
Exploring the Diversity and Biotechnological Potential of Cultured and Uncultured Coral-Associated Bacteria



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
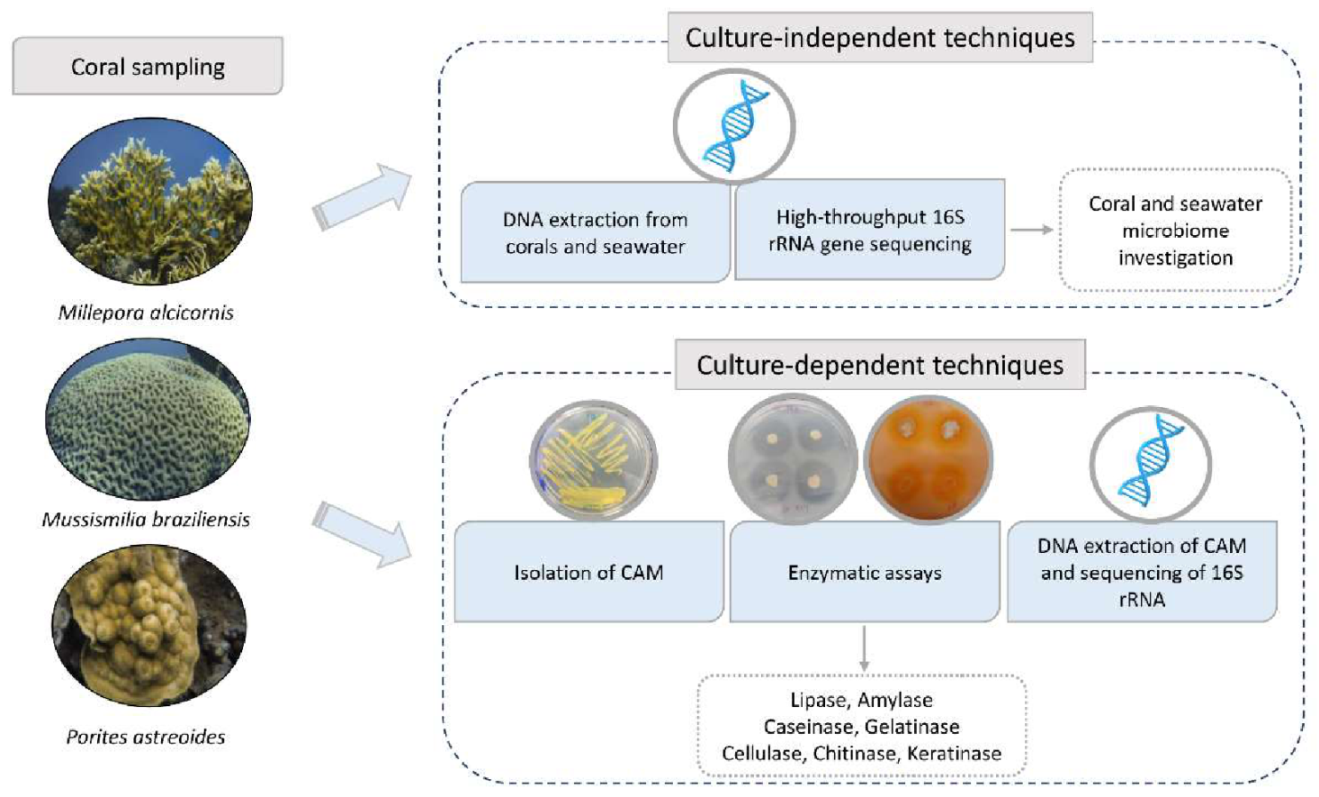
2. Materials and Methods
2.1. Sample Collection
2.2. DNA Extraction from Corals and Seawater
2.3. Bacterial Composition by 16S rRNA Gene Sequencing
2.4. Bacteria Isolation
2.5. Enzymatic Assays
2.6. DNA Extraction from Isolates
2.7. Identification of Isolates
3. Results
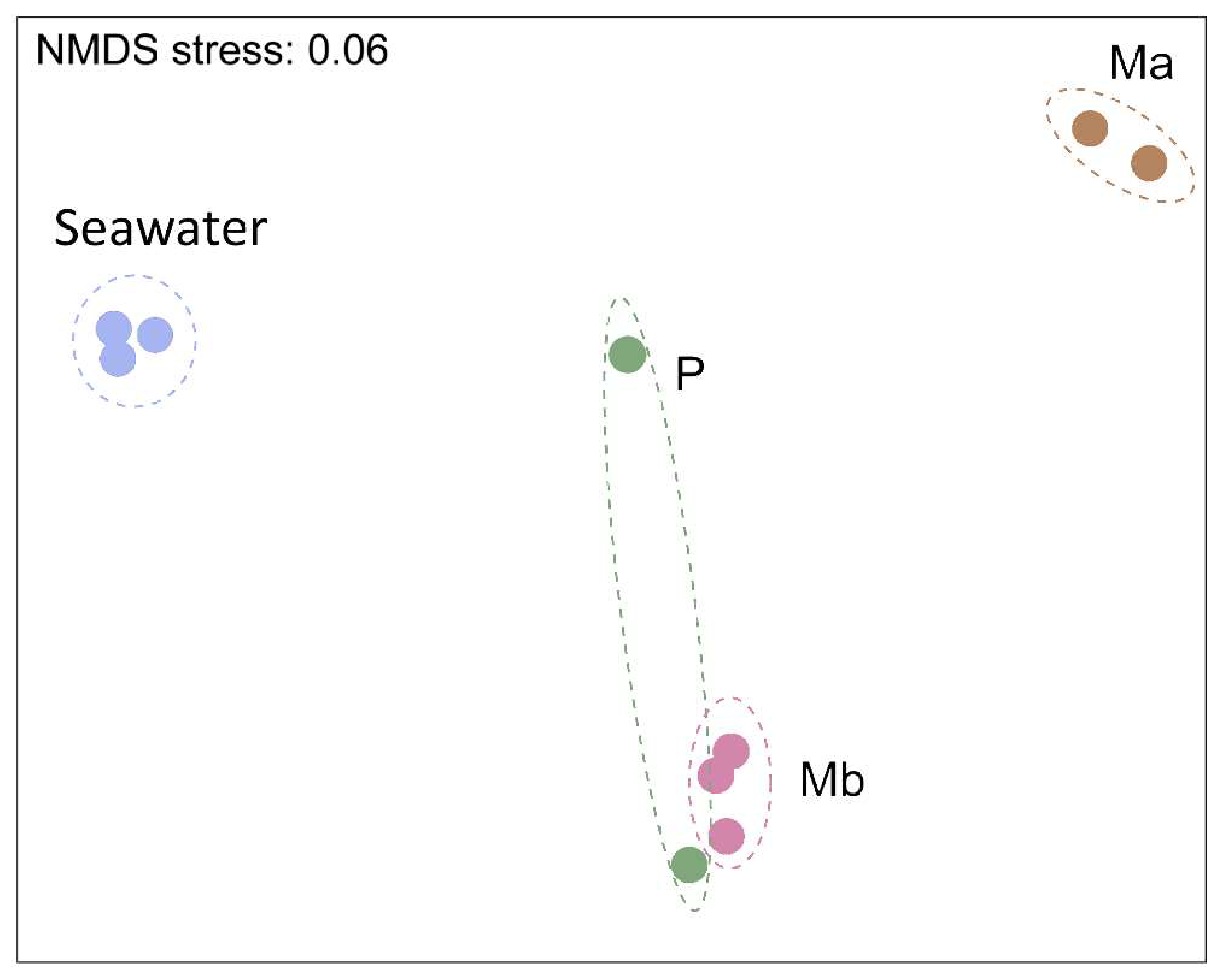
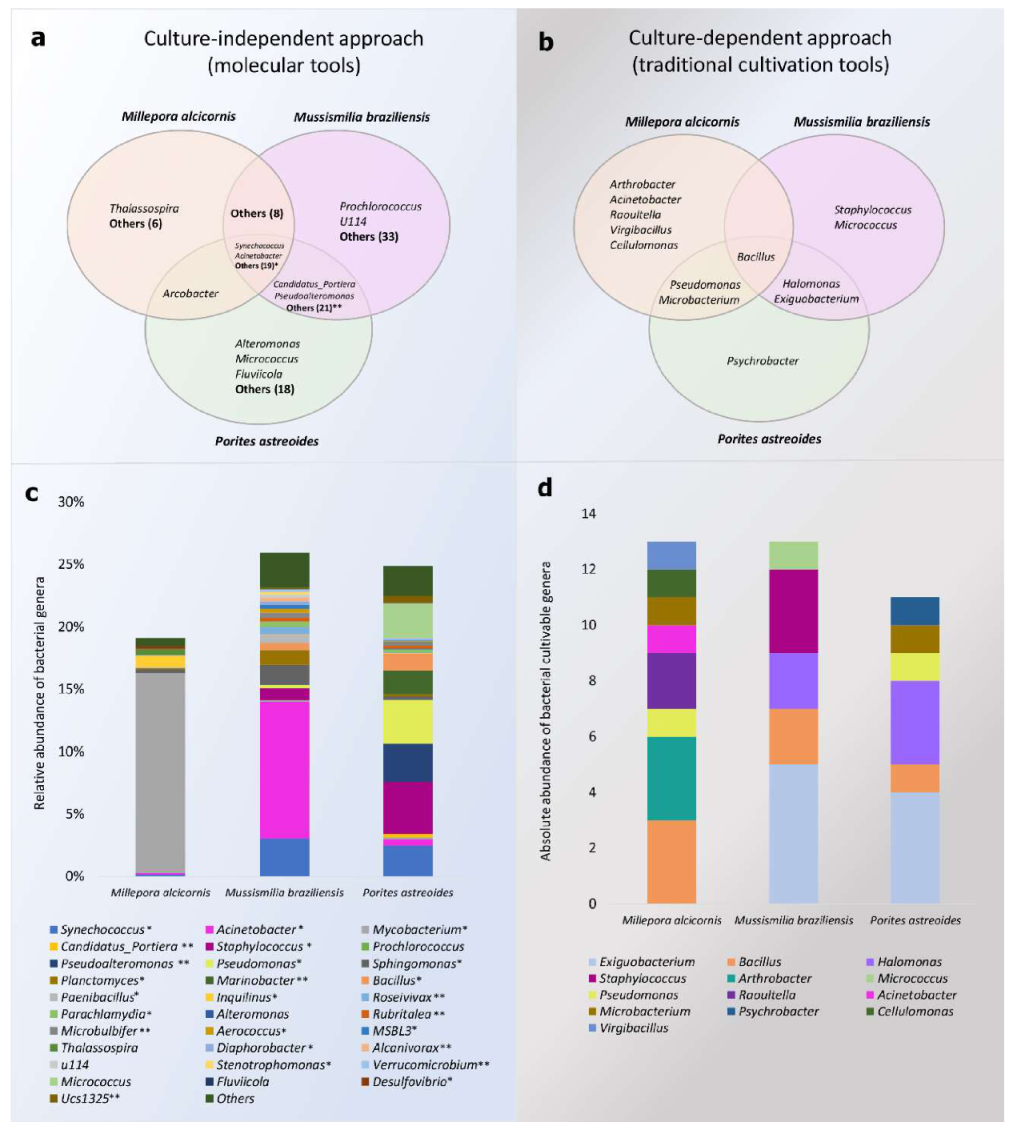
3.1. Alpha-Diversity and Microbiome Survey Provided by Molecular Tools
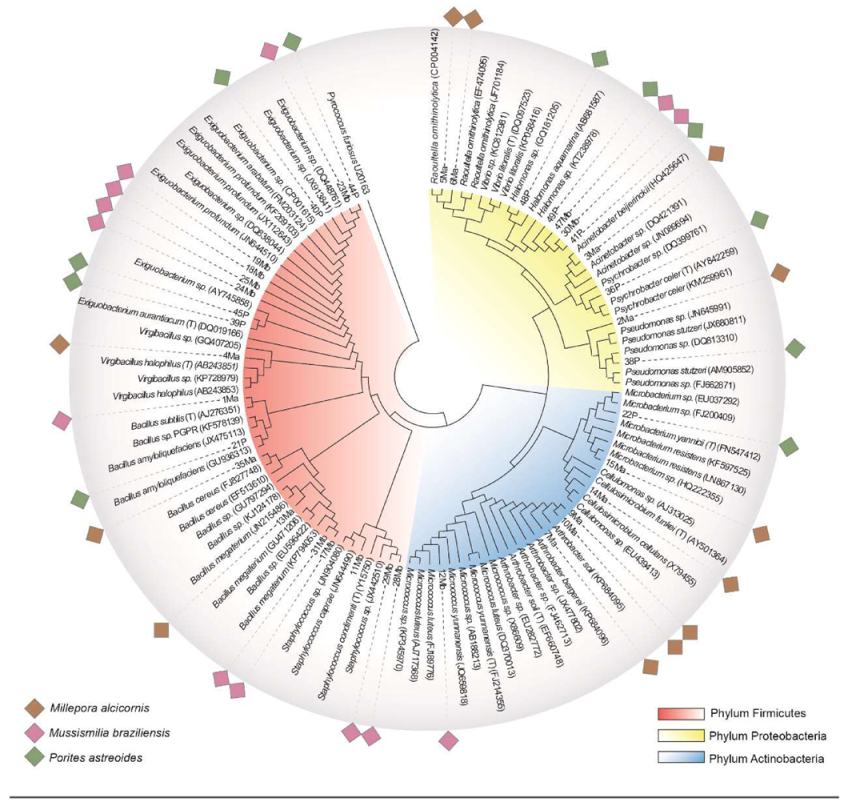
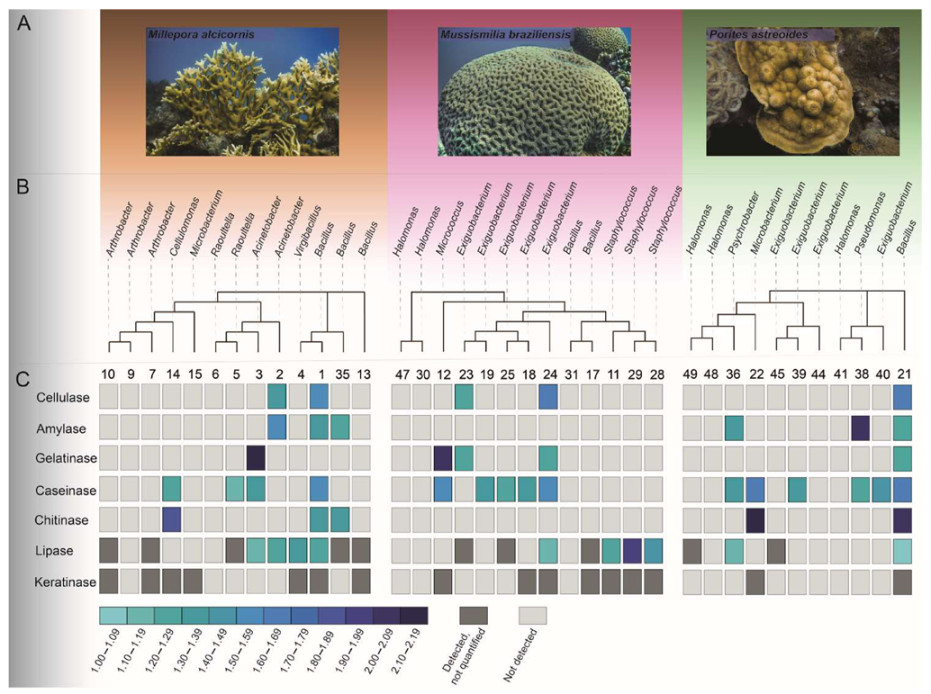
3.2. Isolation and Identification of CAM
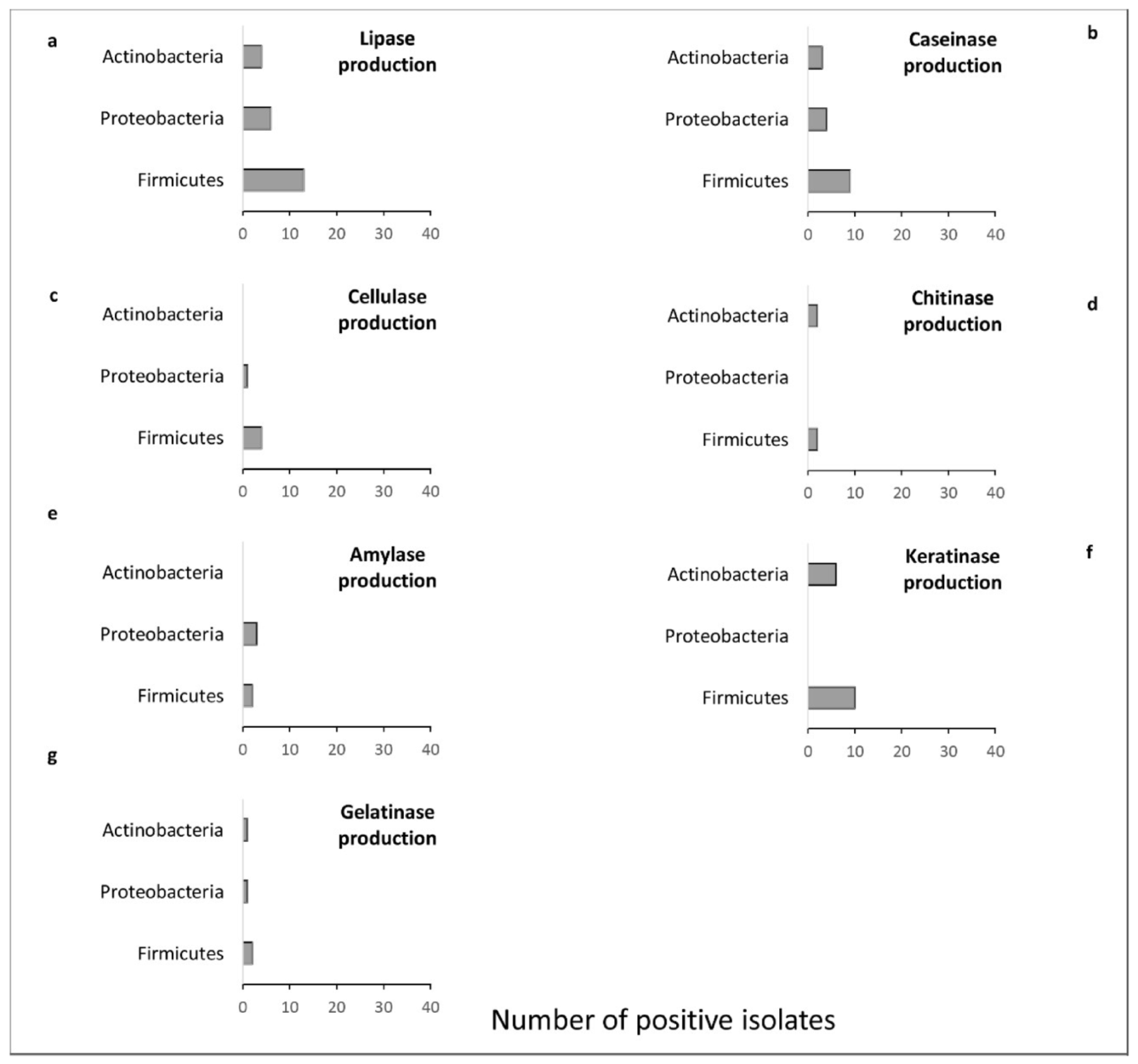
3.3. Enzymatic Potential of the Bacterial Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parte, S.; Sirisha, V.L.; D’Souza, J.S. Biotechnological applications of marine enzymes from algae, bacteria, fungi, and sponges. In Advances in Food and Nutrition Research; Academic Press: Cambridge, MA, USA, 2017; pp. 75–106. [Google Scholar]
- Chapman, J.; Ismail, A.E.; Dinu, C.Z. Industrial applications of enzymes: Recent advances, techniques, and outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef]
- Vogel, K. Analytics of Enzymes. In Enzymes in Human and Animal Nutrition; Elsevier: Amsterdam, The Netherlands, 2018; pp. 441–455. [Google Scholar]
- Peixoto, R.S.; Vermelho, A.B.; Rosado, A.S. Petroleum-degrading enzymes: Bioremediation and new prospects. Enzym. Res. 2011, 475193. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, N. Hydrolase-Catalyzed Promiscuous Reactions and Applications in Organic Synthesis. In Molecular Biotechnology; IntechOpen: London, UK, 2019; pp. 1–37. [Google Scholar]
- Kumar, V.; Sangwan, P.; Singh, D.; Gill, P.K. Global scenario of industrial enzyme market. In Industrial Enzymes: Trends, Scope and Relevance; Nova Science Publishers: Hauppauge, NY, USA, 2014; pp. 176–196. [Google Scholar]
- Kirk, O.; Borchert, T.V.; Fuglsang, C.C. Industrial Enzyme Applications. Curr. Opin. Biotechnol. 2002, 13, 345–351. [Google Scholar] [CrossRef]
- Lammle, K.; Zipper, H.; Breuer, M.; Hauer, B.; Buta, C.; Brunner, H.; Rupp, S. Identification of novel enzymes with different hydrolytic activities by metagenome expression cloning. J. Biotechnol. 2007, 127, 575–592. [Google Scholar] [CrossRef]
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial Enzymes: Industrial Progress in 21st Century. 3 Biotech 2016, 6, 174. [Google Scholar] [CrossRef]
- Fenical, W. New Pharmaceuticals from Marine Organisms. Trends Biotechnol. 1997, 15, 339–341. [Google Scholar] [CrossRef]
- Wijffels, R.H. Potential of Sponges and Microalgae for Marine Biotechnology. Trends Biotechnol. 2008, 26, 26–31. [Google Scholar] [CrossRef]
- Querellou, J.; Cadoret, J.-P.; Allen, M.J.; Collén, J. Marine Biotechnology. In Introduction to Marine Genomics; Springer: Dordrecht, The Netherlands, 2010; pp. 287–313. [Google Scholar]
- Ferrer, M.; Méndez-García, C.; Bargiela, R.; Chow, J.; Alonso, S.; García-Moyano, A.; Bjerga, G.E.K.; Steen, I.H.; Schwabe, T.; Blom, C.; et al. The INMARE Consortium, Decoding the ocean’s microbiological secrets for marine enzyme biodiscovery. FEMS Microbiol. Lett. 2019, 366, fny285. [Google Scholar] [CrossRef]
- Raj, Y.; Chauhan, V.S.A. Isolation, Characterization and Screening of Novel Antibiotic Producing Bacteria from Natural Habitats of Western Himalayas and Industrial Waste Soil Samples. Int. J. Comput. Syst. 2019, 3, 3282–3288. [Google Scholar]
- Salas-Ovilla, R.; Gálvez-López, D.; Vázquez-Ovando, A.; Salvador-Figueroa, M.; Rosas-Quijano, R. Isolation and Identification of Marine Strains of Stenotrophomona maltophilia with High Chitinolytic Activity. PeerJ 2019, 7, e6102. [Google Scholar] [CrossRef]
- Arora, N.K.; Mishra, J.; Mishra, V. Microbial Enzymes: Roles and Applications in Industries; Springer: Singapore, 2021. [Google Scholar]
- Singh, S.K.; Singh, M.K.; Singh, V.K.; Modi, A.; Jaiswal, P.; Rashmi, K.; Kumar, A. Microbial Enzymes and Their Exploitation in Remediation of Environmental Contaminants. In Microbe Mediated Remediation of Environmental Contaminants; Elsevier: Amsterdam, The Netherlands, 2021; pp. 59–71. [Google Scholar]
- Illanes, A.; Cauerhff, A.; Wilson, L.; Castro, G.R. Recent Trends in Biocatalysis Engineering. Bioresour. Technol. 2012, 115, 48–57. [Google Scholar] [CrossRef]
- Guerrand, D. Lipases Industrial Applications: Focus on Food and Agroindustries. OCL 2017, 24, D403. [Google Scholar] [CrossRef]
- Bonugli-Santos, R.C.; dos Santos Vasconcelos, M.R.; Passarini, M.R.Z.; Vieira, G.A.L.; Lopes, V.C.P.; Mainardi, P.H.; dos Santos, J.A.; de Azevedo Duarte, L.; Otero, I.V.R.; da Silva Yoshida, A.M.; et al. Marine-Derived Fungi: Diversity of Enzymes and Biotechnological Applications. Front. Microbiol. 2015, 6, 269. [Google Scholar] [CrossRef]
- Gomes, N.; Dasari, R.; Chandra, S.; Kiss, R.; Kornienko, A. Marine Invertebrate Metabolites with Anticancer Activities: Solutions to the “Supply Problem”. Mar. Drugs 2016, 14, 98. [Google Scholar] [CrossRef]
- Morlighem, R.L.; Huang, J.É.; Liao, C.; Braga Gomes, Q.; Daniel Pérez, P.; de Brandão Prieto-da-Silva, Á.; Ming-Yuen Lee, S.; Rádis-Baptista, G. The Holo-Transcriptome of the Zoantharian Protopalythoa variabilis (Cnidaria: Anthozoa): A Plentiful Source of Enzymes for Potential Application in Green Chemistry, Industrial and Pharmaceutical Biotechnology. Mar. Drugs 2018, 16, 207. [Google Scholar] [CrossRef]
- Barzkar, N. Marine Microbial Alkaline Protease: An Efficient and Essential Tool for Various Industrial Applications. Int. J. Biol. Macromol. 2020, 161, 1216–1229. [Google Scholar] [CrossRef]
- Zhang, J.; Jiang, L.; Chen, X.; Lv, K.; Basiony, M.; Zhu, G.; Karthik, L.; Ouyang, , L.; Zhang, L.; Liu, X. Recent advances in biotechnology for marine enzymes and molecules. Curr. Opin. Biotechnol. 2021, 69, 308–315. [Google Scholar] [CrossRef]
- Tello, E.; Castellanos, L.; Arevalo-Ferro, C.; Duque, C. Cembranoid Diterpenes from the Caribbean Sea Whip Eunicea knighti. J. Nat. Prod. 2009, 72, 1595–1602. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; McIntosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The Odyssey of Marine Pharmaceuticals: A Current Pipeline Perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef]
- Mayer, A.; Rodríguez, A.; Taglialatela-Scafati, O.; Fusetani, N. Marine Pharmacology in 2009–2011: Marine Compounds with Antibacterial, Antidiabetic, Antifungal, Anti-Inflammatory, Antiprotozoal, Antituberculosis, and Antiviral Activities; Affecting the Immune and Nervous Systems, and Other Miscellaneous Mechanisms of Action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar] [CrossRef]
- Blackall, L.L.; Wilson, B.; van Oppen, M.J.H. Coral—the World’s Most Diverse Symbiotic Ecosystem. Mol. Ecol. 2015, 24, 5330–5347. [Google Scholar] [CrossRef] [PubMed]
- Braun, D.R.; Chevrette, M.G.; Acharya, D.; Currie, C.R.; Rajski, S.R.; Ritchie, K.B.; Bugni, T.S. Complete Genome Sequence of Dietzia sp. Strain WMMA184, a Marine Coral-Associated Bacterium. Genome Announc. 2018, 6, e01582-17. [Google Scholar] [CrossRef] [PubMed]
- Sang, V.T.; Dat, T.T.H.; Vinh, L.B.; Cuong, L.C.V.; Oanh, P.T.T.; Ha, H.; Kim, Y.H.; Anh, H.L.T.; Yang, S.Y. Coral and Coral-Associated Microorganisms: A Prolific Source of Potential Bioactive Natural Products. Mar. Drugs 2019, 17, 468. [Google Scholar] [CrossRef] [PubMed]
- Sweet, M.; Villela, H.; Keller-Costa, T.; Costa, R.; Romano, S.; Bourne, D.; Cardenas, A.; Huggett, M.; Kerwin, A.; Kuek, F.; et al. Insights into the Cultured Bacterial Fraction of Corals. mSystems 2020, 6, e01249-20. [Google Scholar]
- Bosch, T.C.G.; McFall-Ngai, M.J. Metaorganisms as the New Frontier. Zoology 2011, 114, 185–190. [Google Scholar] [CrossRef]
- Putnam, H.M.; Stat, M.; Pochon, X.; Gates, R.D. Endosymbiotic Flexibility Associates with Environmental Sensitivity in Scleractinian Corals. Proc. R. Soc. B Biol. Sci. 2012, 279, 4352–4361. [Google Scholar] [CrossRef]
- Santoro, E.P.; Borges, R.M.; Espinoza, J.L.; Freire, M.; Messias, C.S.M.A.; Villela, H.D.M.; Pereira, L.M.; Vilela, C.L.S.; Rosado, J.G.; Cardoso, P.M.; et al. Coral Microbiome Manipulation Elicits Metabolic and Genetic Restructuring to Mitigate Heat Stress and Evade Mortality. Sci. Adv. 2021, 7, eabg3088. [Google Scholar] [CrossRef]
- Rohwer, F.; Seguritan, V.; Azam, F.; Knowlton, N. Diversity and Distribution of Coral-Associated Bacteria. Mar. Ecol. Prog. Ser. 2002, 243, 1–10. [Google Scholar] [CrossRef]
- Rosenberg, E.; Koren, O.; Reshef, L.; Efrony, R.; Zilber-Rosenberg, I. The Role of Microorganisms in Coral Health, Disease and Evolution. Nat. Rev. Microbiol. 2007, 5, 355–362. [Google Scholar] [CrossRef]
- Pollock, F.J.; McMinds, R.; Smith, S.; Bourne, D.G.; Willis, B.L.; Medina, M.; Thurber, R.V.; Zaneveld, J.R. Coral-Associated Bacteria Demonstrate Phylosymbiosis and Cophylogeny. Nat. Commun. 2018, 9, 4921. [Google Scholar] [CrossRef]
- Bourne, D.; Iida, Y.; Uthicke, S.; Smith-Keune, C. Changes in Coral-Associated Microbial Communities during a Bleaching Event. ISME J. 2007, 2, 350–363. [Google Scholar] [CrossRef]
- Ziegler, M.; Grupstra, C.G.B.; Barreto, M.M.; Eaton, M.; BaOmar, J.; Zubier, K.; Al-Sofyani, A.; Turki, A.J.; Ormond, R.; Voolstra, C.R. Coral Bacterial Community Structure Responds to Environmental Change in a Host-Specific Manner. Nat. Commun. 2019, 10, 3092. [Google Scholar] [CrossRef]
- Webster, N.S.; Reusch, T.B.H. Microbial Contributions to the Persistence of Coral Reefs. ISME J. 2017, 11, 2167–2174. [Google Scholar] [CrossRef]
- Leite, D.C.; Salles, J.F.; Calderon, E.N.; Castro, C.B.; Bianchini, A.; Marques, J.A.; Van Elsas, J.D.; Peixoto, R.S. Coral bacterial-core abundance and network complexity as proxies for anthropogenic pollution. Front. Microbiol. 2018, 9, 833. [Google Scholar] [CrossRef]
- Leite, D.C.; Leão, P.; Garrido, A.G.; Lins, U.; Santos, H.F.; Pires, D.O.; Castro, C.B.; van Elsas, J.D.; Zilberberg, C.; Rosado, A.S.; et al. Broadcast spawning coral Mussismilia hispida can vertically transfer its associated bacterial core. Front. Microbiol. 2017, 8, 176. [Google Scholar] [CrossRef]
- Modolon, F.; Barno, A.R.; Villela, H.D.M.; Peixoto, R.S. Ecological and Biotechnological Importance of Secondary Metabolites Produced by Coral-associated Bacteria. J. Appl. Microbiol. 2020, 129, 1441–1457. [Google Scholar] [CrossRef]
- Villela, H.D.M.; Vilela, C.L.S.; Assis, J.M.; Varona, N.; Burke, C.; Coil, D.A.; Eisen, J.A.; Peixoto, R.S. Prospecting Microbial Strains for Bioremediation and Probiotics Development for Metaorganism Research and Preservation. J. Vis. Exp. 2019, 2019, e60238. [Google Scholar] [CrossRef]
- Morrow, K.M.; Moss, A.G.; Chadwick, N.E.; Liles, M.R. Bacterial Associates of Two Caribbean Coral Species Reveal Species-Specific Distribution and Geographic Variability. Appl. Environ. Microbiol. 2012, 78, 6438–6449. [Google Scholar] [CrossRef]
- Neave, M.J.; Rachmawati, R.; Xun, L.; Michell, C.T.; Bourne, D.G.; Apprill, A.; Voolstra, C.R. Differential Specificity between Closely Related Corals and Abundant Endozoicomonas Endosymbionts across Global Scales. ISME J. 2016, 11, 186–200. [Google Scholar] [CrossRef]
- Peixoto, R.S.; Rosado, P.M.; Leite, D.C.D.A.; Rosado, A.S.; Bourne, D.G. Beneficial Microorganisms for Corals (BMC): Proposed Mechanisms for Coral Health and Resilience. Front. Microbiol. 2017, 8, 341. [Google Scholar] [CrossRef]
- Peixoto, R.S.; Sweet, M.; Villela, H.D.; Cardoso, P.; Thomas, T.; Voolstra, C.R.; Høj, L.; Bourne, D.G. Coral probiotics: Premise, promise, prospects. Annu. Rev. Anim. Biosci. 2021, 9, 265–288. [Google Scholar] [CrossRef]
- Silva, D.P.; Villela, H.D.; Santos, H.F.; Duarte, G.A.; Ribeiro, J.R.; Ghizelini, A.M.; Vilela, C.L.; Rosado, P.M.; Fazolato, C.S.; Santoro, E.P.; et al. Multi-domain probiotic consortium as an alternative to chemical remediation of oil spills at coral reefs and adjacent sites. Microbiome 2021, 9, 1–19. [Google Scholar] [CrossRef]
- do Carmo, F.L.; dos Santos, H.F.; Martins, E.F.; van Elsas, J.D.; Rosado, A.S.; Peixoto, R.S. Bacterial Structure and Characterization of Plant Growth Promoting and Oil Degrading Bacteria from the Rhizospheres of Mangrove Plants. J. Microbiol. 2011, 49, 535–543. [Google Scholar] [CrossRef]
- Santos, H.F.; Duarte, G.A.S.; da Costa Rachid, C.T.; Chaloub, R.M.; Calderon, E.N.; de Barros Marangoni, L.F.; Bianchini, A.; Nudi, A.H.; Do Carmo, F.L.; Van Elsas, J.D.; et al. Impact of oil spills on coral reefs can be reduced by bioremediation using probiotic microbiota. Sci. Rep. 2015, 5, 1–11. [Google Scholar]
- Hoyt, J.R.; Langwig, K.E.; White, J.P.; Kaarakka, H.M.; Redell, J.A.; Parise, K.L.; Frick, W.F.; Foster, J.T.; Kilpatrick, A.M. Field trial of a probiotic bacteria to protect bats from white-nose syndrome. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Becker, M.H.; Harris, R.N.; Minbiole, K.P.; Schwantes, C.R.; Rollins-Smith, L.A.; Reinert, L.K.; Brucker, R.M.; Domangue, R.J.; Gratwicke, B. Towards a better understanding of the use of probiotics for preventing chytridiomycosis in Panamanian golden frogs. Ecohealth 2011, 8, 501–506. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Labes, A.; Wiese, J. Bio-Mining the Microbial Treasures of the Ocean: New Natural Products. Biotechnol. Adv. 2011, 29, 468–482. [Google Scholar] [CrossRef]
- Radjasa, O.K.; Khoeri, M.M.; Darusallam, C.C.; Trimasanto, H.; Sudoyo, H. Bacterial Symbionts of Reef Invertebrates: Screening for Anti-Pathogenic Bacteria Activity. Biodiversity 2013, 14, 80–86. [Google Scholar] [CrossRef]
- Rizzo, C.; Lo Giudice, A. Marine Invertebrates: Underexplored Sources of Bacteria Producing Biologically Active Molecules. Diversity 2018, 10, 52. [Google Scholar] [CrossRef]
- El-Gendy, M.M.A. Keratinase Production by Endophytic Penicillium spp. Morsy1 Under Solid-State Fermentation Using Rice Straw. Appl. Biochem. Biotechnol. 2009, 162, 780–794. [Google Scholar] [CrossRef]
- Bakkiyaraj, D.; Sivasankar, C.; Pandian, S.K. Anti-Pathogenic Potential of Coral Associated Bacteria Isolated from Gulf of Mannar Against Pseudomonas aeruginosa. Indian J. Microbiol. 2012, 53, 111–113. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mabrouk, M.E.; Youssif, E.M.; Sabry, S.A. Biosurfactant production by a newly isolated soft coral-associated marine Bacillus sp. E34: Statistical optimization and characterization. Life Sci. J. 2014, 11, 756–768. [Google Scholar]
- Ahila, N.K.; Prakash, S.; Manikandan, B.; Ravindran, J.; Prabhu, N.M.; Kannapiran, E. Bio-Prospecting of Coral (Porites lutea) Mucus Associated Bacteria, Palk Bay Reefs, Southeast Coast of India. Microb. Pathog. 2017, 113, 113–123. [Google Scholar] [CrossRef] [PubMed]
- Ledoux, J.B.; Antunes, A. Beyond the beaten path: Improving natural products bioprospecting using an eco-evolutionary framework—The case of the octocorals. Crit. Rev. Biotechnol. 2018, 38, 184–198. [Google Scholar] [CrossRef]
- Knowlton, N.; Grottoli, A.G.; Kleypas, J.; Obura, D.; Corcoran, E.; de Goeij, J.; Felis, T.; Harding, S.; Mayfield, A.; Miller, M.; et al. Rebuilding Coral Reefs: A Decadal Grand Challenge. Int. Coral Reef Soc. Future Earth Coasts 2021, 56. [Google Scholar] [CrossRef]
- Seoane, J.C.S.; Arantes, R.C.M.; Castro, C.B.; Tedesco, E.C.; Lopes, L.S. Atlas Físico do Parque Municipal Marinho do Recife de Fora; Projeto Coral Vivo: Rio de Janeiro, Brasil, 2008. [Google Scholar]
- Paula, Y.C.D.; Schiavetti, A.; Sampaio, C.L.; Calderon, E. The effects of fish feeding by visitors on reef fish in a Marine Protected Area open to tourism. Biota Neotrop. 2018, 18. [Google Scholar] [CrossRef]
- Costa Jr, O.S.; Attrill, M.J.; Pedrini, A.G.; De-Paula, J.C. Spatial and seasonal distribution of seaweeds on coral reefs from Southern Bahia, Brazil. Bot. Mar. 2002, 45, 346–355. [Google Scholar] [CrossRef]
- Voolstra, C.R.; Quigley, K.M.; Davies, S.W.; Parkinson, J.E.; Peixoto, R.S.; Aranda, M.; Baker, A.C.; Barno, A.R.; Barshis, D.J.; Benzoni, F.; et al. Consensus guidelines for advancing coral holobiont genome and specimen voucher deposition. Front. Mar. Sci. 2021, 1029. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global Patterns of 16S RRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2010, 108 (Suppl. 1), 4516–4522. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef]
- Vilela, C.L.S.; Villela, H.D.M.; Duarte, G.A.; Santoro, E.P.; Rachid, C.T.; Peixoto, R.S. Estrogen induces shift in abundances of specific groups of the coral microbiome. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glockner, F.O. SILVA: A Comprehensive Online Resource for Quality Checked and Aligned Ribosomal RNA Sequence Data Compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Schleif, R.F.; Wensink, P.C. Practical Methods in Molecular Biology; Springer Science & Business Media: New York, NY, USA, 2012. [Google Scholar]
- Hankin, L.; Zucker, M.; Sands, D.C. Improved Solid Medium for the Detection and Enumeration of Pectolytic Bacteria. Appl. Microbiol. 1971, 22, 205–209. [Google Scholar] [CrossRef]
- Carrim, A.J.I.; Barbosa, E.C.; Vieira, J.D.G. Enzymatic activity of endophytic bacterial isolates of Jacaranda decurrens Cham. (carobinha-do-campo). Braz. Arch. Biol. Technol. 2006, 49, 353–359. [Google Scholar] [CrossRef]
- de Castro, A.L.M.; Vollú, R.E.; Peixoto, R.S.; Grigorevski-Lima, A.L.; Coelho, R.R.R.; Bon, E.P.S.; Rosado, A.S.; Seldin, L. Cellulolytic potential of a novel strain of Paenibacillus sp. isolated from the armored catfish Parotocinclus maculicauda Gut. Braz. J. Microbiol. 2011, 42, 1608–1615. [Google Scholar] [CrossRef]
- Hankin, L.; Anagnostakis, S.L. The Use of Solid Media for Detection of Enzyme Production by Fungi. Mycologia 1975, 67, 597. [Google Scholar] [CrossRef]
- Williams, S.T.; Goodfellow, M.; Alderson, G.; Wellington, E.M.H.; Sneath, P.H.A.; Sackin, M.J. Numerical Classification of Streptomyces and Related Genera. Microbiology 1983, 129, 1743–1813. [Google Scholar] [CrossRef]
- Bairagi, A.; Ghosh, K.S.; Sen, S.K.; Ray, A.K. Enzyme producing bacterial flora isolated from fish digestive tracts. Aquac. Int. 2002, 10, 109–121. [Google Scholar] [CrossRef]
- Hsu, S.C.; Lockwood, J.L. Powdered Chitin Agar as a Selective Medium for Enumeration of Actinomycetes in Water and Soil. Appl. Microbiol. 1975, 29, 422–426. [Google Scholar] [CrossRef]
- Chaturvedi, V.; Bhange, K.; Bhatt, R.; Verma, P. Production of keratinases using chicken feathers as substrate by a novel multifunctional strain of Pseudomonas stutzeri and its dehairing application. Biocatal. Agric. Biotechnol. 2014, 3, 167–174. [Google Scholar] [CrossRef]
- Lane, D.J. 16S/23S rRNA sequencing. In Nucleic Acid Techniques in Bacterial Systematics; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Hall, T. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Cole, J.R.; Wang, Q.; Cardenas, E.; Fish, J.; Chai, B.; Farris, R.J.; Kulam-Syed-Mohideen, A.S.; McGarrell, D.M.; Marsh, T.; Garrity, G.M.; et al. The Ribosomal Database Project: Improved Alignments and New Tools for RRNA Analysis. Nucleic Acids Res. 2009, 37, D141–D145. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for MacOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M. Estimation of the Number of Nucleotide Substitutions in the Control Region of Mitochondrial DNA in Humans and Chimpanzees. Mol. Biol. Evol. 1993, 10, 512–526. [Google Scholar] [CrossRef]
- Nakamura, K.; Iizuka, R.; Nishi, S.; Yoshida, T.; Hatada, Y.; Takaki, Y.; Iguchi, A.; Yoon, D.H.; Sekiguchi, T.; Shoji, S.; et al. Culture-Independent Method for Identification of Microbial Enzyme-Encoding Genes by Activity-Based Single-Cell Sequencing Using a Water-in-Oil Microdroplet Platform. Sci. Rep. 2016, 6, 22259. [Google Scholar] [CrossRef]
- Vivero, R.J.; Mesa, G.B.; Robledo, S.M.; Herrera, C.X.M.; Cadavid-Restrepo, G. Enzymatic, Antimicrobial, and Leishmanicidal Bioactivity of Gram-Negative Bacteria Strains from the Midgut of Lutzomyia evansi, an Insect Vector of Leishmaniasis in Colombia. Biotechnol. Rep. 2019, 24, e00379. [Google Scholar] [CrossRef]
- De Castro, A.P.; Araújo, S.D., Jr.; Reis, A.M.M.; Moura, R.L.; Francini-Filho, R.B.; Pappas, G., Jr.; Rodrigues, T.B.; Thompson, F.L.; Krüger, R.H. Bacterial Community Associated with Healthy and Diseased Reef Coral Mussismilia hispida from Eastern Brazil. Microb. Ecol. 2010, 59, 658–667. [Google Scholar] [CrossRef]
- Hernandez-Agreda, A.; Gates, R.D.; Ainsworth, T.D. Defining the Core Microbiome in Corals’ Microbial Soup. Trends Microbiol. 2017, 25, 125–140. [Google Scholar] [CrossRef]
- Arotsker, L.; Siboni, N.; Ben-Dov, E.; Kramarsky-Winter, E.; Loya, Y.; Kushmaro, A. Vibrio sp. as a potentially important member of the Black Band Disease (BBD) consortium in Favia sp. corals. FEMS Microbiol. Ecol. 2009, 70, 515–524. [Google Scholar] [CrossRef]
- Kushmaro, A.; Banin, E.; Loya, Y.; Stackebrandt, E.; Rosenberg, E. Vibrio shiloi sp. Nov., the Causative Agent of Bleaching of the Coral Oculina patagonica. Int. J. Syst. Evol. Microbiol. 2001, 51, 1383–1388. [Google Scholar] [CrossRef]
- Sabdono, A.; Wijayanti, D.P.; Sarjito, S. Antipathogenic Activity of Bacteria Associated with Acroporid Corals Against Black Band Disease of Karimunjawa, Indonesia. Res. J. Microbiol. 2017, 12, 154–160. [Google Scholar] [CrossRef][Green Version]
- Matsumiya, Y.; Wakita, D.; Kimura, A.; Sanpa, S.; Kubo, M. Isolation and Characterization of a Lipid-Degrading Bacterium and Its Application to Lipid-Containing Wastewater Treatment. J. Biosci. Bioeng. 2007, 103, 325–330. [Google Scholar] [CrossRef]
- Liu, X.; Ren, B.; Chen, M.; Wang, H.; Kokare, C.R.; Zhou, X.; Wang, J.; Dai, H.; Song, F.; Liu, M.; et al. Production and Characterization of a Group of Bioemulsifiers from the Marine Bacillus velezensis Strain H3. Appl. Microbiol. Biotechnol. 2010, 87, 1881–1893. [Google Scholar] [CrossRef]
- Sabat, S.; Murthy, V.K.; Pavithra, M.; Mayur, P.; Chandavar, A. Production and characterisation of extracellular lipase from Bacillus stearothermophilus MTCC 37 under different fermentation conditions. Int. J. Eng. Res. Appl. 2012, 2, 1775–1781. [Google Scholar]
- Mahale, P.K.; Desai, S.V.; Hombalimath, V.S.; Achappa, S. Isolation, screening and characterization of lipase producing strain from oil contaminated soil of Hubballi, Karnataka. Int. J. Basic Appl. Biol. 2014, 2, 198–201. [Google Scholar]
- Gricajeva, A.; Bendikienė, V.; Kalėdienė, L. Lipase of Bacillus stratosphericus L1: Cloning, Expression and Characterization. Int. J. Biol. Macromol. 2016, 92, 96–104. [Google Scholar] [CrossRef]
- Zhao, J.; Liu, S.; Gao, Y.; Ma, M.; Yan, X.; Cheng, D.; Wan, D.; Zeng, Z.; Yu, P.; Gong, D. Characterization of a novel lipase from Bacillus licheniformis NCU CS-5 for applications in detergent industry and biodegradation of 2, 4-D butyl ester. Int. J. Biol. Macromol. 2021, 176, 126–136. [Google Scholar] [CrossRef]
- Khunt, M.; Pandhi, N.; Rana, A. Media Optimization for Lipase from Halomonas salina Mk-23, Moderate Halophiles Isolated from Wild Ass Excreta. J. Pure Appl. Microbiol. 2012, 6, 845–850. [Google Scholar]
- Li, C.-Y.; Cheng, C.-Y.; Chen, T.-L. Fed-Batch Production of Lipase by Acinetobacter radioresistens Using Tween 80 as the Carbon Source. Biochem. Eng. J. 2004, 19, 25–31. [Google Scholar] [CrossRef]
- Li, C.-Y.; Chen, S.-J.; Cheng, C.-Y.; Chen, T.-L. Production of Acinetobacter radioresistens Lipase with Repeated Fed-Batch Culture. Biochem. Eng. J. 2005, 25, 195–199. [Google Scholar] [CrossRef]
- Javed, S.; Azeem, F.; Hussain, S.; Rasul, I.; Siddique, M.H.; Riaz, M.; Afzal, M.; Kouser, A.; Nadeem, H. Bacterial Lipases: A Review on Purification and Characterization. Prog. Biophys. Mol. Biol. 2018, 132, 23–34. [Google Scholar] [CrossRef]
- Patel, R.K.; Shah, R.K.; Prajapati, V.S.; Patel, K.C.; Trivedi, U.B. Draft Genome Analysis of Acinetobacter indicus Strain UBT1, an Efficient Lipase and Biosurfactant Producer. Curr. Microbiol. 2021, 78, 1238–1244. [Google Scholar] [CrossRef]
- Phukon, L.C.; Chourasia, R.; Kumari, M.; Godan, T.K.; Sahoo, D.; Parameswaran, B.; Rai, A.K. Production and characterisation of lipase for application in detergent industry from a novel Pseudomonas helmanticensis HS6. Bioresour. Technol. 2020, 309, 123352. [Google Scholar] [CrossRef]
- Sarmah, N.; Revathi, D.; Sheelu, G.; Yamuna Rani, K.; Sridhar, S.; Mehtab, V.; Sumana, C. Recent advances on sources and industrial applications of lipases. Biotechnol. Prog. 2018, 34, 5–28. [Google Scholar] [CrossRef]
- Filho, D.G.; Silva, A.G.; Guidini, C.Z. Lipases: Sources, Immobilization Methods, and Industrial Applications. Appl. Microbiol. Biotechnol. 2019, 103, 7399–7423. [Google Scholar] [CrossRef]
- Nielsen, T. Industrial Application Possibilities for Lipase. Fette Seifen Anstrichm. 1985, 87, 15–19. [Google Scholar] [CrossRef]
- Navvabi, A.; Razzaghi, M.; Fernandes, P.; Karami, L.; Homaei, A. Novel Lipases Discovery Specifically from Marine Organisms for Industrial Production and Practical Applications. Process. Biochem. 2018, 70, 61–70. [Google Scholar] [CrossRef]
- Kiran, G.S.; Shanmughapriya, S.; Jayalakshmi, J.; Selvin, J.; Gandhimathi, R.; Sivaramakrishnan, S.; Arunkumar, M.; Thangavelu, T.; Natarajaseenivasan, K. Optimization of Extracellular Psychrophilic Alkaline Lipase Produced by Marine Pseudomonas sp. (MSI057). Bioprocess. Biosyst. Eng. 2008, 31, 483–492. [Google Scholar] [CrossRef]
- Hassan, S.W.M.; Abd El Latif, H.H.; Ali, S.M. Production of Cold-Active Lipase by Free and Immobilized Marine Bacillus cereus HSS: Application in Wastewater Treatment. Front. Microbiol. 2018, 9, 2377. [Google Scholar] [CrossRef]
- Verma, S.; Meghwanshi, G.K.; Kumar, R. Current perspectives for microbial lipases from extremophiles and metagenomics. Biochimie 2021, 182, 26–36. [Google Scholar] [CrossRef]
- Gupta, R.; Beg, Q.K.; Lorenz, P. Bacterial Alkaline Proteases: Molecular Approaches and Industrial Applications. Appl. Microbiol. Biotechnol. 2002, 59, 15–32. [Google Scholar] [CrossRef]
- Hammami, A.; Hamdi, M.; Abdelhedi, O.; Jridi, M.; Nasri, M.; Bayoudh, A. Surfactant- and Oxidant-Stable Alkaline Proteases from Bacillus invictae: Characterization and Potential Applications in Chitin Extraction and as a Detergent Additive. Int. J. Biol. Macromol. 2017, 96, 272–281. [Google Scholar] [CrossRef]
- Thakur, N.; Goyal, M.; Sharma, S.; Kumar, D. Proteases: Industrial applications and approaches used in strain improvement. Biol. Forum–An. Int. J. 2018, 10, 158–167. [Google Scholar]
- Barzkar, N.; Homaei, A.; Hemmati, R.; Patel, S. Thermostable Marine Microbial Proteases for Industrial Applications: Scopes and Risks. Extremophiles 2018, 22, 335–346. [Google Scholar] [CrossRef]
- Vojcic, L.; Jakob, F.; Martinez, R.; Hellmuth, H.; O’Connell, T.; Mühl, H.; Lorenz, M.G.; Schwaneberg, U. Engineering Proteases for Industrial Applications. In Applied Biocatalysis: From Fundamental Science to Industrial Applications; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2016; pp. 101–119. [Google Scholar]
- Suriya, J.; Bharathiraja, S.; Krishnan, M.; Manivasagan, P.; Kim, S.-K. Extremozymes from Marine Actinobacteria. In Marine Enzymes Biotechnology: Production and Industrial Applications, Part II—Marine Organisms Producing Enzymes; Elsevier: Amsterdam, The Netherlands, 2016; pp. 43–66. [Google Scholar]
- Hamza, H.M.; Ali, S.M.; Hassan, H.G. Partial purification of gelatinase enzyme from local isolate of Brevibacillus laterosporus. Natl. J. 2006, 23, 437–442. [Google Scholar]
- Sameena, F.M.; Radha, P. Fish Waste Gelatin: A Renewable Substrate for Production of Gelatinase by Serratia Marcescens MF599353. J. Solid Waste Technol. Manag. 2020, 46, 168–177. [Google Scholar] [CrossRef]
- Sai-Ut, S.; Benjakul, S.; Sumpavapol, P. Gelatinolytic Enzymes from Bacillus amyloliquefaciens Isolated from Fish Docks: Characteristics and Hydrolytic Activity. Food Sci. Biotechnol. 2013, 22, 1015–1021. [Google Scholar] [CrossRef]
- Yuivar, Y.; Alcaino, J.; Cifuentes, V.; Baeza, M. Characterization of Gelatinase Produced by Antarctic mrakia sp. J. Basic Microbiol. 2019, 59, 846–852. [Google Scholar] [CrossRef]
- Chimetto, L.A.; Cleenwerck, I.; Thompson, C.C.; Brocchi, M.; Willems, A.; De Vos, P.; Thompson, F.L. Photobacterium jeanii sp. nov., Isolated from Corals and Zoanthids. Int. J. Syst. Evol. Microbiol. 2010, 60, 2843–2848. [Google Scholar] [CrossRef]
- Yoshizawa, S.; Tsuruya, Y.; Fukui, Y.; Sawabe, T.; Yokota, A.; Kogure, K.; Higgins, M.; Carson, J.; Thompson, F.L. Vibrio jasicida sp. nov., a Member of the Harveyi Clade, Isolated from Marine Animals (Packhorse Lobster, Abalone and Atlantic Salmon). Int. J. Syst. Evol. Microbiol. 2012, 62 Pt 8, 1864–1870. [Google Scholar] [CrossRef]
- Ramlath, L.; Keerthana, P.P.; Safvana Fathima, P.; Mashhoor, K. Bacteria from Coral Ecosystem of Kiltan Island, Lakshadweep: Resource for Hydrolytic Enzymes. Int. J. Cell Sci. Biotechnol. 2018, 7, 1–9. [Google Scholar]
- Ben-Haim, Y.; Thompson, F.L.; Thompson, C.C.; Cnockaert, M.C.; Hoste, B.; Swings, J.; Rosenberg, E. Vibrio coralliilyticus sp. nov., a Temperature-Dependent Pathogen of the Coral Pocillopora damicornis. Int. J. Syst. Evol. Microbiol. 2003, 53, 309–315. [Google Scholar] [CrossRef]
- Thompson, F.L.; Barash, Y.; Sawabe, T.; Sharon, G.; Swings, J.; Rosenberg, E. Thalassomonas loyana sp. nov., a Causative Agent of the White Plague-like Disease of Corals on the Eilat Coral Reef. Int. J. Syst. Evol. Microbiol. 2006, 56, 365–368. [Google Scholar] [CrossRef]
- Stetler-Stevenson, W.G.; Aznavoorian, S.; Liotta, L.A. Tumor Cell Interactions with the Extracellular Matrix During Invasion and Metastasis. Annu. Rev. Cell Biol. 1993, 9, 541–573. [Google Scholar] [CrossRef]
- Balan, S.S.; Nethaji, R.; Sankar, S.; Jayalakshmi, S. Production of Gelatinase Enzyme from Bacillus Spp Isolated from the Sediment Sample of Porto Novo Coastal Sites. Asian Pac. J. Trop. Biomed. 2012, 2, S1811–S1816. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, F.; Wang, L.; Xie, C.; Wu, P.; Du, S.; Zhou, S.; Sun, Z.; Liu, Q.; Yu, L.; et al. Enhanced and Prolonged Antitumor Effect of Salinomycin-Loaded Gelatinase-Responsive Nanoparticles via Targeted Drug Delivery and Inhibition of Cervical Cancer Stem Cells. Int. J. Nanomed. 2020, 15, 1283–1295. [Google Scholar] [CrossRef]
- Mohan, R.; Chintala, S.K.; Jung, J.C.; Villar, W.V.L.; McCabe, F.; Russo, L.A.; Lee, Y.; McCarthy, B.E.; Wollenberg, K.R.; Jester, J.V.; et al. Matrix Metalloproteinase Gelatinase B (MMP-9) Coordinates and Effects Epithelial Regeneration. J. Biol. Chem. 2002, 277, 2065–2072. [Google Scholar] [CrossRef]
- Shulga, A.P.; Lamash, N.E. Proteinases with gelatinase activity and their role in ambulacrum regeneration in holothurians Eupentacta fraudatrix (D’yakonov and Baranova, 1958) and Cucumaria japonica (Semper, 1868) (Echinodermata: Holothuroidea). Russ. J. Mar. Biol. 2020, 46, 461–471. [Google Scholar] [CrossRef]
- Robyt, J.F.; Ackerman, R.J. Isolation, Purification, and Characterization of a Maltotetraose-Producing Amylase from Pseudomonas stutzeri. Arch. Biochem. Biophys. 1971, 145, 105–114. [Google Scholar] [CrossRef]
- Dutta, P.; Deb, A.; Majumdar, S. Optimization of the Medium for the Production of Extracellular Amylase by The Pseudomonas stutzeri ISL B5 Isolated from Municipal Solid Waste. Int. J. Microbiol. 2016, 2016, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Jin, T.; Xie, X.; Ban, X.; Li, C.; Hong, Y.; Cheng, L.; Gu, Z.; Li, Z. Structure of Maltotetraose-Forming Amylase from Pseudomonas saccharophila STB07 Provides Insights into Its Product Specificity. Int. J. Biol. Macromol. 2020, 154, 1303–1313. [Google Scholar] [CrossRef]
- Safari, M.; Kari Dolatabad, H.; Ndu, U.; Rivera, N.A. Protective Effect of Pseudomonas Spp. Isolates and Zinc on Seed Germination and β-Amylase Activity in Wheat Cultivars under Cadmium Stress. Acta Physiol. Plant. 2020, 42, 1–10. [Google Scholar] [CrossRef]
- Rosado, P.M.; Leite, D.C.A.; Duarte, G.A.S.; Chaloub, R.M.; Jospin, G.; Nunes da Rocha, U.; Saraiva, J.P.; Dini-Andreote, F.; Eisen, J.A.; Bourne, D.G.; et al. Marine Probiotics: Increasing Coral Resistance to Bleaching through Microbiome Manipulation. ISME J. 2018, 13, 921–936. [Google Scholar] [CrossRef]
- Douglas, N.L.; Mullen, K.M.; Talmage, S.C.; Harvell, C.D. Exploring the Role of Chitinolytic Enzymes in the Sea Fan Coral, Gorgonia ventalina. Mar. Biol. 2006, 150, 1137–1144. [Google Scholar] [CrossRef]
- Kumar, M.N.R. A review of chitin and chitosan applications. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar] [CrossRef]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Stoykov, Y.M.; Pavlov, A.I.; Krastanov, A.I. Chitinase Biotechnology: Production, Purification, and Application. Eng. Life Sci. 2014, 15, 30–38. [Google Scholar] [CrossRef]
- Nayak, S.K.; Nayak, S.; Mohanty, S.; Sundaray, J.K.; Mishra, B.B. Microbial Chitinases and Their Applications: An Overview. In Environmental and Agricultural Microbiology: Applications for Sustainability; Wiley: Hoboken, NJ, USA, 2021; pp. 313–340. [Google Scholar]
- Yoshioka, Y.; Tanabe, T.; Iguchi, A. The Presence of Genes Encoding Enzymes That Digest Carbohydrates in Coral Genomes and Analysis of Their Activities. PeerJ 2017, 5, e4087. [Google Scholar] [CrossRef]
- Brandelli, A. Bacterial Keratinases: Useful Enzymes for Bioprocessing Agroindustrial Wastes and Beyond. Food Bioprocess. Technol. 2007, 1, 105–116. [Google Scholar] [CrossRef]
- Nnolim, N.E.; Udenigwe, C.C.; Okoh, A.I.; Nwodo, U.U. Microbial keratinase: Next generation green catalyst and prospective applications. Front. Microbiol. 2020, 11, 3280. [Google Scholar] [CrossRef]
- Hassan, M.A.; Abol-Fotouh, D.; Omer, A.M.; Tamer, T.M.; Abbas, E. Comprehensive insights into microbial keratinases and their implication in various biotechnological and industrial sectors: A review. Int. J. Biol. Macromol. 2020, 154, 567–583. [Google Scholar] [CrossRef]
- Brandelli, A.; Daroit, D.J.; Riffel, A. Biochemical Features of Microbial Keratinases and Their Production and Applications. Appl. Microbiol. Biotechnol. 2009, 85, 1735–1750. [Google Scholar] [CrossRef]
- Lin, X.; Lee, C.-G.; Casale, E.S.; Shih, J.C.H. Purification and Characterization of a Keratinase from a Feather-Degrading Bacillus licheniformis Strain. Appl. Environ. Microbiol. 1992, 58, 3271–3275. [Google Scholar] [CrossRef]
- Herzog, B.; Overy, D.P.; Haltli, B.; Kerr, R.G. Discovery of Keratinases Using Bacteria Isolated from Marine Environments. Syst. Appl. Microbiol. 2016, 39, 49–57. [Google Scholar] [CrossRef]
- Arokiyaraj, S.; Varghese, R.; Ali Ahmed, B.; Duraipandiyan, V.; Al-Dhabi, N.A. Optimizing the Fermentation Conditions and Enhanced Production of Keratinase from Bacillus cereus Isolated from Halophilic Environment. Saudi J. Biol. Sci. 2019, 26, 378–381. [Google Scholar] [CrossRef]
- Bhange, K.; Chaturvedi, V.; Bhatt, R. Potential Biofilm Dispersal by a Partially Purified Keratinase Produced by Stenotrophomonas maltophilia Strain Kb2. Biocatal. Agric. Biotechnol. 2015, 4, 801–805. [Google Scholar] [CrossRef]
- Steinert, G.; Whitfield, S.; Taylor, M.W.; Thoms, C.; Schupp, P.J. Application of Diffusion Growth Chambers for the Cultivation of Marine Sponge-Associated Bacteria. Mar. Biotechnol. 2014, 16, 594–603. [Google Scholar] [CrossRef]
- Berdy, B.; Spoering, A.L.; Ling, L.L.; Epstein, S.S. In Situ Cultivation of Previously Uncultivable Microorganisms Using the Ichip. Nat. Protoc. 2017, 12, 2232–2242. [Google Scholar] [CrossRef]
- Palma Esposito, F.; Ingham, C.J.; Hurtado-Ortiz, R.; Bizet, C.; Tasdemir, D.; de Pascale, D. Isolation by Miniaturized Culture Chip of an Antarctic Bacterium Aequorivita Sp. with Antimicrobial and Anthelmintic Activity. Biotechnol. Rep. 2018, 20, e00281. [Google Scholar] [CrossRef]
- MacIntyre, L.W.; Haltli, B.A.; Kerr, R.G. Draft Genome Sequence of Alteromonas sp. Strain RKMC-009, Isolated from Xestospongia muta via In Situ Culturing Using an Isolation Chip Diffusion Chamber. Microbiol. Resour. Announc. 2019, 8, e00508-19. [Google Scholar] [CrossRef]
- Peixoto, R.S.; Sweet, M.; Bourne, D.G. Customized Medicine for Corals. Front. Mar. Sci. 2019, 6, 686. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vilela, C.L.S.; Villela, H.D.M.; Rachid, C.T.C.d.C.; Carmo, F.L.d.; Vermelho, A.B.; Peixoto, R.S. Exploring the Diversity and Biotechnological Potential of Cultured and Uncultured Coral-Associated Bacteria. Microorganisms 2021, 9, 2235. https://doi.org/10.3390/microorganisms9112235
Vilela CLS, Villela HDM, Rachid CTCdC, Carmo FLd, Vermelho AB, Peixoto RS. Exploring the Diversity and Biotechnological Potential of Cultured and Uncultured Coral-Associated Bacteria. Microorganisms. 2021; 9(11):2235. https://doi.org/10.3390/microorganisms9112235
Chicago/Turabian StyleVilela, Caren Leite Spindola, Helena Dias Müller Villela, Caio Tavora Coelho da Costa Rachid, Flávia Lima do Carmo, Alane Beatriz Vermelho, and Raquel Silva Peixoto. 2021. "Exploring the Diversity and Biotechnological Potential of Cultured and Uncultured Coral-Associated Bacteria" Microorganisms 9, no. 11: 2235. https://doi.org/10.3390/microorganisms9112235
APA StyleVilela, C. L. S., Villela, H. D. M., Rachid, C. T. C. d. C., Carmo, F. L. d., Vermelho, A. B., & Peixoto, R. S. (2021). Exploring the Diversity and Biotechnological Potential of Cultured and Uncultured Coral-Associated Bacteria. Microorganisms, 9(11), 2235. https://doi.org/10.3390/microorganisms9112235