exp. Rheumatol. 2010, 28 (Suppl. 57), 51–55. [Google Scholar]
- Seiti Yamada Yoshikawa, F.; Feitosa de Lima, J.; Notomi Sato, M.; Alefe Leuzzi Ramos, Y.; Aoki, V.; Leao Orfali, R. Exploring the role of Staphylococcus aureus toxins in atopic dermatitis. Toxins 2019, 11, 321. [Google Scholar] [CrossRef] [PubMed]
- Tabarya, D.; Hoffman, W.L. Staphylococcus aureus nasal carriage in rheumatoid arthritis: Antibody response to toxic shock syndrome toxin-1. Ann. Rheum. Dis. 1996, 55, 823–828. [Google Scholar] [CrossRef] [PubMed]
- Bachert, C.; Claeys, S.E.; Tomassen, P.; van Zele, T.; Zhang, N. Rhinosinusitis and asthma: A link for asthma severity. Curr. Allergy Asthma Rep. 2010, 10, 194–201. [Google Scholar] [CrossRef]
- Bachert, C.; Holtappels, G. Pathophysiology of chronic rhinosinusitis, pharmaceutical therapy options. GMS Curr. Top. Otorhinolaryngol. Head Neck Surg. 2015, 14, 1–40. [Google Scholar] [CrossRef]
- Bernstein, J.M.; Kansal, R. Superantigen hypothesis for the early development of chronic hyperplastic sinusitis with massive nasal polyposis. Curr. Opin. Otolaryngol. Head Neck Surg. 2005, 13, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Bachert, C.; Gevaert, P.; van Cauwenberge, P. Staphylococcus aureus enterotoxins: A key in airway disease? Allergy 2002, 57, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.; Rasooly, A. Staphylococcal enterotoxins. Int. J. Food Microbiol. 2000, 61, 1–10. [Google Scholar] [CrossRef]
- Foster, T.J.; McDevitt, D. Surface-associated proteins of Staphylococcus aureus: Their possible roles in virulence. FEMS Microbiol. Lett. 1994, 118, 199–205. [Google Scholar] [CrossRef]
- Oliveira, D.; Borges, A.; Simoes, M. Staphylococcus aureus toxins and their molecular activity in infectious diseases. Toxins 2018, 10, 252. [Google Scholar] [CrossRef]
- Bachert, C.; Gevaert, P.; Holtappels, G.; Johansson, S.G.; van Cauwenberge, P. Total and specific IgE in nasal polyps is related to local eosinophilic inflammation. J. Allergy Clin. Immunol. 2001, 107, 607–614. [Google Scholar] [CrossRef]
- Foreman, A.; Holtappels, G.; Psaltis, A.J.; Jervis-Bardy, J.; Field, J.; Wormald, P.J.; Bachert, C. Adaptive immune responses in Staphylococcus aureus biofilm-associated chronic rhinosinusitis. Allergy 2011, 66, 1449–1456. [Google Scholar] [CrossRef]
- Foreman, A.; Jervis-Bardy, J.; Wormald, P.J. Do biofilms contribute to the initiation and recalcitrance of chronic rhinosinusitis? Laryngoscope 2011, 121, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Thunberg, U.; Hugosson, S.; Fredlund, H.; Cao, Y.; Ehricht, R.; Monecke, S.; Muller, E.; Engelmann, S.; Söderquist, B. Anti-staphylococcal humoral immune response in patients with chronic rhinosinusitis. Rhinol. Online 2019, 2, 50–58. [Google Scholar] [CrossRef]
- Sahl, J.W.; Lemmer, D.; Travis, J.; Schupp, J.M.; Gillece, J.D.; Aziz, M.; Driebe, E.M.; Drees, K.P.; Hicks, N.D.; Williamson, C.H.D.; et al. NASP: An accurate, rapid method for the identification of SNPs in WGS datasets that supports flexible input and output formats. Microb. Genom. 2016, 2, e000074. [Google Scholar] [CrossRef] [PubMed]
- Stegger, M.; Driebe, E.M.; Roe, C.; Lemmer, D.; Bowers, J.R.; Engelthaler, D.M.; Keim, P.; Andersen, P.S. Genome sequence of Staphylococcus aureus strain CA-347, a USA600 methicillin-resistant isolate. Genome Announc. 2013, 1, e00517-13. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kloppot, P.; Selle, M.; Kohler, C.; Stentzel, S.; Fuchs, S.; Liebscher, V.; Müller, E.; Kale, D.; Ohlsen, K.; Bröker, B.M.; et al. Microarray-based identification of human antibodies against Staphylococcus aureus antigens. Proteom. Clin. Appl. 2015, 9, 1003–1011. [Google Scholar] [CrossRef]
- Selle, M.; Hertlein, T.; Oesterreich, B.; Klemm, T.; Kloppot, P.; Müller, E.; Ehricht, R.; Stentzel, S.; Bröker, B.M.; Engelmann, S.; et al. Global antibody response to Staphylococcus aureus live-cell vaccination. Sci. Rep. 2016, 6, 24754. [Google Scholar] [CrossRef]
- Holm, S. A simple sequentially rejective multiple test procedure. Scand. J. Stat. 1979, 6, 65–70. [Google Scholar]
- Duchêne, S.; Holt, K.E.; Weill, F.X.; Le Hello, S.; Hawkey, J.; Edwards, D.J.; Fourment, M.; Holmes, E.C. Genome-scale rates of evolutionary change in bacteria. Microb. Genom. 2016, 2, e000094. [Google Scholar] [CrossRef]
- Wertheim, H.F.; Verveer, J.; Boelens, H.A.; van Belkum, A.; Verbrugh, H.A.; Vos, M.C. Effect of mupirocin treatment on nasal, pharyngeal, and perineal carriage of Staphylococcus aureus in healthy adults. Antimicrob. Agents Chemother. 2005, 49, 1465–1467. [Google Scholar] [CrossRef] [PubMed]
- Eriksen, N.H.; Espersen, F.; Rosdahl, V.T.; Jensen, K. Carriage of Staphylococcus aureus among 104 healthy persons during a 19-month period. Epidemiol. Infect. 1995, 115, 51–60. [Google Scholar] [CrossRef]
- VandenBergh, M.F.; Yzerman, E.P.; van Belkum, A.; Boelens, H.A.; Sijmons, M.; Verbrugh, H.A. Follow-up of Staphylococcus aureus nasal carriage after 8 years: Redefining the persistent carrier state. J. Clin. Microbiol. 1999, 37, 3133–3140. [Google Scholar] [CrossRef]
- Harvey, R.J.; Snidvongs, K.; Kalish, L.H.; Oakley, G.M.; Sacks, R. Corticosteroid nasal irrigations are more effective than simple sprays in a randomized double-blinded placebo-controlled trial for chronic rhinosinusitis after sinus surgery. Int. Forum. Allergy Rhinol. 2018, 8, 461–470. [Google Scholar] [CrossRef]
- Lees, K.A.; Orlandi, R.R.; Oakley, G.; Alt, J.A. The role of macrolides and doxycycline in chronic rhinosinusitis. Immunol. Allergy Clin. N. Am. 2020, 40, 303–315. [Google Scholar] [CrossRef]
- Holtfreter, S.; Grumann, D.; Schmudde, M.; Nguyen, H.T.; Eichler, P.; Strommenger, B.; Kopron, K.; Kolata, J.; Giedrys-Kalemba, S.; Steinmetzet, I.; et al. Clonal distribution of superantigen genes in clinical Staphylococcus aureus isolates. J. Clin. Microbiol. 2007, 45, 2669–2680. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, G.; Monecke, S.; Ehricht, R.; Soderquist, B. Prevalence of clonal complexes and virulence genes among commensal and invasive Staphylococcus aureus isolates in Sweden. PLoS ONE 2013, 8, e77477. [Google Scholar] [CrossRef] [PubMed]
- Acton, D.S.; Plat-Sinnige, M.J.; van Wamel, W.; de Groot, N.; van Belkum, A. Intestinal carriage of Staphylococcus aureus: How does its frequency compare with that of nasal carriage and what is its clinical impact? Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Dancer, S.J.; Noble, W.C. Nasal, axillary, and perineal carriage of Staphylococcus aureus among women: Identification of strains producing epidermolytic toxin. J. Clin. Pathol. 1991, 44, 681–684. [Google Scholar] [CrossRef]
- Verkaik, N.J.; de Vogel, C.P.; Boelens, H.A.; Grumann, D.; Hoogenboezem, T.; Vink, C.; Hooijkaas, H.; Foster, T.; Verbrugh, H.; Van Belkum, A.; et al. Anti-staphylococcal humoral immune response in persistent nasal carriers and noncarriers of Staphylococcus aureus. J. Infect. Dis. 2009, 199, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Soderquist, B.; Danielsson, D.; Holmberg, H.; Vikerfors, T. Granulocyte colony-stimulating factor (G-CSF) and interleukin (IL)-8 in sera from patients with Staphylococcus aureus septicemia. Clin. Microbiol. Infect. 1995, 1, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Dryla, A.; Prustomersky, S.; Gelbmann, D.; Hanner, M.; Bettinger, E.; Kocsis, B.; Kustos, T.; Henics, T.; Meinke, A.; Nagy, E. Comparison of antibody repertoires against Staphylococcus aureus in healthy individuals and in acutely infected patients. Clin. Diagn. Lab. Immunol. 2005, 12, 387–398. [Google Scholar] [CrossRef] [PubMed]
- Banks, M.C.; Kamel, N.S.; Zabriskie, J.B.; Larone, D.H.; Ursea, D.; Posnett, D.N. Staphylococcus aureus express unique superantigens depending on the tissue source. J. Infect. Dis. 2003, 187, 77–86. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Holtfreter, S.; Bauer, K.; Thomas, D.; Feig, C.; Lorenz, V.; Roschack, K.; Friebe, E.; Selleng, K.; Lövenich, S.; Greve, T.; et al. egc-encoded superantigens from Staphylococcus aureus are neutralized by human sera much less efficiently than are classical staphylococcal enterotoxins or toxic shock syndrome toxin. Infect. Immun. 2004, 72, 4061–4071. [Google Scholar] [CrossRef]
- Holtfreter, S.; Kolata, J.; Broker, B.M. Towards the immune proteome of Staphylococcus aureus—The anti-S. aureus antibody response. Int. J. Med. Microbiol. 2010, 300, 176–192. [Google Scholar] [CrossRef]
- Radke; E. E.; Brown, S.M.; Pelzek, A.J.; Fulmer, Y.; Hernandez, D.N.; Torres, V.J.; Thomsen, I.P.; Chiang, W.K.; Miller, A.O.; Shopsin, B.; et al. Hierarchy of human IgG recognition within the Staphylococcus aureus immunome. Sci. Rep. 2018, 8, 13296.
- Van Belkum, A.; Verkaik, N.; De Vogel, C.P.; Boelens, H.A.; Verveer, J.; Nouwen, J.L.; Verbrugh, H.A.; Wertheim, H.F.L. Reclassification of Staphylococcus aureus nasal carriage types. J. Infect. Dis. 2009, 199, 1820–1826. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Patient ID | Clonal Complex | Time Point | Sample Site | SNP Differences between C1 and C2 | SNP Differences Comparing C1 to C1 or C2 to C2 |
|---|---|---|---|---|---|
| 2 c | CC50 | C1 | N | Not applicable | 0 |
| 2 | CC50 | C1 | MS | ||
| 4 c | CC45 | C1 | N | Not applicable | 1 |
| 4 | CC45 | C1 | MS | ||
| 6 | CC30 | C1 | N | Not applicable | Not applicable |
| 7 c | CC8 | C1 | N | Not applicable | 2 |
| 7 | CC8 | C1 | MS | ||
| 8 | CC45 | C1 | N | 37,758–37,761 | 3 |
| 8 | CC50 | C2 | N | ||
| 8 | CC50 | C2 | MS | ||
| 9 | CC45 | C1 | N | 33,462–33,463 | 1 |
| 9 | CC30 | C2 | N | ||
| 9 | CC30 | C2 | MS | ||
| 11 | CC30 | C1 | N | 32,890–32,892 | 1 |
| 11 | CC30 | C1 | MS | ||
| 11 | CC50 | C2 | N | 3 | |
| 11 | CC50 | C2 | MS | ||
| 12 | CC45 | C1 | N | 29–30 a | 1 |
| 12 | CC45 | C2 | MS | ||
| 12 | CC45 | C2 | N | ||
| 15 | CC45 | C1 | N | 41,088–41,089 | 1 |
| 15 | CC45 | C1 | MS | ||
| 15 | CC15 | C2 | N | 2 | |
| 15 | CC15 | C2 | MS | ||
| 17 b | CC45 | C2 | N | Not applicable | 0 |
| 17 | CC45 | C2 | MS | ||
| 18 | CC30 | C1 | N | 65–67 a | 2 |
| 18 | CC30 | C1 | MS | ||
| 18 | CC30 | C2 | N | 0 | |
| 18 | CC30 | C2 | MS | ||
| 19 b | CC30 | C2 | N | Not applicable | 2 |
| 19 | CC30 | C2 | MS | ||
| 22 | CC9 | C1 | N | 18,896 | Not applicable |
| 22 | CC15 | C2 | N | ||
| 23 | CC59 | C1 | N | 32,884–32,885 | 3 |
| 23 | CC59 | C1 | MS | ||
| 23 | CC30 | C2 | N | ||
| 26 | CC59 | C2 | MS | Not applicable | Not applicable |
| 27 | CC45 | C1 | N | 33,500–33,505 | 1 |
| 27 | CC45 | C1 | MS | ||
| 27 | CC30 | C2 | N | 4 | |
| 27 | CC30 | C2 | MS | ||
| 31 | CC50 | C2 | N | Not applicable | Not applicable |
| 32 | CC1 | C1 | N | 13,939–17,065 | 15,884 |
| 32 | CC15 | C1 | MS | ||
| 32 | CC5 | C2 | N | ||
| 33 | CC8 | C1 | N | 48–51 a | 2 |
| 33 | CC8 | C1 | MS | ||
| 33 | CC8 | C2 | N | 5 | |
| 33 | CC8 | C2 | MS | ||
| 35 b | CC22 | C2 | N | Not applicable | 25 |
| 35 | CC22 | C2 | MS | ||
| 37 | CC182 | C1 | N | 29,807 | 0 |
| 37 | CC5 | C2 | N | ||
| 37 | CC5 | C2 | MS | ||
| 38 | CC30 | C1 | N | 817–818 | 2 |
| 38 | CC30 | C1 | MS | ||
| 38 | CC30 | C2 | N | 1 | |
| 38 | CC30 | C2 | MS | ||
| 39 | CC12 | C1 | MS | 12,182 | Not applicable |
| 39 | CC15 | C2 | N | ||
| 41 | CC22 | C1 | N | 33,882–33,883 | 1 |
| 41 | CC22 | C1 | MS | ||
| 41 | CC398 | C2 | N | ||
| 46 c | CC5 | C1 | N | Not applicable | 1 |
| 46 | CC5 | C1 | MS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thunberg, U.; Hugosson, S.; Ehricht, R.; Monecke, S.; Müller, E.; Cao, Y.; Stegger, M.; Söderquist, B. Long-Term Sinonasal Carriage of Staphylococcus aureus and Anti-Staphylococcal Humoral Immune Response in Patients with Chronic Rhinosinusitis. Microorganisms 2021, 9, 256. https://doi.org/10.3390/microorganisms9020256
Thunberg U, Hugosson S, Ehricht R, Monecke S, Müller E, Cao Y, Stegger M, Söderquist B. Long-Term Sinonasal Carriage of Staphylococcus aureus and Anti-Staphylococcal Humoral Immune Response in Patients with Chronic Rhinosinusitis. Microorganisms. 2021; 9(2):256. https://doi.org/10.3390/microorganisms9020256
Chicago/Turabian StyleThunberg, Ulrica, Svante Hugosson, Ralf Ehricht, Stefan Monecke, Elke Müller, Yang Cao, Marc Stegger, and Bo Söderquist. 2021. "Long-Term Sinonasal Carriage of Staphylococcus aureus and Anti-Staphylococcal Humoral Immune Response in Patients with Chronic Rhinosinusitis" Microorganisms 9, no. 2: 256. https://doi.org/10.3390/microorganisms9020256
APA StyleThunberg, U., Hugosson, S., Ehricht, R., Monecke, S., Müller, E., Cao, Y., Stegger, M., & Söderquist, B. (2021). Long-Term Sinonasal Carriage of Staphylococcus aureus and Anti-Staphylococcal Humoral Immune Response in Patients with Chronic Rhinosinusitis. Microorganisms, 9(2), 256. https://doi.org/10.3390/microorganisms9020256