
Quorum Sensing Regulation in Phytopathogenic Bacteria


Abstract
1. Introduction
2. Acyl Homoserine Lactone-Mediated Quorum Sensing
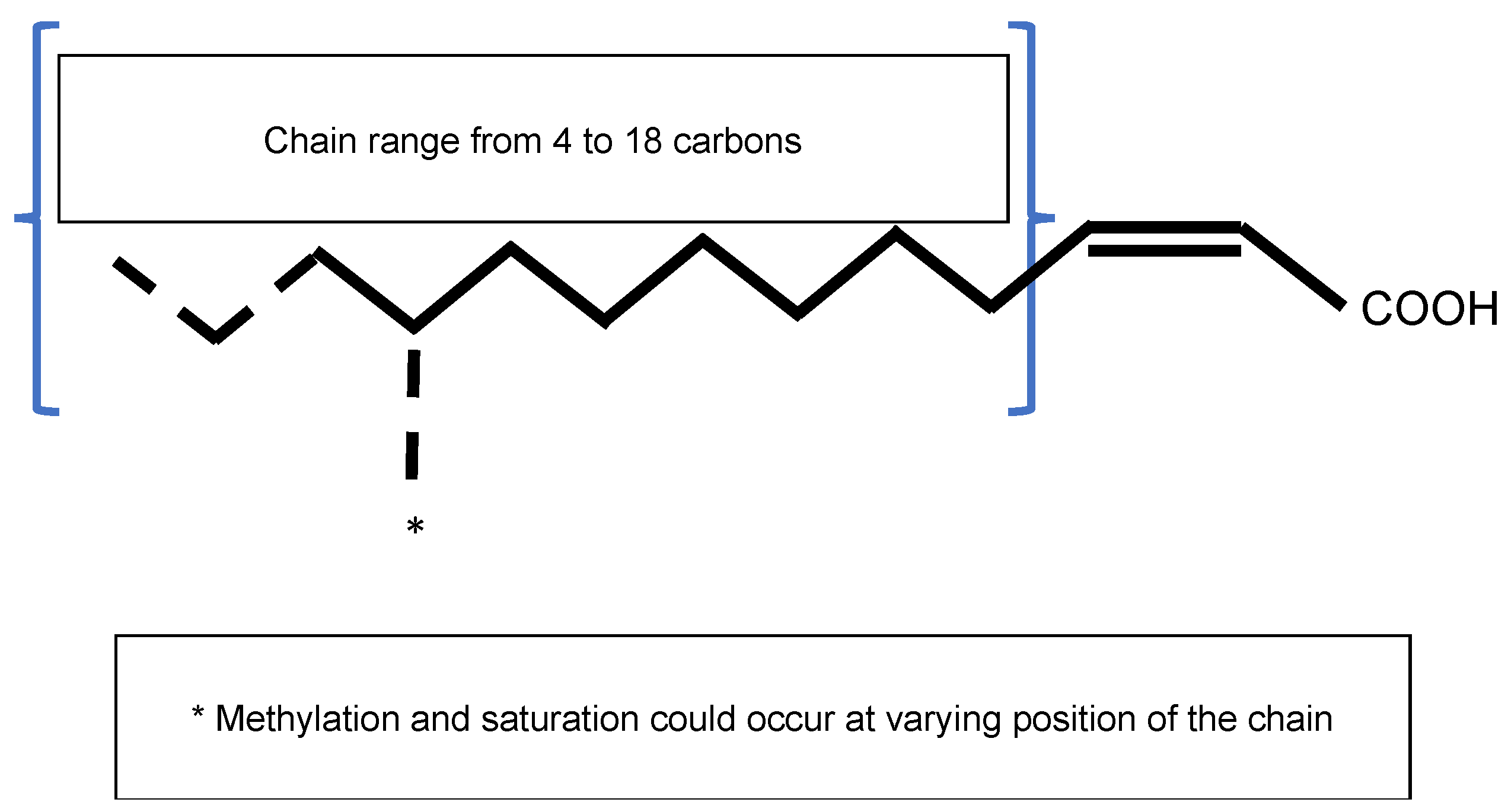
2.1. The Signal Molecule AHL
2.2. AHL-Mediated Quorum Sensing Overview
2.3. Functions Regulated by AHL-Mediated QS in Plant Pathogens
2.3.1. AHL-Mediated QS Is Important for Pseudomonas syringae Survival in Planta
2.3.2. AHL-Mediated QS Is Not Involved in Virulence Regulation in R. solanacearum
2.3.3. AHL-Mediated QS is Involved in the Conjugation of the Tumor-Inducing Plasmid in Agrobacterium spp.
2.3.4. The Involvement of AHL-Mediated QS in Virulence Is Questioned in Erwinia amylovora
2.3.5. The Involvement of AHL-Mediated QS in Virulence Is Questioned in Dickeya spp.
2.3.6. AHL-Mediated QS Is Involved in the Regulation of Virulence in Pectobacterium spp.
2.3.7. AHL-Mediated QS Negatively Impact Gene Expression in Pantoea stewartii
2.4. Induction, Maintenance, and Turnover of AHL-Mediated Quorum Sensing
2.4.1. Induction of AHL Synthesis
2.4.2. Turnover Mechanisms of QS: Quorum Quenching
3. Diffusible Signal Factor-Mediated Quorum Sensing
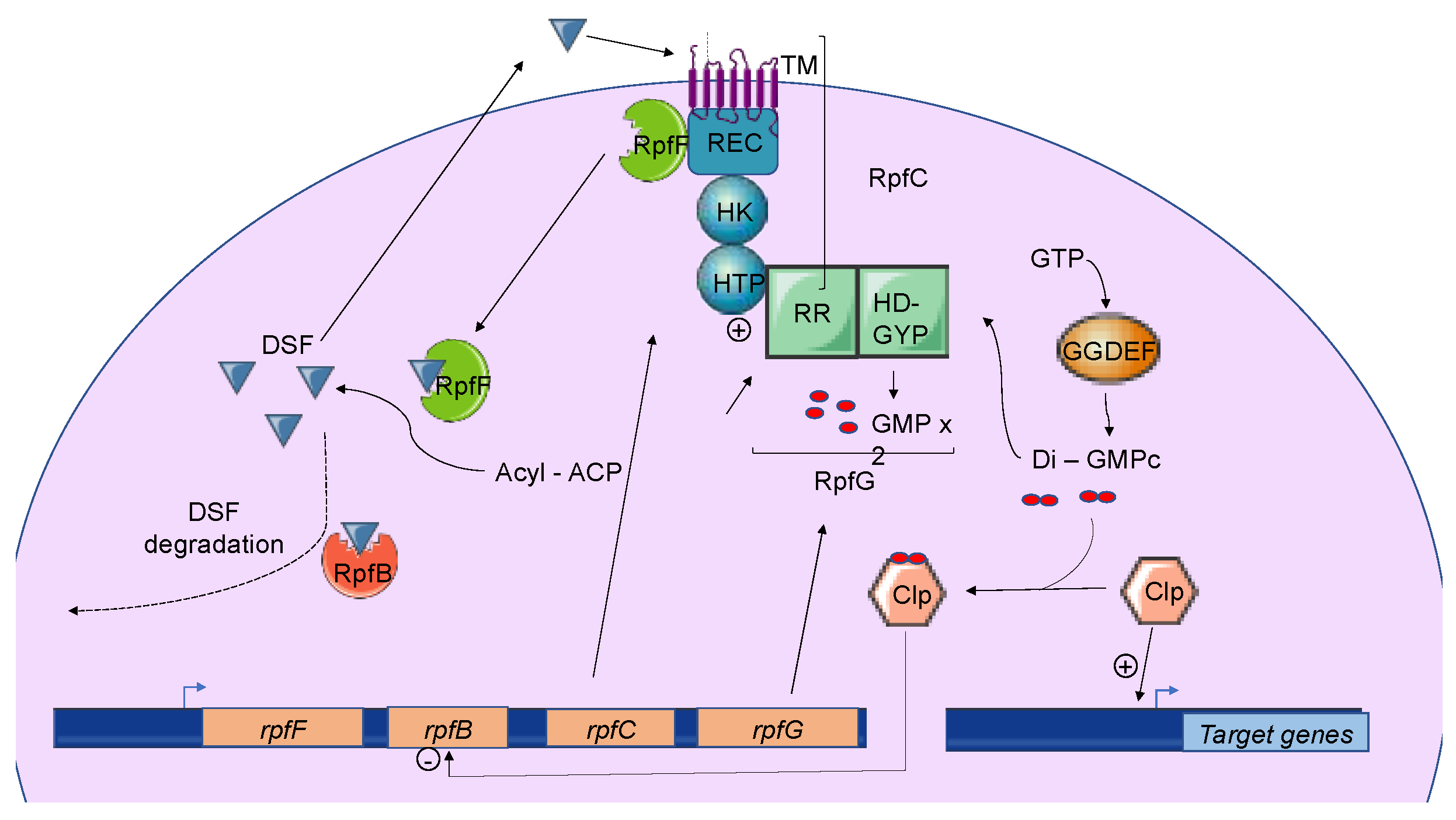
3.1. Overview of DSF-Mediated Quorum Sensing
3.2. Functions Regulated by DSF-Mediated Quorum Sensing in Plant Pathogens
3.2.1. Xanthomonas spp. Including X. oryzae, X. campestris, and X. axonopodis
3.2.2. In Xylella fastidiosa
3.2.3. Other DSF-Derived Signals
3.3. Induction, Maintenance, and Turnover of DSF-Mediated Quorum Sensing
3.3.1. Induction and Maintenance
3.3.2. Turnover of DSF Signals
4. The VFM Quorum Sensing System
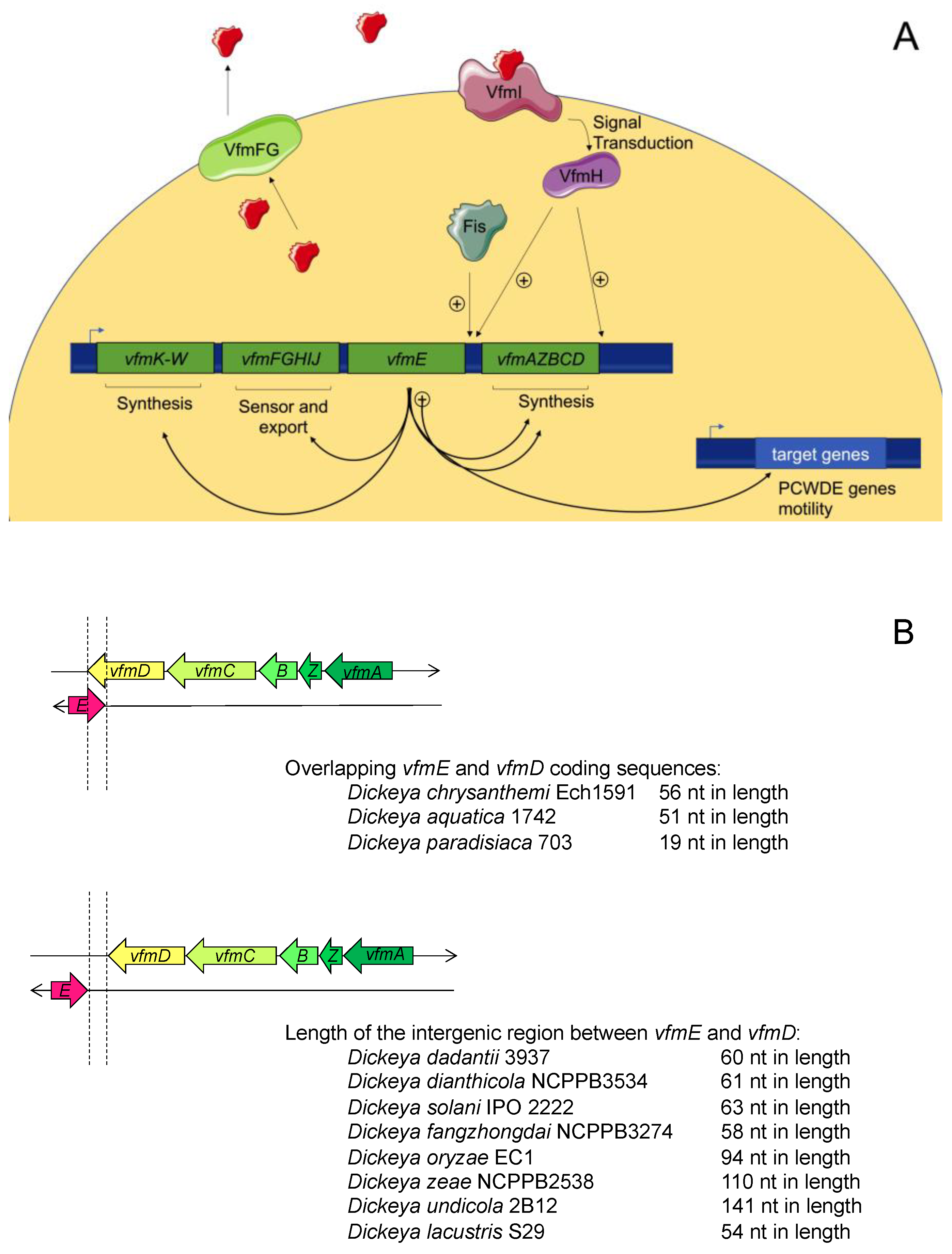
4.1. Overview of the VFM System
4.2. Regulation of the VFM System
5. Concluding Remarks and Future Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Eberhard, A.; Burlingame, A.L.; Kenyon, G.L.; Nealson, K.H.; Oppenheimer, N.J. Structural identification of autoinducer of Photobacterium fischeri luciferase. Biochemistry 1981, 20, 2444–2449. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.G.; Meighen, E. Purification and structural identification of an autoinducer for the luminescence system of Vibrio harveyi. J. Biol. Chem. 1989, 264, 21670–21676. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal–response systems in Gram-negative bacteria. Nat. Rev. Genet. 2016, 14, 576–588. [Google Scholar] [CrossRef] [PubMed]
- Bainton, N.J.; Stead, P.; Chhabra, S.R.; Bycroft, B.W.; Salmond, G.P.C.; Stewart, G.S.A.B.; Williams, P. N-(3-oxohexanoyl)-l-homoserine lactone regulates carbapenem antibiotic production in Erwinia carotovora. Biochem. J. 1992, 288, 997–1004. [Google Scholar] [CrossRef] [PubMed]
- Williams, P. Quorum sensing, communication and cross-kingdom signaling in the bacterial world. Microbiology 2007, 153, 3923–3938. [Google Scholar] [CrossRef]
- Von Bodman, S.B.; Bauer, W.D.; Coplin, D.L. Quorum sensing in plant-pathogenic bacteria. Annu. Rev. Phytopathol. 2003, 41, 455–482. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef]
- Chhabra, S.R.; Philipp, B.; Eberl, L.; Givskov, M.; Williams, P.; Cámara, M. Extracellular communication in bacteria. In Topics in Current Chemistry; Springer Nature: Berlin/Heidelberg, Germany, 2004; pp. 279–315. [Google Scholar]
- Yates, E.A.; Philipp, B.; Williams, P.; Buckley, C.; Atkinson, S.; Chhabra, S.R.; Sockett, R.E.; Goldner, M.; Dessaux, Y.; Cámara, M.; et al. N-acylhomoserine lactones undergo lactonolysis in a pH-, temperature-, and acyl chain length-dependent manner during growth of Yersinia pseudotuberculosis and Pseudomonas aeruginosa. Infect. Immun. 2002, 70, 5635–5646. [Google Scholar] [CrossRef]
- Parsek, M.R.; Val, D.L.; Hanzelka, B.L.; Cronan, J.E.; Greenberg, E.P. Acyl homoserine-lactone quorum-sensing signal generation. Proc. Natl. Acad. Sci. USA 1999, 96, 4360–4365. [Google Scholar] [CrossRef]
- Urbanowski, M.L.; Lostroh, C.P.; Greenberg, E.P. Reversible acyl-homoserine lactone binding to purified Vibrio fischeri LuxR protein. J. Bacteriol. 2004, 186, 631–637. [Google Scholar] [CrossRef]
- Zhang, R.-G.; Pappas, K.M.; Brace, J.L.; Miller, P.C.; Oulmassov, T.; Molyneaux, J.M.; Anderson, J.C.; Bashkin, J.K.; Winans, S.C.; Joachimiak, A. Structure of a bacterial quorum-sensing transcription factor complexed with pheromone and DNA. Nat. Cell Biol. 2002, 417, 971–974. [Google Scholar] [CrossRef] [PubMed]
- Whitehead, N.A.; Barnard, A.M.L.; Slater, H.; Simpson, N.J.L.; Salmond, G.P.C. Quorum-sensing in Gram-negative bacteria. FEMS Microbiol. Rev. 2001, 25, 365–404. [Google Scholar] [CrossRef] [PubMed]
- Hudaiberdiev, S.; Choudhary, K.S.; Alvarez, R.V.; Gelencsér, Z.; Ligeti, B.; Lamba, D.; Pongor, S. Census of solo luxR genes in prokaryotic genomes. Front. Cell. Infect. Microbiol. 2015, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, B.; Pujol, C.J.; Lindow, S.E. Regulation of AHL production and its contribution to epiphytic fitness in Pseudomonas syringae. Mol. Plant Microbe Interact. 2004, 17, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Quiñones, B.; Dulla, G.; Lindow, S.E. Quorum sensing regulates exopolysaccharide production, motility, and virulence in Pseudomonas syringae. Mol. Plant Microbe Interact. 2005, 18, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Ma, A.; Zhuang, X.; He, X.; Zhuang, G. N-(3-oxo-hexanoyl)-homoserine lactone has a critical contribution to the quorum-sensing-dependent regulation in phytopathogen Pseudomonas syringae pv. tabaci 11528. FEMS Microbiol. Lett. 2016, 363, fnw265. [Google Scholar] [CrossRef]
- Cheng, F.; Ma, A.; Luo, J.; Zhuang, X.; Zhuang, G. N-acylhomoserine lactone-regulation of genes mediating motility and pathogenicity in Pseudomonas syringae pathovar tabaci 11528. Microbiol. Open 2017, 6, e00440. [Google Scholar] [CrossRef]
- Patel, H.K.; Ferrante, P.; Covaceuszach, S.; Lamba, D.; Scortichini, M.; Venturi, V. The kiwifruit emerging pathogen Pseudomonas syringae pv. actinidiae does not produce AHLs but possesses three LuxR solos. PLoS ONE 2014, 9, e87862. [Google Scholar] [CrossRef]
- Cellini, A.; Donati, I.; Fiorentini, L.; Vandelle, E.; Polverari, A.; Venturi, V.; Buriani, G.; Vanneste, J.L.; Spinelli, F. N-acyl homoserine lactones and LuxR solos regulate social behaviour and virulence of Pseudomonas syringae pv. actinidiae. Microb. Ecol. 2020, 79, 383–396. [Google Scholar] [CrossRef]
- Flavier, A.B.; Ganova-Raeva, L.M.; Schell, M.; Denny, T.P. Hierarchical autoinduction in Ralstonia solanacearum: Control of acyl-homoserine lactone production by a novel autoregulatory system responsive to 3-hydroxypalmitic acid methyl ester. J. Bacteriol. 1997, 179, 7089–7097. [Google Scholar] [CrossRef]
- Chen, C.-N.; Chen, C.-J.; Liao, C.-T.; Lee, C.-Y. A probable aculeacin A acylase from the Ralstonia solanacearum GMI1000 is N-acyl-homoserine lactone acylase with quorum-quenching activity. BMC Microbiol. 2009, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Burke, A.K.; Duong, D.A.; Jensen, R.V.; Stevens, A.M. Analyzing the transcriptomes of two quorum-sensing controlled transcription factors, RcsA and LrhA, important for Pantoea stewartii virulence. PLoS ONE 2015, 10, e0145358. [Google Scholar] [CrossRef]
- Lang, J.; Faure, D. Functions and regulation of quorum-sensing in Agrobacterium tumefaciens. Front. Plant Sci. 2014, 5, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Murphy, P.J.; Kerr, A.; Tate, M.E. Agrobacterium conjugation and gene regulation by N-acyl-L-homoserine lactones. Nat. Cell Biol. 1993, 362, 446–448. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Winans, S.C. TraA, TraC and TraD autorepress two divergent quorum-regulated promoters near the transfer origin of the Ti plasmid of Agrobacterium tumefaciens. Mol. Microbiol. 2007, 63, 1769–1782. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Khan, S.R.; Farrand, S.K. Induction and loss of Ti plasmid conjugative competence in response to the acyl-homoserine lactone quorum-sensing signal. J. Bacteriol. 2008, 190, 4398–4407. [Google Scholar] [CrossRef]
- Li, P.L.; Everhart, D.M.; Farrand, S.K. Genetic and sequence analysis of the pTiC58 Trb locus, encoding a mating-pair formation system related to members of the type IV secretion family. J. Bacteriol. 1998, 180, 6164–6172. [Google Scholar] [CrossRef]
- Hwang, I.; Li, P.L.; Zhang, L.; Piper, K.R.; Cook, D.M.; Tate, M.E.; Farrand, S.K. TraI, a LuxI homologue, is responsible for production of conjugation factor, the Ti plasmid N-acylhomoserine lactone autoinducer. Proc. Natl. Acad. Sci. USA 1994, 91, 4639–4643. [Google Scholar] [CrossRef]
- Piqué, N.; Miñana-Galbis, D.; Merino, S.; Tomás, J.M. Virulence factors of Erwinia amylovora: A review. Int. J. Mol. Sci. 2015, 16, 12836–12854. [Google Scholar] [CrossRef]
- Venturi, V.; Venuti, C.; Devescovi, G.; Lucchese, C.; Friscina, A.; Degrassi, G.; Aguilar, C.; Mazzucchi, U. The plant pathogen Erwinia amylovora produces acyl-homoserine lactone signal molecules in vitro and in planta. FEMS Microbiol. Lett. 2004, 241, 179–183. [Google Scholar] [CrossRef]
- Molina, L.; Rezzonico, F.; Défago, G.; Duffy, B. Autoinduction in Erwinia amylovora: Evidence of an acyl-homoserine lactone signal in the fire blight pathogen. J. Bacteriol. 2005, 187, 3206–3213. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, M.; Geider, K. Autoinducer-2 of the fire blight pathogen Erwinia amylovora and other plant-associated bacteria. FEMS Microbiol. Lett. 2007, 266, 34–41. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rezzonico, F.; Duffy, B. The role of LuxS in the fire blight pathogen Erwinia amylovora is limited to metabolism and does not involve quorum sensing. Mol. Plant-Microbe Interactions 2007, 20, 1284–1297. [Google Scholar] [CrossRef]
- Nasser, W.; Bouillant, M.L.; Salmond, G.; Reverchon, S. Characterization of the Erwinia chrysanthemi expI–expR locus directing the synthesis of two N-acyl-homoserine lactone signal molecules. Mol. Microbiol. 1998, 29, 1391–1405. [Google Scholar] [CrossRef] [PubMed]
- Castang, S.; Reverchon, S.; Gouet, P.; Nasser, W. Direct evidence for the modulation of the activity of the Erwinia chrysanthemi quorum-sensing regulator ExpR by acylhomoserine lactone pheromone. J. Biol. Chem. 2006, 281, 29972–29987. [Google Scholar] [CrossRef] [PubMed]
- Reverchon, S.; Bouillant, M.L.; Salmond, G.; Nasser, W. Integration of the quorum-sensing system in the regulatory networks controlling virulence factor synthesis in Erwinia chrysanthemi. Mol. Microbiol. 1998, 29, 1407–1418. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.B.B.M.; Zhang, H.-B.; Xu, J.-L.; Liu, Q.; Jiang, Z.; Zhang, L.-H. The acyl-homoserine lactone-type quorum-sensing system modulates cell motility and virulence of Erwinia chrysanthemi pv. zeae. J. Bacteriol. 2007, 190, 1045–1053. [Google Scholar] [CrossRef]
- Potrykus, M.; Hugouvieux-Cotte-Pattat, N.; Lojkowska, E. Interplay of classic Exp and specific Vfm quorum sensing systems on the phenotypic features of Dickeya solani strains exhibiting different virulence levels. Mol. Plant Pathol. 2017, 19, 1238–1251. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Chatterjee, A.; Hasegawa, H.; Dixit, V.; Leigh, N.; Chatterjee, A.K. ExpR, a LuxR homolog of Erwinia carotovora subsp. carotovora, activates transcription of rsmA, which specifies a global regulatory RNA-binding protein. J. Bacteriol. 2005, 187, 4792–4803. [Google Scholar] [CrossRef]
- Põllumaa, L.; Alamäe, T.; Mäe, A. Quorum sensing and expression of virulence in Pectobacteria. Sensors 2012, 12, 3327–3349. [Google Scholar] [CrossRef] [PubMed]
- McGowan, S.; Sebaihia, M.; Jones, S.; Yu, B.; Bainton, N.; Chan, P.F.; Bycroft, B.; Stewart, G.; Williams, P.; Salmond, G.P.C. Carbapenem antibiotic production in Erwinia carotovora is regulated by CarR, a homologue of the LuxR transcriptional activator. Microbiology 1995, 141, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Yu, B.; Bainton, N.; Birdsall, M.; Bycroft, B.; Chhabra, S.; Cox, A.; Golby, P.; Reeves, P.; Stephens, S. The lux autoinducer regulates the production of exoenzyme virulence determinants in Erwinia carotovora and Pseudomonas aeruginosa. EMBO J. 1993, 12, 2477–2482. [Google Scholar] [CrossRef]
- Liu, H.; Coulthurst, S.J.; Salmond, G.P.C.; Toth, I.K.; Pritchard, L.; Hedley, P.E.; Ravensdale, M.; Humphris, S.; Burr, T.; Takle, G.; et al. Quorum sensing coordinates brute force and stealth modes of infection in the plant pathogen Pectobacterium atrosepticum. PLoS Pathog. 2008, 4, e1000093. [Google Scholar] [CrossRef] [PubMed]
- Patel, H.K.; Suarezmoreno, Z.R.; Degrassi, G.; Subramoni, S.; Gonzalez, J.F.; Venturi, V. Bacterial LuxR solos have evolved to respond to different molecules including signals from plants. Front. Plant Sci. 2013, 4, 447. [Google Scholar] [CrossRef]
- Von Bodman, S.B.; Majerczak, D.R.; Coplin, D.L. A negative regulator mediates quorum-sensing control of exopolysaccharide production in Pantoea stewartii subsp. stewartii. Proc. Natl. Acad. Sci. USA 1998, 95, 7687–7692. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.; Babujee, L.; Jacobs, J.M.; Allen, C. Comparative transcriptome analysis reveals cool virulence factors of Ralstonia solanacearum race 3 biovar 2. PLoS ONE 2015, 10, e0139090. [Google Scholar] [CrossRef]
- Kai, K.; Ohnishi, H.; Shimatani, M.; Ishikawa, S.; Mori, Y.; Kiba, A.; Ohnishi, K.; Tabuchi, M.; Hikichi, Y. Methyl 3-hydroxymyristate, a diffusible signal mediating phc quorum sensing in Ralstonia solanacearum. ChemBioChem 2015, 16, 2309–2318. [Google Scholar] [CrossRef]
- Ujita, Y.; Sakata, M.; Yoshihara, A.; Hikichi, Y.; Kai, K. Signal production and response specificity in the phc quorum sensing systems of Ralstonia solanacearum species complex. ACS Chem. Biol. 2019, 14, 2243–2251. [Google Scholar] [CrossRef] [PubMed]
- Gelvin, S.B. Traversing the cell: Agrobacterium T-DNA’s journey to the host genome. Front. Plant Sci. 2012, 3, 52. [Google Scholar] [CrossRef]
- Bourras, S.; Rouxel, T.; Meyer, M. Agrobacterium tumefaciens gene transfer: How a plant pathogen hacks the nuclei of plant and non plant organisms. Phytopathology 2015, 105, 1288–1301. [Google Scholar] [CrossRef]
- Dessaux, Y.; Petit, A.; Farrand, S.K.; Murphy, P.J. Opines and opine-like molecules involved in plant-Rhizobiaceae interactions. In The Rhizobiaceae: Molecular and Biology of Model Plant-Associated bacteria; Spaink, H.P., Kondorosi, A., Hooykaas, P.J., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Boston, MA, USA; London, UK, 1998; pp. 173–197. [Google Scholar]
- Samson, R.; Legendre, J.B.; Christen, R.; Saux, M.F.-L.; Achouak, W.; Gardan, L. Transfer of Pectobacterium chrysanthemi (Burkholder et al. 1953) Brenner et al. 1973 and Brenneria paradisiaca to the genus Dickeya gen. nov. as Dickeya chrysanthemi comb. nov. and Dickeya paradisiaca comb. nov. and delineation of four novel species, Dickeya dadantii sp. nov., Dickeya dianthicola sp. nov., Dickeya dieffenbachiae sp. nov. and Dickeya zeae sp. nov. Int. J. Syst. Evol. Microbiol. 2005, 55, 1415–1427. [Google Scholar] [CrossRef] [PubMed]
- Brady, C.L.; Cleenwerck, I.; Denman, S.; Venter, S.N.; Rodríguez-Palenzuela, P.; Coutinho, T.A.; De Vos, P. Proposal to reclassify Brenneria quercina (Hildebrand and Schroth 1967) Hauben et al. 1999 into a new genus, Lonsdalea gen. nov., as Lonsdalea quercina comb. nov., descriptions of Lonsdalea quercina subsp. quercina comb. nov., Lonsdalea quercina subsp. iberica subsp. nov. and Lonsdalea quercina subsp. britannica subsp. nov., emendation of the description of the genus Brenneria, reclassification of Dickeya dieffenbachiae as Dickeya dadantii subsp. dieffenbachiae comb. nov., and emendation of the description of Dickeya dadantii. Int. J. Syst. Evol. Microbiol. 2012, 62, 1592–1602. [Google Scholar] [CrossRef] [PubMed]
- Van Der Wolf, J.M.; Nijhuis, E.H.; Kowalewska, M.J.; Saddler, G.S.; Parkinson, N.; Elphinstone, J.G.; Pritchard, L.; Toth, I.K.; Lojkowska, E.; Potrykus, M.; et al. Dickeya solani sp. nov., a pectinolytic plant-pathogenic bacterium isolated from potato (Solanum tuberosum). Int. J. Syst. Evol. Microbiol. 2014, 64, 768–774. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Zhao, Y.; Yuan, X.; Yi, J.; Fan, J.; Xu, Z.; Hu, B.; De Boer, S.H.; Li, X. Dickeya fangzhongdai sp. nov., a plant-pathogenic bacterium isolated from pear trees (Pyrus pyrifolia). Int. J. Syst. Evol. Microbiol. 2016, 66, 2831–2835. [Google Scholar] [CrossRef] [PubMed]
- Hugouvieux-Cotte-Pattat, N.; Jacot-Des-Combes, C.; Briolay, J. Dickeya lacustris sp. nov., a water-living pectinolytic bacterium isolated from lakes in France. Int. J. Syst. Evol. Microbiol. 2019, 69, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Hugouvieux-Cotte-Pattat, N.; Brochier-Armanet, C.; Flandrois, J.-P.; Reverchon, S. Dickeya poaceiphila sp. nov., a plant-pathogenic bacterium isolated from sugar cane (Saccharum officinarum). Int. J. Syst. Evol. Microbiol. 2020, 70, 4508–4514. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; He, S.-W.; Guo, H.-B.; Han, J.-G.; Thin, K.K.; Gao, J.-S.; Wang, Y.; Zhang, X.-X. Dickeya oryzae sp. nov., isolated from the roots of rice. Int. J. Syst. Evol. Microbiol. 2020, 70, 4171–4178. [Google Scholar] [CrossRef]
- Feng, L.; Schaefer, A.L.; Hu, M.; Chen, R.; Greenberg, E.P.; Zhou, J. Virulence factor identification in the banana pathogen Dickeya zeae ms2. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef]
- Ham, J.H.; Cui, Y.; Alfano, J.R.; Rodríguez-Palenzuela, P.; Rojas, C.M.; Chatterjee, A.K.; Collmer, A. Analysis of Erwinia chrysanthemi EC16 pelE∷uidA, pelL∷uida, and hrpN∷uidA mutants reveals strain-specific atypical regulation of the Hrp type III secretion system. Mol. Plant Microbe Interact. 2004, 17, 184–194. [Google Scholar] [CrossRef]
- Nasser, W.; Dorel, C.; Wawrzyniak, J.; Van Gijsegem, F.; Groleau, M.-C.; Déziel, E.; Reverchon, S. Vfm a new quorum sensing system controls the virulence of Dickeya dadantii. Environ. Microbiol. 2012, 15, 865–880. [Google Scholar] [CrossRef]
- Genome-Based Phylogeny and Taxonomy of the “Enterobacteriales”: Proposal for Enterobacterales ord. nov. divided into the Families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov-PubMed. Available online: https://pubmed-ncbi-nlm-nih-gov.insb.bib.cnrs.fr/27620848/ (accessed on 1 January 2021).
- Von Bodman, S.B.; Ball, J.K.; Faini, M.A.; Herrera, C.M.; Minogue, T.D.; Urbanowski, M.L.; Stevens, A.M. The quorum sensing negative regulators EsaR and ExpREcc, homologues within the LuxR family, retain the ability to function as activators of transcription. J. Bacteriol. 2003, 185, 7001–7007. [Google Scholar] [CrossRef] [PubMed]
- Dumenyo, C.; Mukherjee, A.; Chun, W.; Chatterjee, A.K. Genetic and physiological evidence for the production of N-acyl homoserine lactones by Pseudomonas syringae pv. syringae and other fluorescent plant pathogenic Pseudomonas species. Eur. J. Plant Pathol. 1998, 104, 569–582. [Google Scholar] [CrossRef]
- Marutani, M.; Taguchi, F.; Ogawa, Y.; Hossain, M.; Inagaki, Y.; Toyoda, K.; Shiraishi, T.; Ichinose, Y. Gac two-component system in Pseudomonas syringae pv. tabaci is required for virulence but not for hypersensitive reaction. Mol. Genet. Genom. 2007, 279, 313–322. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chatterjee, A.; Cui, Y.; Yang, H.; Collmer, A.; Alfano, J.R.; Chatterjee, A.K. GacA, the response regulator of a two-component system, acts as a master regulator in Pseudomonas syringae pv. tomato DC3000 by controlling regulatory RNA, transcriptional activators, and alternate sigma factors. Mol. Plant Microbe Interact. 2003, 16, 1106–1117. [Google Scholar] [CrossRef]
- Sawada, T.; Eguchi, M.; Asaki, S.; Kashiwagi, R.; Shimomura, K.; Taguchi, F.; Matsui, H.; Yamamoto, M.; Noutoshi, Y.; Toyoda, K.; et al. MexEF-OprN multidrug efflux pump transporter negatively controls N-acyl-homoserine lactone accumulation in Pseudomonas syringae pv. tabaci 6605. Mol. Genet. Genom. 2018, 293, 907–917. [Google Scholar] [CrossRef]
- Piper, K.R.; Von Bodman, S.B.; Farrand, S.K. Conjugation factor of Agrobacterium tumefaciens regulates Ti plasmid transfer by autoinduction. Nat. Cell Biol. 1993, 362, 448–450. [Google Scholar] [CrossRef]
- Hwang, I.; Cook, D.M.; Farrand, S.K. A new regulatory element modulates homoserine lactone-mediated autoinduction of Ti plasmid conjugal transfer. J. Bacteriol. 1995, 177, 449–458. [Google Scholar] [CrossRef]
- Zhu, J.; Winans, S.C. Autoinducer binding by the quorum-sensing regulator TraR increases affinity for target promoters in vitro and decreases TraR turnover rates in whole cells. Proc. Natl. Acad. Sci. USA 1999, 96, 4832–4837. [Google Scholar] [CrossRef]
- Diel, B.; Dequivre, M.; Wisniewski-Dyé, F.; Vial, L.; Hommais, F. A novel plasmid-transcribed regulatory sRNA, QfsR, controls chromosomal polycistronic gene expression in Agrobacterium fabrum. Environ. Microbiol. 2019, 21, 3063–3075. [Google Scholar] [CrossRef]
- Shepherd, R.W.; Lindow, S.E. Two dissimilar N-acyl-homoserine lactone acylases of Pseudomonas syringae influence colony and biofilm morphology. Appl. Environ. Microbiol. 2008, 75, 45–53. [Google Scholar] [CrossRef]
- Haudecoeur, E.; Tannières, M.; Cirou, A.; Raffoux, A.; Dessaux, Y.; Faure, D. Different regulation and roles of lactonases AiiB and AttM in Agrobacterium tumefaciens C58. Mol. Plant Microbe Interact. 2009, 22, 529–537. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.D.; Chai, Y.; Winans, S.C. The quorum-sensing protein TraR of Agrobacterium tumefaciens is susceptible to intrinsic and TraM-mediated proteolytic instability. Mol. Microbiol. 2012, 84, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wu, J.; Tao, F.; Zhang, L.-H. Listening to a new language: DSF-based quorum sensing in Gram-negative bacteria. Chem. Rev. 2011, 111, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, L.-H.; Cámara, M.; He, Y. The DSF family of quorum sensing signals: Diversity, biosynthesis, and turnover. Trends Microbiol. 2017, 25, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Almeida, R.P.P.; Killiny, N.; Newman, K.L.; Chatterjee, S.; Ionescu, M.; Lindow, S.E. Contribution of RpfB to cell-to-cell signal synthesis, virulence, and vector transmission of Xylella fastidiosa. Mol. Plant Microbe Interact. 2012, 25, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, M.; Yokota, K.; Antonova, E.; Garcia, A.; Beaulieu, E.; Hayes, T.; Iavarone, A.T.; Lindow, S.E. Promiscuous diffusible signal factor production and responsiveness of the Xylella fastidiosa Rpf system. mBio 2016, 7, e01054-16. [Google Scholar] [CrossRef] [PubMed]
- Slater, H.; Alvarez-Morales, A.; Barber, C.E.; Daniels, M.J.; Dow, M. A two-component system involving an HD-GYP domain protein links cell-cell signalling to pathogenicity gene expression in Xanthomonas campestris. Mol. Microbiol. 2002, 38, 986–1003. [Google Scholar] [CrossRef]
- An, S.-Q.; Potnis, N.; Dow, M.; Vorhölter, F.-J.; He, Y.-Q.; Becker, A.; Teper, D.; Li, Y.; Wang, N.; Bleris, L.; et al. Mechanistic insights into host adaptation, virulence and epidemiology of the phytopathogen Xanthomonas. FEMS Microbiol. Rev. 2019, 44, 1–32. [Google Scholar] [CrossRef]
- Timilsina, S.; Potnis, N.; Newberry, E.A.; Liyanapathiranage, P.; Iruegas-Bocardo, F.; White, F.F.; Goss, E.M.; Jones, J.B. Xanthomonas diversity, virulence and plant–pathogen interactions. Nat. Rev. Genet. 2020, 18, 415–427. [Google Scholar] [CrossRef]
- He, Y.; Xu, M.; Lin, K.; Ng, Y.-J.A.; Wen, C.-M.; Wang, L.-H.; Liu, Z.-D.; Zhang, H.-B.; Dong, Y.-H.; Dow, J.M.; et al. Genome scale analysis of diffusible signal factor regulon in Xanthomonas campestris pv. campestris: Identification of novel cell-cell communication-dependent genes and functions. Mol. Microbiol. 2005, 59, 610–622. [Google Scholar] [CrossRef]
- Gudesblat, G.E.; Torres, P.S.; Vojnov, A. Xanthomonas campestris overcomes Arabidopsis stomatal innate immunity through a DSF cell-to-cell signal-regulated virulence factor. Plant Physiol. 2008, 149, 1017–1027. [Google Scholar] [CrossRef] [PubMed]
- Kakkar, A.; Nizampatnam, N.R.; Kondreddy, A.; Pradhan, B.B.; Chatterjee, S. Xanthomonas campestris cell-cell signalling molecule DSF (diffusible signal factor) elicits innate immunity in plants and is suppressed by the exopolysaccharide xanthan. J. Exp. Bot. 2015, 66, 6697–6714. [Google Scholar] [CrossRef] [PubMed]
- He, Y.-W.; Wu, J.; Cha, J.-S.; Zhang, L.-H. Rice bacterial blight pathogen Xanthomonas oryzae pv. oryzae produces multiple DSF-family signals in regulation of virulence factor production. BMC Microbiol. 2010, 10, 187. [Google Scholar] [CrossRef]
- Thowthampitak, J.; Shaffer, B.T.; Prathuangwong, S.; Loper, J.E. Role of RpfF in virulence and exoenzyme production of Xanthomonas axonopodis pv. glycines, the causal agent of bacterial pustule of soybean. Phytopathology 2008, 98, 1252–1260. [Google Scholar] [CrossRef] [PubMed]
- Roper, M.C.; Castro, C.; Ingel, B. Xylella fastidiosa: Bacterial parasitism with hallmarks of commensalism. Curr. Opin. Plant Biol. 2019, 50, 140–147. [Google Scholar] [CrossRef]
- Chatterjee, S.; Wistrom, C.; Lindow, S.E. A cell-cell signaling sensor is required for virulence and insect transmission of Xylella fastidiosa. Proc. Natl. Acad. Sci. USA 2008, 105, 2670–2675. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Yu, Y.; Chen, X.; Diab, A.A.; Ruan, L.; He, J.; Wang, H.; He, Y. The multiple DSF-family QS signals are synthesized from carbohydrate and branched-chain amino acids via the FAS elongation cycle. Sci. Rep. 2015, 5, srep13294. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wu, J.; Yin, W.; Li, P.; Zhou, J.; Chen, S.; He, F.; Cai, J.; Zhang, L.-H. Diffusible signal factor family signals provide a fitness advantage to Xanthomonas campestris pv. campestris in interspecies competition. Environ. Microbiol. 2016, 18, 1534–1545. [Google Scholar] [CrossRef]
- Torres, P.S.; Malamud, F.; Rigano, L.A.; Russo, D.M.; Marano, M.R.; Castagnaro, A.P.; Zorreguieta, A.; Bouarab, K.; Dow, M.A.; Vojnov, A. Controlled synthesis of the DSF cell-cell signal is required for biofilm formation and virulence in Xanthomonas campestris. Environ. Microbiol. 2007, 9, 2101–2109. [Google Scholar] [CrossRef]
- Andrade, M.O.; Alegria, M.C.; Guzzo, C.R.; Docena, C.; Rosa, M.C.P.; Ramos, C.H.I.; Farah, C.S. The HD-GYP domain of RpfG mediates a direct linkage between the Rpf quorum-sensing pathway and a subset of diguanylate cyclase proteins in the phytopathogen Xanthomonas axonopodis pv citri. Mol. Microbiol. 2006, 62, 537–551. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, X.-Y.; Sun, S.; Yang, L.-C.; Jiang, B.-L.; He, Y. Identification and characterization of naturally occurring DSF-family quorum sensing signal turnover system in the phytopathogen Xanthomonas. Environ. Microbiol. 2015, 17, 4646–4658. [Google Scholar] [CrossRef] [PubMed]
- He, Y.-W.; Zhang, L.-H. Quorum sensing and virulence regulation in Xanthomonas campestris. FEMS Microbiol. Rev. 2008, 32, 842–857. [Google Scholar] [CrossRef] [PubMed]
- Ionescu, M.; Zaini, P.A.; Baccari, C.; Tran, S.; Da Silva, A.M.; Lindow, S.E. Xylella fastidiosa outer membrane vesicles modulate plant colonization by blocking attachment to surfaces. Proc. Natl. Acad. Sci. USA 2014, 111, E3910–E3918. [Google Scholar] [CrossRef] [PubMed]
- Lowe-Power, T.M.; Khokhani, D.; Allen, C. How Ralstonia solanacearum exploits and thrives in the flowing plant xylem environment. Trends Microbiol. 2018, 26, 929–942. [Google Scholar] [CrossRef]
- Flavier, A.B.; Clough, S.J.; Schell, M.A.; Denny, T.P. Identification of 3-hydroxypalmitic acid methyl ester as a novel autoregulator controlling virulence in Ralstonia solanacearum. Mol. Microbiol. 1997, 26, 251–259. [Google Scholar] [CrossRef]
- Barber, C.E.; Tang, J.L.; Feng, J.X.; Pan, M.Q.; Wilson, T.J.G.; Slater, H.; Dow, J.M.; Williams, P.; Daniels, M.J. A novel regulatory system required for pathogenicity of Xanthomonas campestris is mediated by a small diffusible signal molecule. Mol. Microbiol. 1997, 24, 555–566. [Google Scholar] [CrossRef]
- Bi, H.; Yu, Y.; Dong, H.; Wang, H.; Cronan, J.E. Xanthomonas campestris RpfB is a fatty Acyl-CoA ligase required to counteract the thioesterase activity of the RpfF diffusible signal factor (DSF) synthase. Mol. Microbiol. 2014, 93, 262–275. [Google Scholar] [CrossRef]
- Espinosa-Urgel, M. Learning when (and how) to shut up: Intercellular signal turnover in Xanthomonas. Environ. Microbiol. 2016, 18, 314–315. [Google Scholar] [CrossRef]
- Duprey, A.; Taib, N.; Leonard, S.; Garin, T.; Flandrois, J.; Nasser, W.; Brochier-Armanet, C.; Reverchon, S. The phytopathogenic nature of Dickeya aquatica 174/2 and the dynamic early evolution of Dickeya pathogenicity. Environ. Microbiol. 2019, 21, 2809–2835. [Google Scholar] [CrossRef]
- Hommais, F.; Oger-Desfeux, C.; Van Gijsegem, F.; Castang, S.; Ligori, S.; Expert, D.; Nasser, W.; Reverchon, S. PecS is a global regulator of the symptomatic phase in the phytopathogenic bacterium Erwinia chrysanthemi 3937. J. Bacteriol. 2008, 190, 7508–7522. [Google Scholar] [CrossRef]
- Leonard, S.; Hommais, F.; Nasser, W.; Reverchon, S. Plant-phytopathogen interactions: Bacterial responses to environmental and plant stimuli. Environ. Microbiol. 2017, 19, 1689–1716. [Google Scholar] [CrossRef] [PubMed]
- Rio-Alvarez, I.; Rodríguez-Herva, J.J.; Cuartas-Lanza, R.; Toth, I.; Pritchard, L.; Rodríguez-Palenzuela, P.; López-Solanilla, E. Genome-wide analysis of the response of Dickeya dadantii 3937 to plant antimicrobial peptides. Mol. Plant Microbe Interact. 2012, 25, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Lv, M.; Chen, Y.; Liao, L.; Liang, Z.; Shi, Z.; Tang, Y.; Ye, S.; Zhou, J.; Zhang, L.-H. Fis is a global regulator critical for modulation of virulence factor production and pathogenicity of Dickeya zeae. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Leonard, S.; Meyer, S.; Lacour, S.; Nasser, W.; Hommais, F.; Reverchon, S. APERO: A genome-wide approach for identifying bacterial small RNAs from RNA-Seq data. Nucleic Acids Res. 2019, 47, e88. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Arana, A.; Lasa, I. Advances in bacterial transcriptome understanding: From overlapping transcription to the excludon concept. Mol. Microbiol. 2020, 113, 593–602. [Google Scholar] [CrossRef]
- Achari, G.A.; Ramesh, R. Chapter 15-Recent advances in quorum quenching of plant pathogenic bacteria. In Advances in Biological Science Research; Meena, S.N., Naik, M.M., Eds.; Academic Press: London, UK; Cambridge, UK; Oxford, UK; San Diego, CA, USA, 2019; pp. 233–245. ISBN 978-0-12-817497-5. [Google Scholar]
- Faure, D.; Dessaux, Y. Quorum sensing as a target for developing control strategies for the plant pathogen Pectobacterium. Eur. J. Plant Pathol. 2007, 119, 353–365. [Google Scholar] [CrossRef]
- Lindow, S.E.; Newman, K.; Chatterjee, S.; Baccari, C.; Iavarone, A.T.; Ionescu, M. Production of Xylella fastidiosa diffusible signal factor in transgenic grape causes pathogen confusion and reduction in severity of Pierce’s disease. Mol. Plant Microbe Interact. 2014, 27, 244–254. [Google Scholar] [CrossRef]
- Vega, C.; Rodríguez, M.; Llamas, I.; Béjar, V.; Sampedro, I. Silencing of phytopathogen communication by the halotolerant PGPR Staphylococcus equorum strain EN21. Microorganisms 2019, 8, 42. [Google Scholar] [CrossRef]
- Rodríguez, M.; Torres, M.; Blanco, L.; Béjar, V.; Sampedro, I.; Llamas, I. Plant growth-promoting activity and quorum quenching-mediated biocontrol of bacterial phytopathogens by Pseudomonas segetis strain P6. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Leitão, A.L.; Costa, M.C.; Gabriel, A.F.G.; Enguita, F.J. Interspecies communication in holobionts by non-coding RNA exchange. Int. J. Mol. Sci. 2020, 21, 2333. [Google Scholar] [CrossRef]
- Samal, B.; Chatterjee, S. New insight into bacterial social communication in natural host: Evidence for interplay of heterogeneous and unison quorum response. PLoS Genet. 2019, 15, e1008395. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Top 10 Rank [7] | Bacterial Pathogen Species | QS Mechanisms | Involvement in Virulence |
|---|---|---|---|
| 1 | Pseudomonas syringae | AHL | Yes |
| 2 | Ralstonia solanacearum | AHL | No |
| DSF-derived signals | Yes | ||
| 3 | Agrobacterium tumefaciens with pTi | AHL | Yes |
| 4 | Xanthomonas oryzae pv oryzae | DSF | Yes |
| 5 | Xanthomonas campestris pv | DSF | Yes |
| 6 | Xanthomonas axonopodis pv | DSF | Yes |
| 7 | Erwinia amylovora | AHL | Yes * |
| 8 | Xylella fastidiosa | DSF | Yes |
| 9 | Dickeya spp. | AHL | No ** |
| Vfm | Yes | ||
| 10 | Pectobacterium carotovorum (and atrosepticum) | AHL | Yes |
| Signal Molecule | Species | Studied Strains | QS System | Pathology | Hosts | Targeted Functions | References |
|---|---|---|---|---|---|---|---|
| OHHL OOHL | Pseudomonas syringae pv. tabaci | 11528 | PsyI/PsyR | Wild-fire disease | Tobacco plants | Swarming, flagellum synthesis, assembly of pili, biofilm formation, chemotaxis, colonization, epiphytic viability, SST2, SST6, alginate synthesis | [15,16,17,18] |
| No production of AHL | Pseudomonas syringae pv. actinidiae | PsaR1, PsaR2, PsaR3 | Bacterial canker | Kiwifruit plants | Regulation of traits associated with survival in planta, cellular multiplication, swarming, oxidative stress resistance | [19,20] | |
| HHL C8-HSL | Ralstonia solanacearum | GMI1000 | SolI/ SolR | Wilting | No Data | [6,21,22,23] | |
| OOHL | Agrobacterium fabrum (tumefaciens) | C58 | TraI/ TraR | Crown Gall | DNA replication, plasmid segregation in daughter cells, conjugative transfer of plasmid Ti | [24,25,26,27,28,29] | |
| OHHL HHL | Erwinia amylovora | Ea2 | EamR/EamI | fire blight | Apple, Pear | Amylovoran, levan, tolerance to hydrogen peroxide | [30,31,32,33,34] |
| OHHL HHL DHL | Dickeya dadantii | 3937 | ExpR/ExpI | Soft rot | Pineapple, Potato, Sweet potato, Banana, Maize, Dianthus spp., Philodendron, Pelargonium, Saintpaulia | No implication in virulence | [35,36,37] |
| OHHL OOHL | Dickeya zeae | EC1 | ExpR/ExpI | Soft rot | Maize, Potato, Pineapple, Banana, Tobacco, Rice, Brachiaria, Chrysanthemum | Swarming, pigment synthesis, cellular aggregate formation, plant colonization, rice seed germination No implication in PCWDE production | [38] |
| OHHL HHL | Dickeya solani | ExpR/ExpI | Soft rot | Potato, Hyacinth | PCWDEs | [39] | |
| OHHL OOHL | Pectobacterium carotovorum | ExpI/ExpR1—ExpR2 CarR/CarI | Soft rot | Potato, carrot, green pepper | PCWDEs, oxidative stress resistance, antimicrobial activity, carbapenem biosynthesis | [4,6,12,40] | |
| OOHL C8-HSL OHHL | Pectobacterium atrosepticum | ExpR/ExpI | Soft rot | Potato and chicory | Pectates lyases | [12,40,41,42,43,44] | |
| OHHL | Pantoea stewartiisubsp. stewartii | Leaf blight and Stewart disease | Corn | Mobility, stewartan production, carotinoids pigments | [45,46] |
| Signal Molecule | Species | QS System | Pathology | Hosts | Targeted Functions | References |
|---|---|---|---|---|---|---|
| DSF BDSF CDSF IDSF DSF-II cis-9-methyl-2-decenoic acid cis-2-undecenoic acid | Xanthomonas campestris pv. campestris (Xcc) | Rfp system | Black rot | Crucifers | PCWDEs, exo-polysaccharide production, multidrug resistance, oxidative stress resistance, mobility, chemotactic response, iron assimilation, Krebs cycle, membrane components and carriers, fatty acid metabolism, cellular aggregates and biofilms, plant innate immunity | [80,81,82,83,84] |
| DSF BDSF CDSF | Xanthomonas oryzae pv. oryzae (Xoo) | Rfp system | Bacterial blight | Rice | PCWDEs, exo-polysaccharide production, multidrug resistance, oxidative stress resistance, mobility, chemotactic response, iron assimilation, Krebs cycle, membrane components and carriers, fatty acid metabolism, cellular aggregates and biofilms, plant innate immunity | [85,86] |
| DSF | Xanthomonas axonopodis pv. glycines (Xag) | Rfp system | Bacterial pustule | Soybean | PCWDEs, exo-polysaccharide production, multidrug resistance, oxidative stress resistance, mobility, chemotactic response, iron assimilation, Krebs cycle, membrane components and carriers, fatty acid metabolism, cellular aggregates and biofilms, plant innate immunity | [87] |
| XfDSF1 XfDSF2 | Xyllela fastidiosa | Rfp system | Pierce’s disease | Mostly dicots: Grapevine, Citrus, Almond tree, Olive tree | Mobility, biofilm formation, type 4 pili and twitching motility, adhesin, Outer Membrane Vesicle (OMV) liberation, attachment, plant colonization and acquisition by insect vectors | [6,7,78,88,89] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baltenneck, J.; Reverchon, S.; Hommais, F. Quorum Sensing Regulation in Phytopathogenic Bacteria. Microorganisms 2021, 9, 239. https://doi.org/10.3390/microorganisms9020239
Baltenneck J, Reverchon S, Hommais F. Quorum Sensing Regulation in Phytopathogenic Bacteria. Microorganisms. 2021; 9(2):239. https://doi.org/10.3390/microorganisms9020239
Chicago/Turabian StyleBaltenneck, Julie, Sylvie Reverchon, and Florence Hommais. 2021. "Quorum Sensing Regulation in Phytopathogenic Bacteria" Microorganisms 9, no. 2: 239. https://doi.org/10.3390/microorganisms9020239
APA StyleBaltenneck, J., Reverchon, S., & Hommais, F. (2021). Quorum Sensing Regulation in Phytopathogenic Bacteria. Microorganisms, 9(2), 239. https://doi.org/10.3390/microorganisms9020239