
Epstein-Barr Virus Infection Alone or Jointly with Human Papillomavirus Associates with Down-Regulation of miR-145 in Oral Squamous-Cell Carcinoma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Specimen
2.2. Cell Lines
2.3. Detection of HPV and EBV Infection in OSCC Tissues
2.4. Detection of miR-145 and DNMTs Expression by qRT-PCR
2.5. Methylation Status of miR-145 Promoter Determined by Methylation-Specific Real-Time PCR (MSP)
2.6. Statistical Analysis
3. Results
3.1. Patient Characteristics
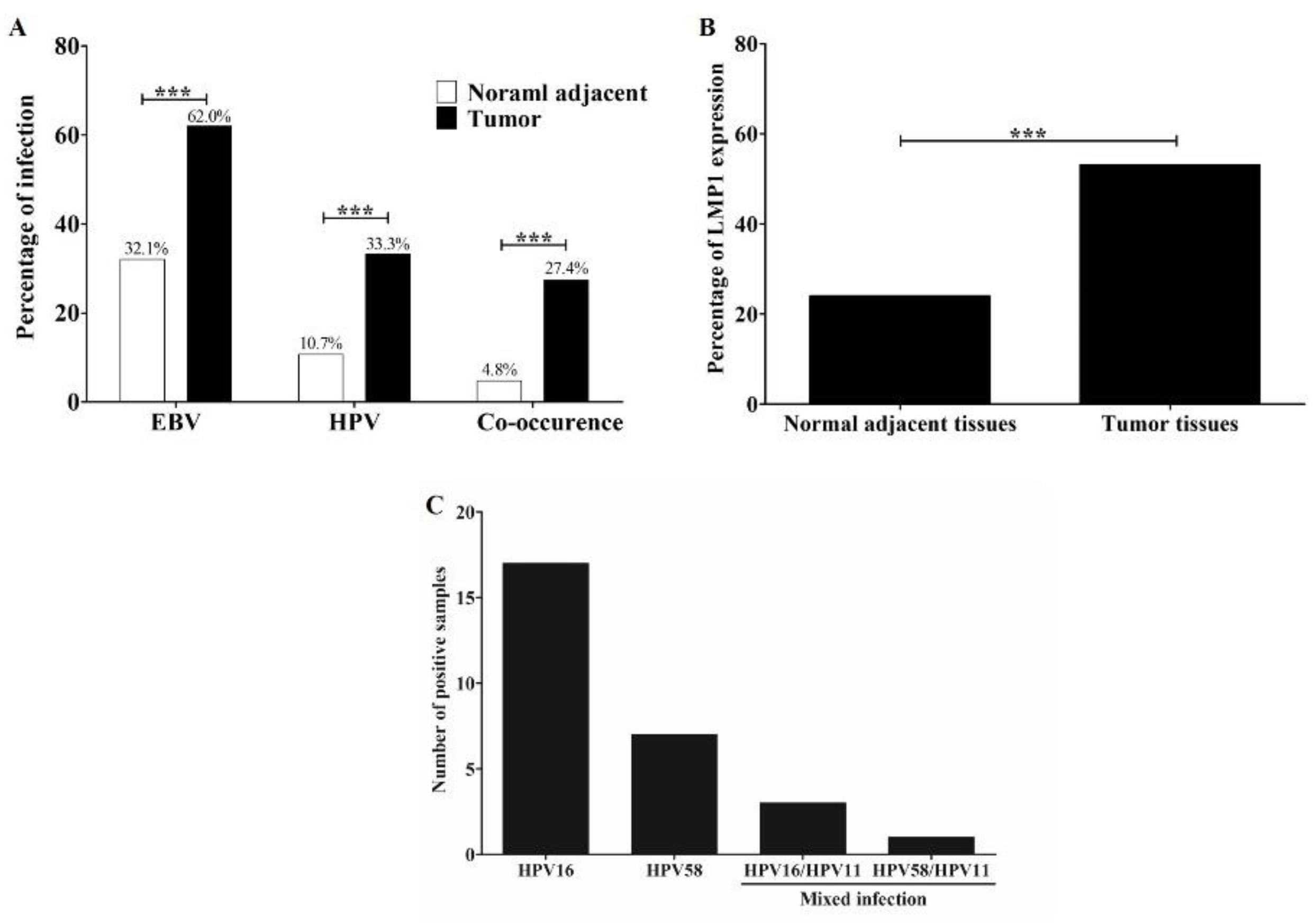
3.2. Prevalence of HPV, EBV, and Their Co-Occurrence in OSCC
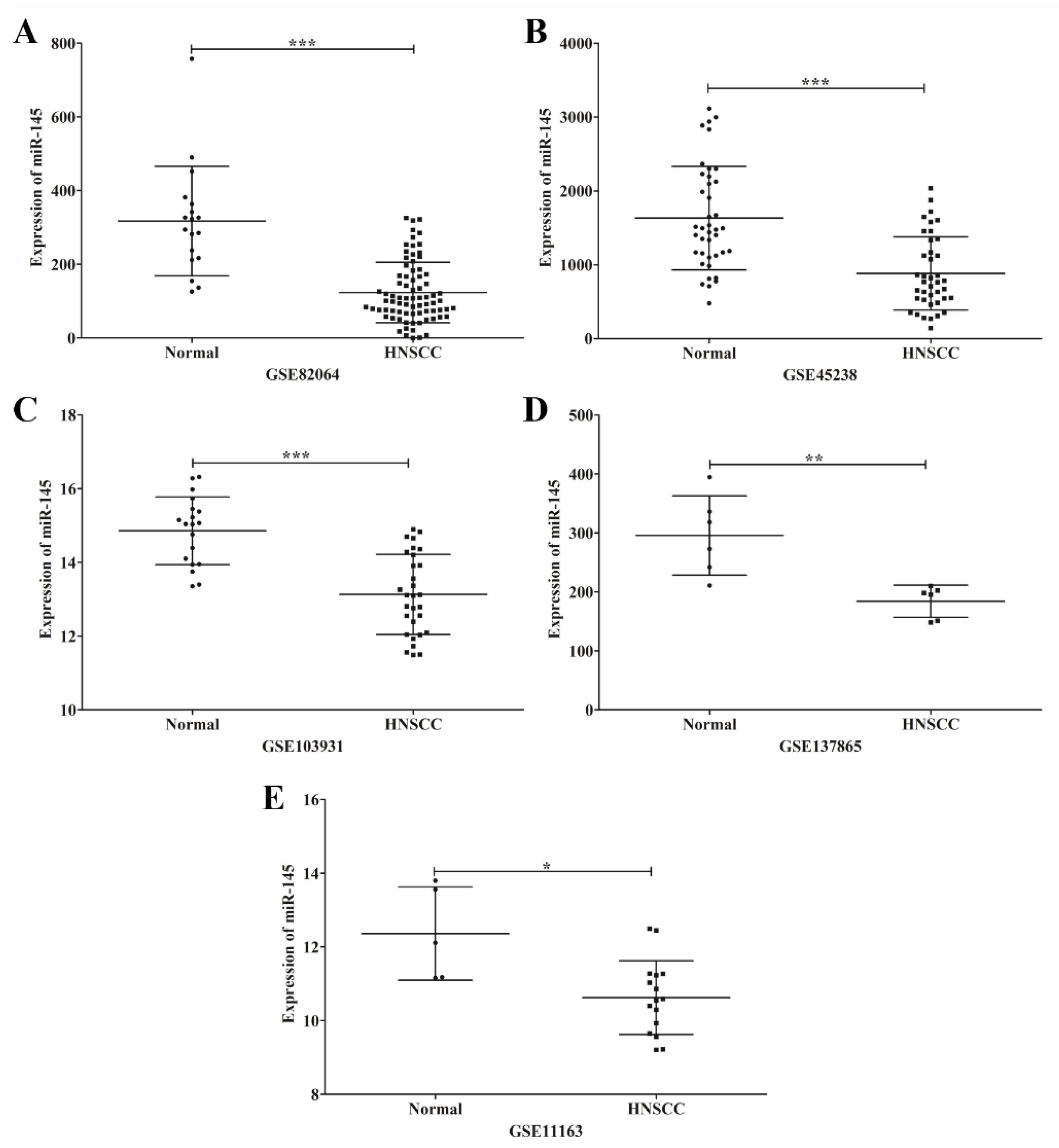
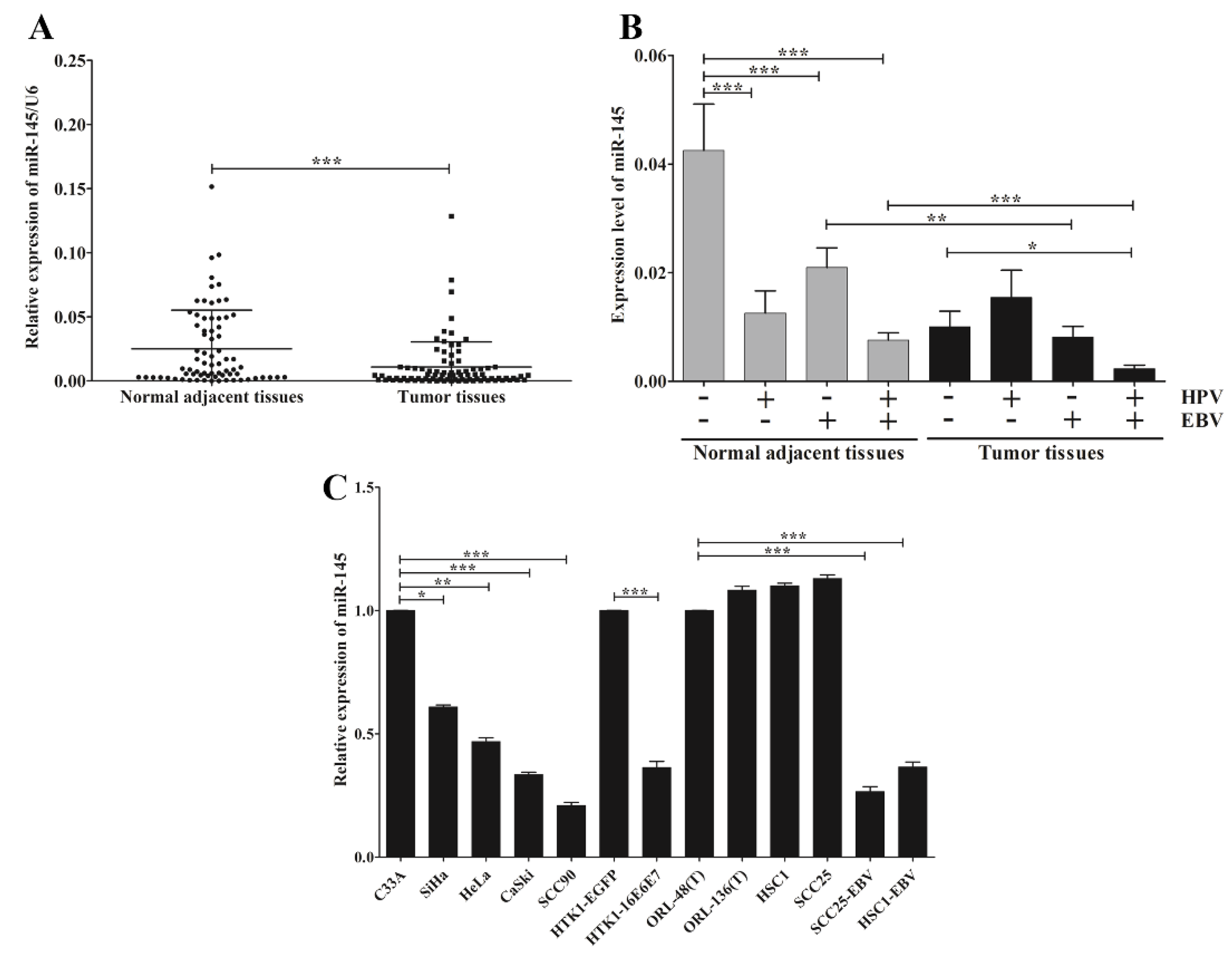
3.3. EBV and HPV Modulate miR-145 Expression in OSCC
3.4. EBV and HPV, Separately or Jointly, Associate with the Silencing of miR-145 via DNA Hypermethylation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, M.; Nanavati, R.; Modi, T.G.; Dobariya, C. Oral cancer: Etiology and risk factors: A review. J. Cancer Res. Ther. 2016, 12, 458. [Google Scholar] [CrossRef]
- Gupta, K.; Metgud, R. Evidences suggesting involvement of viruses in oral squamous cell carcinoma. Patholog. Res. Int. 2013, 2013, 642496. [Google Scholar] [CrossRef] [Green Version]
- Jalouli, J.; Jalouli, M.M.; Sapkota, D.; Ibrahim, S.O.; Larsson, P.A.; Sand, L. Human papilloma virus, herpes simplex virus and epstein barr virus in oral squamous cell carcinoma from eight different countries. Anticancer Res. 2012, 32, 571–580. [Google Scholar]
- Gillison, M.L.; Koch, W.M.; Capone, R.B.; Spafford, M.; Westra, W.H.; Wu, L.; Wu, L.; Zahurak, M.L.; Daniel, R.W.; Viglione, M.; et al. Evidence for a causal association between human papillomavirus and a subset of head and neck cancers. J. Natl. Cancer Inst. 2000, 92, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Ekshyyan, O.; Moore-Medlin, T.; Rong, X.; Nathan, S.; Gu, X.; Abreo, F.; Rosenthal, E.L.; Shi, M.; Guidry, J.T.; et al. Association between human papilloma virus/Epstein–Barr virus coinfection and oral carcinogenesis. J. Oral Pathol. Med. 2015, 44, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Whiteside, M.A.; Siegel, E.M.; Unger, E.R. Human papillomavirus and molecular considerations for cancer risk. Cancer 2008, 113, 2981–2994. [Google Scholar] [CrossRef]
- Scott, R.S. Epstein–Barr virus: A master epigenetic manipulator. Curr. Opin. Virol. 2017, 26, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Wences, H.; Peralta-Zaragoza, O.; Fernandez-Tilapa, G. Human papilloma virus, DNA methylation and microRNA expression in cervical cancer. Oncol. Rep. 2014, 31, 2467–2476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.J.; Shin, Y.S.; Kang, B.W.; Kim, J.G.; Won, K.J.; Lieberman, P.M.; Cho, H.; Kang, H. DNA hypermethylation induced by Epstein-Barr virus in the development of Epstein-Barr virus-associated gastric carcinoma. Arch. Pharm. Res. 2017, 40, 894–905. [Google Scholar] [CrossRef]
- Jansson, M.D.; Lund, A.H. MicroRNA and cancer. Mol. Oncol. 2012, 6, 590–610. [Google Scholar] [CrossRef]
- Lajer, C.B.; Garnaes, E.; Friis-Hansen, L.; Norrild, B.; Therkildsen, M.H.; Glud, M.; Rossing, M.; Lajer, H.; Svane, D.; Skotte, L.; et al. The role of miRNAs in human papilloma virus (HPV)-associated cancers: Bridging between HPV-related head and neck cancer and cervical cancer. Br. J. Cancer. 2012, 106, 1526–1534. [Google Scholar] [CrossRef]
- Cui, S.Y.; Wang, R.; Chen, L.B. MicroRNA-145: A potent tumour suppressor that regulates multiple cellular pathways. J. Cell Mol. Med. 2014, 18, 1913–1926. [Google Scholar] [CrossRef] [PubMed]
- Zeinali, T.; Mansoori, B.; Mohammadi, A.; Baradaran, B. Regulatory mechanisms of miR-145 expression and the importance of its function in cancer metastasis. Biomed. Pharmacother. 2019, 109, 195–207. [Google Scholar] [CrossRef]
- Xu, L.; Zhang, Y.; Tang, J.; Wang, P.; Li, L.; Yan, X.; Zheng, X.; Ren, S.; Zhang, M.; Xu, M. The prognostic value and regulatory mechanisms of microRNA-145 in various tumors: A systematic review and meta-analysis of 50 studies. Cancer Epidemiol. Biomark. Prev. 2019, 28, 867–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Z.; Han, Y.; Song, C.; Wei, H.; Chen, Y.; Huang, K.; Li, S.; Ma, D.; Wang, S.; Wang, J.; et al. Systematic review and meta-analysis of the prognostic significance of microRNAs in cervical cancer. Oncotarget 2018, 9, 17141–17148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, S.; Ekalaksananan, T.; Vatanasapt, P.; Loyha, K.; Phusingha, P.; Promthet, S.; Kongyingyoes, B.; Pientong, C. Association of Epstein-Barr virus infection with oral squamous cell carcinoma in a case-control study. J. Oral Pathol. Med. 2015, 44, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Phusingha, P.; Ekalaksananan, T.; Patravoot Vatanasapt, P.; Loyha, K.; Promthet, S.; Kongyingyoes, B.; Patarapadungkit, N.; Chuerduangphui, J.; Pientong, C. Human papillomavirus (HPV) infection in a case-control study of oral squamous cell carcinoma and its increasing trend in northeastern Thailand. J. Med. Virol. 2017, 89, 1096–1101. [Google Scholar] [CrossRef]
- Heawchaiyaphum, C.; Iizasa, H.; Ekalaksananan, T.; Burassakarn, A.; Kiyono, T.; Kanehiro, Y.; Yoshiyama, H.; Pientong, C. Epstein–Barr Virus Infection of Oral Squamous Cells. Microorganisms 2020, 8, 419. [Google Scholar] [CrossRef] [Green Version]
- van den Brule, A.J.; Pol, R.; Fransen-Daalmeijer, N.; Schouls, L.M.; Meijer, C.J.; Snijders, P.J. GP5+/6+ PCR followed by reverse line blot analysis enables rapid and high-throughput identification of human papillomavirus genotypes. J. Clin. Microbiol. 2002, 40, 779–787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozo, F.; Tenorio, A. Detection and typing of lymphotropic herpesviruses by multiplex polymerase chain reaction. J. Virol. Methods 1999, 79, 9–19. [Google Scholar] [CrossRef]
- Aromseree, S.; Chaiwongkot, A.; Ekalaksananan, T.; Kongyingyoes, B.; Patarapadungkit, N.; Pientong, C. The three most common human papillomavirus oncogenic types and their integration state in Thai women with cervical precancerous lesions and carcinomas. J. Med. Virol. 2014, 86, 1911–1919. [Google Scholar] [CrossRef]
- Guo, Y.H.; Zhang, C.; Shi, J.; Xu, M.H.; Liu, F.; Yuan, H.H.; Wang, J.Y.; Jiang, B.; Gao, F.H. Abnormal activation of the EGFR signaling pathway mediates the downregulation of miR-145 through the ERK1/2 in non-small cell lung cancer. Oncol. Rep. 2014, 31, 1940–1946. [Google Scholar] [CrossRef] [Green Version]
- Girault, I.; Tozlu, S.; Lidereau, R.; Bièche, I. Expression analysis of DNA methyltransferases 1, 3A, and 3B in sporadic breast carcinomas. Clin. Cancer Res. 2003, 9, 4415–4422. [Google Scholar] [PubMed]
- Suh, S.O.; Chen, Y.; Zaman, M.S.; Hirata, H.; Yamamura, S.; Shahryari, V.; Liu, J.; Tabatabai, Z.L.; Kakar, S.; Deng, G.; et al. MicroRNA-145 is regulated by DNA methylation and p53 gene mutation in prostate cancer. Carcinogenesis 2011, 32, 772–778. [Google Scholar] [CrossRef] [Green Version]
- Xue, G.; Ren, Z.; Chen, Y.; Zhu, J.; Du, Y.; Pan, D.; Li, X.; Hu, B. A feedback regulation between miR-145 and DNA methyltransferase 3b in prostate cancer cell and their responses to irradiation. Cancer Lett. 2015, 361, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Hong Peng, H.; Chen, Y.; Gong, P.; Cai, L.; Lyu, X.; Jiang, Q.; Wang, J.; Lu, J.; Yao, K.; Liu, K.; et al. Higher methylation intensity induced by EBV LMP1 via NF-κB/DNMT3b signaling contributes to silencing of PTEN gene. Oncotarget 2016, 7, 40025–40037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuss-Duerkop, S.K.; Westrich, J.A.; Pyeon, D. DNA tumor virus regulation of host DNA methylation and its implications for immune evasion and oncogenesis. Viruses 2018, 10, 82. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Qin, J.; Chen, A.; Zhou, J.; Liu, J.; Cheng, J.; Qiu, J.; Zhang, J. Downregulation of microRNA-145 is associated with aggressive progression and poor prognosis in human cervical cancer. Tumor Biol. 2015, 36, 3703–3708. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Sun, D.; Xie, P. Elevated microRNA-145 inhibits the development of oral squamous cell carcinoma through inactivating ERK/MAPK signaling pathway by down-regulating HOXA1. Biosci. Rep. 2019, 39, BSR20182214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, C.; Du, F.; Sun, L.; Li, T.; Li, T.; Min, Y.; Nie, A.; Wang, X.; Geng, L.; Lu, Y.; et al. miR-143 and miR-145 inhibit gastric cancer cell migration and metastasis by suppressing MYO6. Cell Death Dis. 2017, 8, e3101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Jiang, H.; Chen, Z.; Lu, B.; Zhu, Y.; Mao, J.; Chai, K.; Chen, W. MicroRNA-145 Inhibits Cell Migration and Invasion in Colorectal Cancer by Targeting TWIST. Onco Targets Ther. 2019, 12, 10799–10809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, Y.; Qu, Y.; Dang, S.; Yao, B.; Ji, M. MiR-145 inhibits oral squamous cell carcinoma (OSCC) cell growth by targeting c-Myc and Cdk6. Cancer Cell Int. 2013, 13, 51. [Google Scholar] [CrossRef] [Green Version]
- Bazot, Q.; Paschos, K.; Skalska, L.; Kalchschmidt, J.S.; Parker, G.A.; Allday, M.J. Epstein-Barr virus proteins EBNA3A and EBNA3C together induce expression of the oncogenic microRNA cluster miR-221/miR-222 and ablate expression of its target p57KIP2. PLoS Pathog. 2015, 11, e1005031. [Google Scholar] [CrossRef] [Green Version]
- Gao, L.; Ren, W.; Chang, S.; Guo, B.; Huang, S.; Li, M.; Guo, Y.; Li, Z.; Song, T.; Zhi, K.; et al. Downregulation of miR-145 expression in oral squamous cell carcinomas and its clinical significance. Onkologie 2013, 36, 194–199. [Google Scholar] [CrossRef]
- Gao, W.; Zhang, C.; Li, W.; Li, H.; Sang, J.; Zhao, Q.; Bo, Y.; Luo, H.; Zheng, X.; Lu, Y.; et al. Promoter methylation-regulated miR-145-5p inhibits laryngeal squamous cell carcinoma progression by targeting FSCN1. Mol. Ther. 2019, 27, 365–379. [Google Scholar] [CrossRef] [Green Version]
- Xia, W.; Chen, Q.; Wang, J.; Mao, Q.; Dong, G.; Shi, R.; Zheng, Y.; Xu, L.; Jiang, F. DNA methylation mediated silencing of microRNA-145 is a potential prognostic marker in patients with lung adenocarcinoma. Sci. Rep. 2015, 5, 16901. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Tang, Y. SChLAP1 promotes prostate cancer development through interacting with EZH2 to mediate promoter methylation modification of multiple miRNAs of chromosome 5 with a DNMT3a-feedback loop. Cell Death Dis. 2021, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ji, G.; Xiao, X.; Chen, X.; Qin, W.W.; Yang, F.; Li, Y.F.; Fan, L.N.; Xi, W.J.; Huo, Y.; et al. Epigenetically regulated miR-145 suppresses colon cancer invasion and metastasis by targeting LASP1. Oncotarget 2016, 7, 68674–68687. [Google Scholar] [CrossRef] [Green Version]
- Katsushima, K.; Natsume, A.; Ohka, F.; Shinjo, K.; Hatanaka, A.; Ichimura, N.; Sato, S.; Takahashi, S.; Kimura, H.; Totoki, Y.; et al. Targeting the Notch-regulated non-coding RNA TUG1 for glioma treatment. Nat. Commun. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Du, M.; Zhang, Q.; Zhang, W.; Fan, Y.; Yin, L.; Fei, Q.; Jiang, X.; Chen, W.; Zhu, H.; et al. Long noncoding RNA UCA1 promotes the proliferation, invasion, and migration of nasopharyngeal carcinoma cells via modulation of miR-145. Onco Targets Ther. 2018, 11, 7483–7492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Yu, D.; Li, Z.; Zhao, C.; Su, C.; Ning, J. Long non-coding RNA AFAP1-AS1 facilitates the growth and invasiveness of oral squamous cell carcinoma by regulating the miR-145/HOXA1 axis. Oncol. Rep. 2021, 45, 1094–1104. [Google Scholar] [CrossRef]
- Su, Y.; Yu, T.; Wang, Y.; Huang, X.; Wei, X. Circular RNA circDNM3OS Functions as a miR-145-5p Sponge to Accelerate Cholangiocarcinoma Growth and Glutamine Metabolism by Upregulating MORC2. Onco Targets Ther. 2021, 14, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Saha, A.; Jha, H.C.; Upadhyay, S.K.; Robertson, E.S. Epigenetic silencing of tumor suppressor genes during in vitro Epstein–Barr virus infection. Proc. Natl. Acad. Sci. USA 2015, 112, E5199–E5207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seo, S.Y.; Kim, E.O.; Jang, K.L. Epstein–Barr virus latent membrane protein 1 suppresses the growth-inhibitory effect of retinoic acid by inhibiting retinoic acid receptor-β2 expression via DNA methylation. Cancer Lett. 2008, 270, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.L.; Li, H.P.; Lu, Y.J.; Hsueh, C.; Liang, Y.; Chen, C.L.; Tsao, S.W.; Tse, K.P.; Yu, J.S.; Chang, Y.S. Activation of DNA methyltransferase 1 by EBV LMP1 involves C-Jun NH2-terminal kinase signaling. Cancer Res. 2006, 66, 11668–11676. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward (5′-3′) | Reward (5′-3′) |
|---|---|---|
| Methylated-miR-145 | GGGTTTTCGGTATTTTTTAGGGTAATTGAAGTTTC | TAAAATACCACACGTCGCCG |
| Unmethylated-miR-145 | GGGTTTTTGGTATTTTTTAGGGTAATTGAAGTTTT | AACCAAAATAAAATACCACACATCACCA |
| GP5+/GP6+ | TTTGTTACTGTGGTAGATACTAC | GAAAAATAAACTGTAAATCATATTC |
| BALF5 | CGAGTCATCTACGGGGACACGGA | AGCACCCCCACATATCTCTTCTT |
| EBER | CTACGCTGCCCTAGAGGTTTT | CAGCTGGTACTTGACCGAAGA |
| LMP1 | TCCTCCTCTTGGCGCTACTG | TCATCACTGTGTCGTTGTCC |
| miR-145 | ATCGTCCAGTTTTCCCAGG | CGCCTCCACACACTCACC |
| DNMT1 | TACCTGGACGACCCTGACCTC | CGTTGGCATCAAAGATGGACA |
| DNMT3B | GGCAAGTTCTCCGAGGTCTCTG | TGGTACATGGCTTTTCGATAGGA |
| GAPDH | TCATCAGCAATGCCTCCTGCA | TGGGTAGCAGTGATGGCA |
| Demographic Features | OSCC Tissues |
|---|---|
| (n = 84) | |
| Age, year | 60.65 |
| Range, year | 27–90 |
| Gender | |
| Male | 41 |
| Female | 31 |
| ND | 12 |
| Site of OSCC | |
| Tongue | 42 |
| Lip | 4 |
| Buccal mucosa | 8 |
| Gum | 5 |
| Floor of mouth | 1 |
| Palate | 10 |
| Gingiva | 2 |
| ND | 12 |
| Histological grades | |
| Well differentiated | 46 |
| Moderately differentiated | 15 |
| Poorly differentiated | 3 |
| ND | 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heawchaiyaphum, C.; Ekalaksananan, T.; Patarapadungkit, N.; Worawichawong, S.; Pientong, C. Epstein-Barr Virus Infection Alone or Jointly with Human Papillomavirus Associates with Down-Regulation of miR-145 in Oral Squamous-Cell Carcinoma. Microorganisms 2021, 9, 2496. https://doi.org/10.3390/microorganisms9122496
Heawchaiyaphum C, Ekalaksananan T, Patarapadungkit N, Worawichawong S, Pientong C. Epstein-Barr Virus Infection Alone or Jointly with Human Papillomavirus Associates with Down-Regulation of miR-145 in Oral Squamous-Cell Carcinoma. Microorganisms. 2021; 9(12):2496. https://doi.org/10.3390/microorganisms9122496
Chicago/Turabian StyleHeawchaiyaphum, Chukkris, Tipaya Ekalaksananan, Natcha Patarapadungkit, Suchin Worawichawong, and Chamsai Pientong. 2021. "Epstein-Barr Virus Infection Alone or Jointly with Human Papillomavirus Associates with Down-Regulation of miR-145 in Oral Squamous-Cell Carcinoma" Microorganisms 9, no. 12: 2496. https://doi.org/10.3390/microorganisms9122496
APA StyleHeawchaiyaphum, C., Ekalaksananan, T., Patarapadungkit, N., Worawichawong, S., & Pientong, C. (2021). Epstein-Barr Virus Infection Alone or Jointly with Human Papillomavirus Associates with Down-Regulation of miR-145 in Oral Squamous-Cell Carcinoma. Microorganisms, 9(12), 2496. https://doi.org/10.3390/microorganisms9122496