
YPK9 and WHI2 Negatively Interact during Oxidative Stress


Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains, Media, and Plasmid
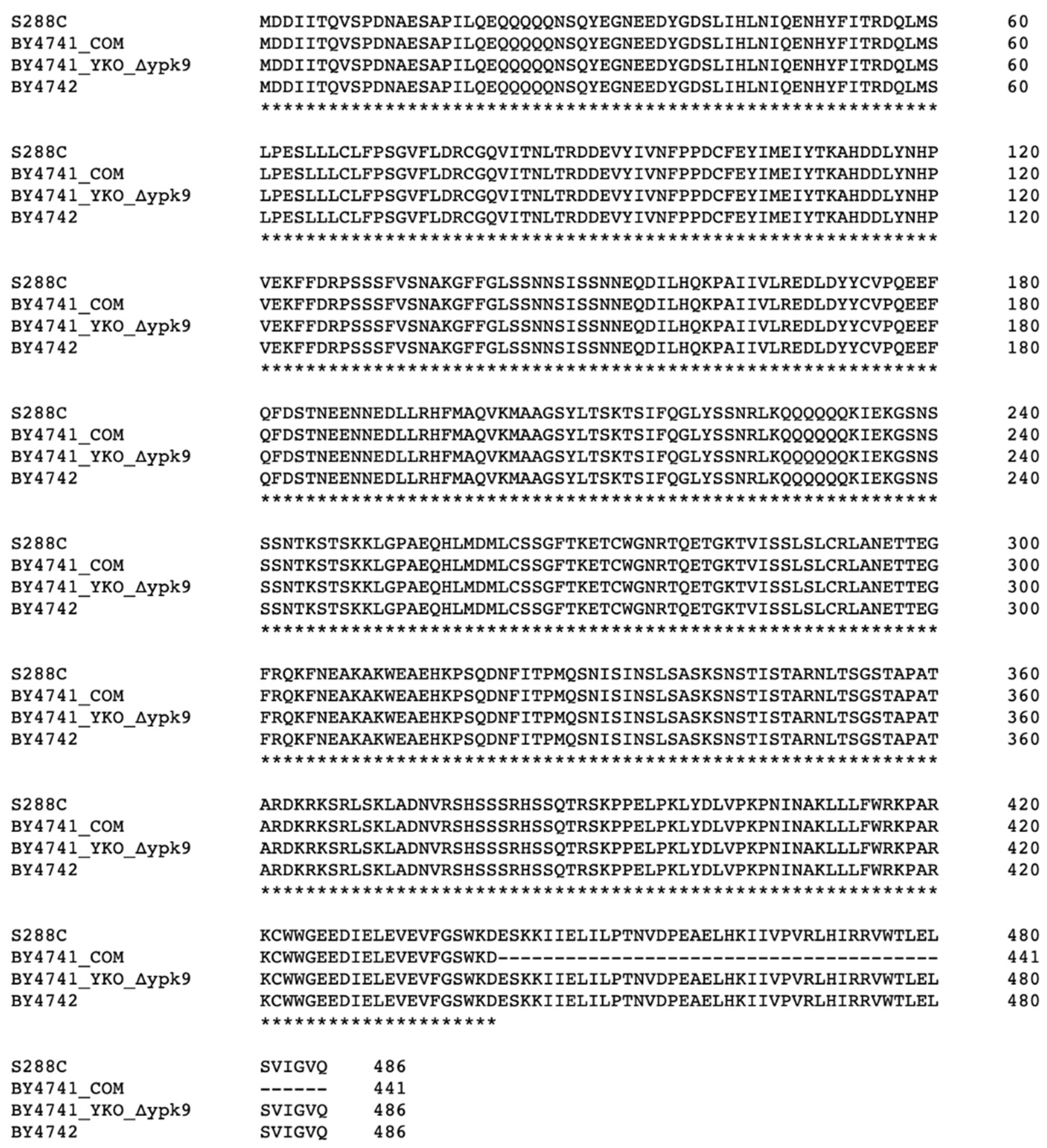
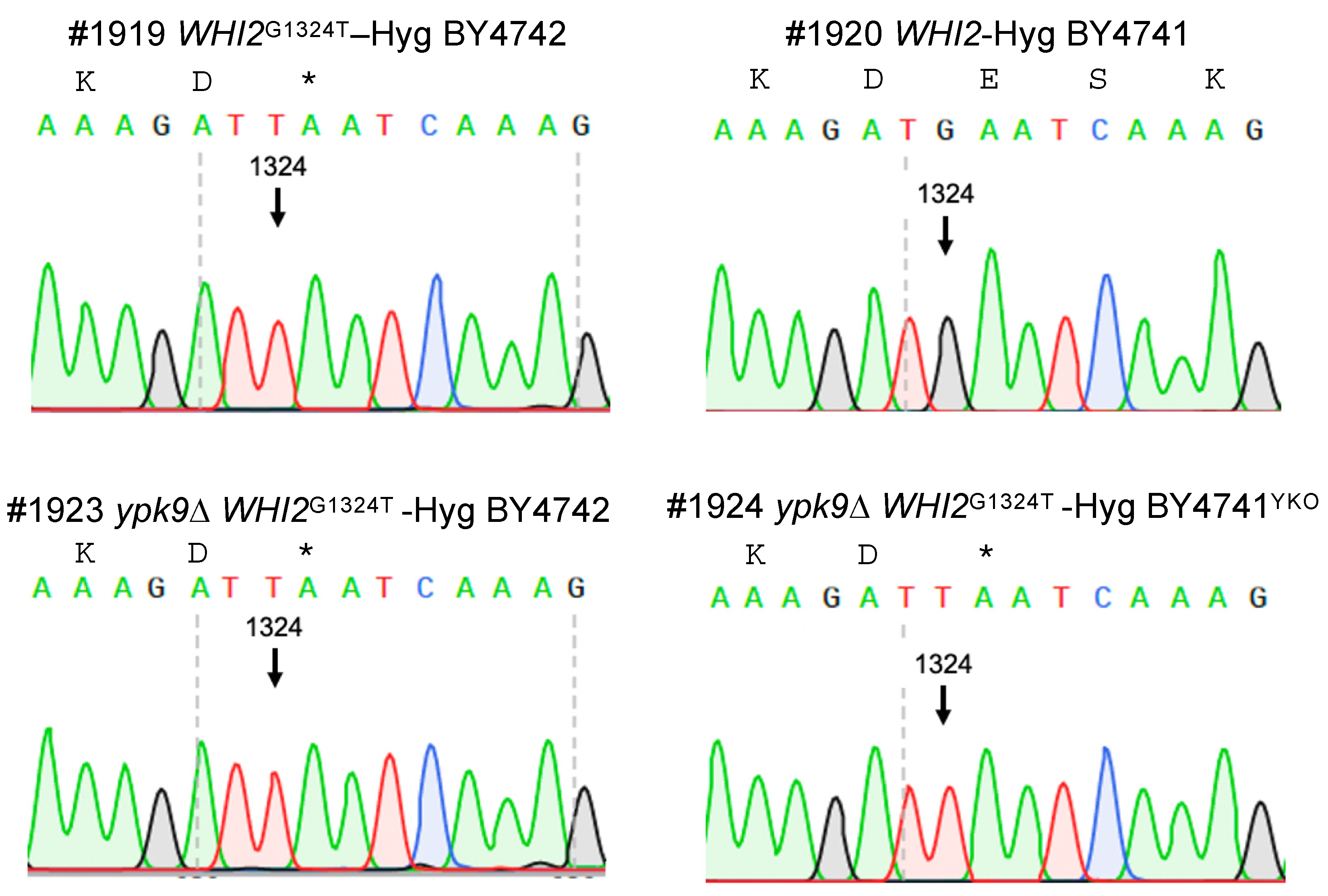
2.2. Strain Genome Sequencing and Analysis
2.3. Construction of WHI2 Variants
2.4. Yeast Growth Assays
2.5. Calculation of the Strength of Genetic Interactions
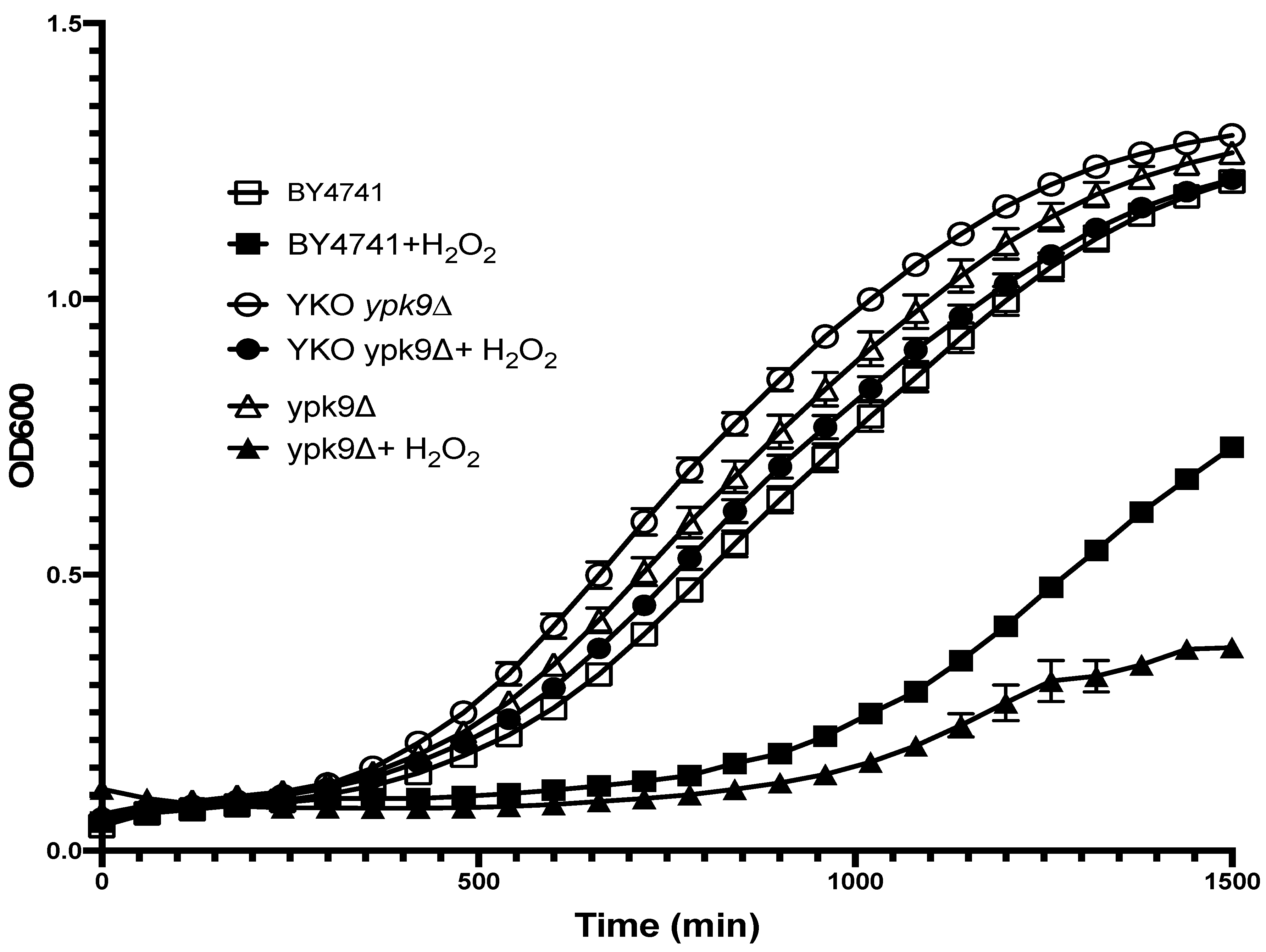
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gitler, A.D.; Chesi, A.; Geddie, M.L.; Strathearn, K.E.; Hamamichi, S.; Hill, K.J.; Caldwell, K.A.; Caldwell, G.A.; Cooper, A.A.; Rochet, J.-C.; et al. Alpha-Synuclein is part of a diverse and highly conserved interaction network that includes PARK9 and manganese toxicity. Nat. Genet. 2009, 41, 308–315. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, K.; Wolfe, D.M.; Stiller, B.; Pearce, D.A. Cd2+, Mn2+, Ni2+ and Se2+ toxicity to Saccharomyces cerevisiae lacking YPK9p the orthologue of human ATP13A2. Biochem. Biophys. Res. Commun. 2009, 383, 198–202. [Google Scholar] [CrossRef] [Green Version]
- van Veen, S.; Martin, S.; Van den Haute, C.; Benoy, V.; Lyons, J.; Vanhoutte, R.; Kahler, J.P.; Decuypere, J.-P.; Gelders, G.; Lambie, E.; et al. ATP13A2 deficiency disrupts lysosomal polyamine export. Nature 2020, 578, 419–424. [Google Scholar] [CrossRef]
- Ramirez, A.; Heimbach, A.; Gründemann, J.; Stiller, B.; Hampshire, D.; Cid, L.P.; Goebel, I.; Mubaidin, A.F.; Wriekat, A.-L.; Roeper, J.; et al. Hereditary parkinsonism with dementia is caused by mutations in ATP13A2, encoding a lysosomal type 5 P-type ATPase. Nat. Genet. 2006, 38, 1184–1191. [Google Scholar] [CrossRef] [PubMed]
- Tabor, C.W.; Tabor, H. Polyamines in microorganisms. Microbiol. Rev. 1985, 49, 81–99. [Google Scholar] [CrossRef]
- Pegg, A.E. Mammalian polyamine metabolism and function. IUBMB Life 2009, 61, 880–894. [Google Scholar] [CrossRef] [PubMed]
- Sudbery, P.E.; Goodey, A.R.; Carter, B.L.A. Genes which control cell proliferation in the yeast Saccharomyces cerevisiae. Nature 1980, 288, 401–404. [Google Scholar] [CrossRef]
- Teng, X.; Yau, E.; Sing, C.; Hardwick, J.M. Whi2 signals low leucine availability to halt yeast growth and cell death. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef]
- Chen, X.; Wang, G.; Zhang, Y.; Dayhoff-Brannigan, M.; Diny, N.L.; Zhao, M.; He, G.; Sing, C.N.; Metz, K.A.; Stolp, Z.D.; et al. Whi2 is a conserved negative regulator of TORC1 in response to low amino acids. PLoS Genet. 2018, 14, e1007592. [Google Scholar] [CrossRef]
- Costanzo, M.; Baryshnikova, A.; Bellay, J.; Kim, Y.; Spear, E.D.; Sevier, C.S.; Ding, H.; Koh, J.L.; Toufighi, K.; Mostafavi, S.; et al. The Genetic Landscape of a Cell. Science 2010, 327, 425–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, S.R.; Dietrich, F.S.; Fisk, D.G.; Binkley, G.; Balakrishnan, R.; Costanzo, M.C.; Dwight, S.S.; Hitz, B.C.; Karra, K.; Nash, R.S.; et al. The Reference Genome Sequence of Saccharomyces cerevisiae: Then and Now. G3: Genes Genomes Genet. 2014, 4, 389–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gietz, R.D.; Schiestl, R.H. High-efficiency yeast transformation using the LiAc/SS carrier DNA/PEG method. Nat. Protoc. 2007, 2, 31–34. [Google Scholar] [CrossRef]
- Goldstein, A.L.; McCusker, J.H. Three new dominant drug resistance cassettes for gene disruption in Saccharomyces cerevisiae. Yeast 1999, 15, 1541–1553. [Google Scholar] [CrossRef]
- Hoffman, C.S.; Winston, F. A ten-minute DNA preparation from yeast efficiently releases autonomous plasmids for transformaion of Escherichia coli. Gene 1987, 57, 267–272. [Google Scholar] [CrossRef]
- Giaever, G.; Chu, A.M.; Ni, L.; Connelly, C.; Riles, L.; Véronneau, S.; Dow, S.; Lucau-Danila, A.; Anderson, K.; André, B.; et al. Functional profiling of the Saccharomyces cerevisiae genome. Nature 2002, 418, 387–391. [Google Scholar] [CrossRef]
- Koh, J.L.Y.; Ding, H.; Costanzo, M.; Baryshnikova, A.; Toufighi, K.; Bader, G.; Myers, C.L.; Andrews, B.J.; Boone, C. DRYGIN: A database of quantitative genetic interaction networks in yeast. Nucleic Acids Res. 2009, 38 (Suppl. 1), D502–D507. [Google Scholar] [CrossRef] [Green Version]
- Teng, X.; Hardwick, J.M. Quantification of Genetically Controlled Cell Death in Budding Yeast. Springer Protoc. Handb. 2013, 1004, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Larroy, C.; Pares, X.; Biosca, J.A. Characterization of a Saccharomyces cerevisiae NADP(H)-dependent alcohol dehydrogenase (ADHVII), a member of the cinnamyl alcohol dehydrogenase family. Eur. J. Biochem. 2002, 269, 5738–5745. [Google Scholar] [CrossRef]
- van Leeuwen, J.; Pons, C.; Mellor, J.C.; Yamaguchi, T.N.; Friesen, H.; Koschwanez, J.; Ušaj, M.M.; Pechlaner, M.; Takar, M.; Ušaj, M.; et al. Exploring genetic suppression interactions on a global scale. Science 2016, 354. [Google Scholar] [CrossRef] [Green Version]
- Hua, Z.; Fatheddin, P.; Graham, T.R. An Essential Subfamily of Drs2p-related P-Type ATPases Is Required for Protein Trafficking between Golgi Complex and Endosomal/Vacuolar System. Mol. Biol. Cell 2002, 13, 3162–3177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pomorski, T.G.; Lombardi, R.; Riezman, H.; Devaux, P.F.; van Meer, G.; Holthuis, J.C.M. Drs2p-related P-type ATPases Dnf1p and Dnf2p Are Required for Phospholipid Translocation across the Yeast Plasma Membrane and Serve a Role in Endocytosis. Mol. Biol. Cell 2003, 14, 1240–1254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherry, J.M.; Hong, E.L.; Amundsen, C.; Balakrishnan, R.; Binkley, G.; Chan, E.; Christie, K.; Costanzo, M.; Dwight, S.S.; Engel, S.; et al. Saccharomyces Genome Database: The genomics resource of budding yeast. Nucleic Acids Res. 2011, 40, D700–D705. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Hiroko, T.; Chaudhuri, A.; Inose, F.; Lord, M.; Tanaka, S.; Chant, J.; Fujita, A. Multigenerational Cortical Inheritance of the Rax2 Protein in Orienting Polarity and Division in Yeast. Science 2000, 290, 1975–1978. [Google Scholar] [CrossRef] [PubMed]
- Fujita, A.; Lord, M.; Hiroko, T.; Hiroko, F.; Chen, T.; Oka, C.; Misumi, Y.; Chant, J. Rax1, a protein required for the establishment of the bipolar budding pattern in yeast. Gene 2004, 327, 161–169. [Google Scholar] [CrossRef]
- Teng, X.; Hardwick, J.M. Whi2: A new player in amino acid sensing. Curr. Genet. 2019, 65, 701–709. [Google Scholar] [CrossRef]
- Kaida, D.; Yashiroda, H.; Toh-e, A.; Kikuchi, Y. Yeast Whi2 and Psr1-phosphatase form a complex and regulate STRE-mediated gene expression. Genes Cells 2002, 7, 543–552. [Google Scholar] [CrossRef]
- Sadeh, A.; Movshovich, N.; Volokh, M.; Gheber, L.; Aharoni, A. Fine-tuning of the Msn2/4–mediated yeast stress responses as revealed by systematic deletion of Msn2/4 partners. Mol. Biol. Cell 2011, 22, 3127–3138. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; López, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Martinez-Pastor, M.T.; Marchler, G.; Schuller, C.; Marchler-Bauer, A.; Ruis, H.; Estruch, F. The Saccharomyces cerevisiae zinc finger proteins Msn2p and Msn4p are required for transcriptional induction through the stress response element (STRE). EMBO J. 1996, 15, 2227–2235. [Google Scholar] [CrossRef] [PubMed]
- Gallegos, J.E.; Hayrynen, S.; Adames, N.R.; Peccoud, J. Challenges and opportunities for strain verification by whole-genome sequencing. Sci. Rep. 2020, 10, 5873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comyn, S.A.; Flibotte, S.; Mayor, T. Recurrent background mutations in WHI2 impair proteostasis and degradation of misfolded cytosolic proteins in Saccharomyces cerevisiae. Sci. Rep. 2017, 7, 4183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, W.-C.; Teng, X.; Park, H.K.; Tucker, C.M.; Dunham, M.; Hardwick, J.M. Fis1 deficiency selects for compensatory mutations responsible for cell death and growth control defects. Cell Death Differ. 2008, 15, 1838–1846. [Google Scholar] [CrossRef]
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, Y.; Gruhler, A.; Heilbut, A.; Bader, G.D.; Moore, L.; Adams, S.-L.; Millar, A.; Taylor, P.; Bennett, K.L.; Boutilier, K.; et al. Systematic identification of protein complexes in Saccharomyces cerevisiae by mass spectrometry. Nature 2002, 415, 180–183. [Google Scholar] [CrossRef] [Green Version]
- Krüger, A.; Vowinckel, J.; Mülleder, M.; Grote, P.; Capuano, F.; Bluemlein, K.; Ralser, M. Tpo1-mediated spermine and spermidine export controls cell cycle delay and times antioxidant protein expression during the oxidative stress response. EMBO Rep. 2013, 14, 1113–1119. [Google Scholar] [CrossRef]
- Sørensen, D.M.; Holemans, T.; Van Veen, S.; Martin, S.; Arslan, T.; Haagendahl, I.W.; Holen, H.W.; Hamouda, N.N.; Eggermont, J.; Palmgren, M.; et al. Parkinson disease related ATP13A2 evolved early in animal evolution. PLoS ONE 2018, 13, e0193228. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, T.; Knauer, H.; Schauer, A.; Büttner, S.; Ruckenstuhl, C.; Carmona-Gutierrez, D.; Ring, J.; Schroeder, S.; Magnes, C.; Antonacci, L.; et al. Induction of autophagy by spermidine promotes longevity. Nat. Cell Biol. 2009, 11, 1305–1314. [Google Scholar] [CrossRef]
- Wirth, M.; Schwarz, C.; Benson, G.; Horn, N.; Buchert, R.; Lange, C.; Köbe, T.; Hetzer, S.; Maglione, M.; Michael, E.; et al. Effects of spermidine supplementation on cognition and biomarkers in older adults with subjective cognitive decline (SmartAge)—study protocol for a randomized controlled trial. Alzheimer’s Res. Ther. 2019, 11, 1–17. [Google Scholar] [CrossRef]
- Lewandowski, N.M.; Ju, S.; Verbitsky, M.; Ross, B.; Geddie, M.L.; Rockenstein, E.; Adame, A.; Muhammad, A.; Vonsattel, J.P.; Ringe, D.; et al. Polyamine pathway contributes to the pathogenesis of Parkinson disease. Proc. Natl. Acad. Sci. USA 2010, 107, 16970–16975. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | ||||||||
|---|---|---|---|---|---|---|---|---|
| Standard Name | Name Description | Chromosome Location | SNP | Amino Acid Change | BY4741COM | BY4741COM ypk9∆ | BY4741YKO ypk9∆ | BY4742 |
| ADH7 | Alcohol Dehydrogenase | III/ 309,566 | G to A | Gly166Asp | + | + | − | − |
| DNF2 | Drs2 Neo1 Family | IV/ 632,116 | G to C | Gly279Arg | + | + | − | − |
| WHI2 | Whiskey 2 | XV/ 412,193 | G to T | Glu442 * | + | + | − | − |
| RAX1 | Revert to AXial 1 | XV/ 881,528 | C to A | Cys188 * | + | + | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joseph, F.; Miller, D.; Evgrafov, O.V.; Chirico, W.J. YPK9 and WHI2 Negatively Interact during Oxidative Stress. Microorganisms 2021, 9, 2584. https://doi.org/10.3390/microorganisms9122584
Joseph F, Miller D, Evgrafov OV, Chirico WJ. YPK9 and WHI2 Negatively Interact during Oxidative Stress. Microorganisms. 2021; 9(12):2584. https://doi.org/10.3390/microorganisms9122584
Chicago/Turabian StyleJoseph, Florenal, Darach Miller, Oleg V. Evgrafov, and William J. Chirico. 2021. "YPK9 and WHI2 Negatively Interact during Oxidative Stress" Microorganisms 9, no. 12: 2584. https://doi.org/10.3390/microorganisms9122584
APA StyleJoseph, F., Miller, D., Evgrafov, O. V., & Chirico, W. J. (2021). YPK9 and WHI2 Negatively Interact during Oxidative Stress. Microorganisms, 9(12), 2584. https://doi.org/10.3390/microorganisms9122584