
Qualitative and Quantitative Characteristics of Soil Microbiome of Barents Sea Coast, Kola Peninsula



Abstract
:1. Introduction
2. Objects and Methods
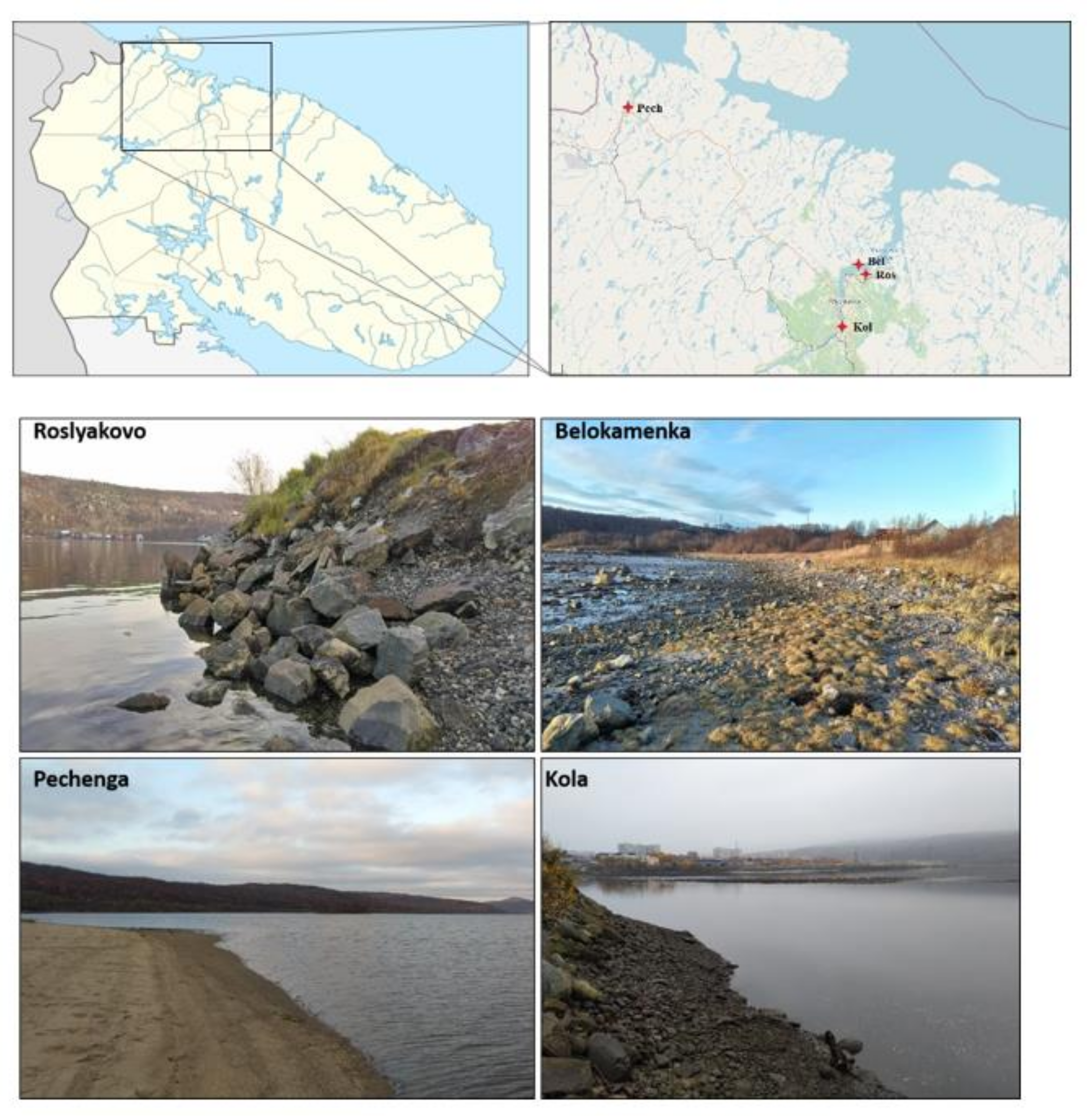
2.1. Research Area
2.2. Site Description
2.3. Sampling Procedure
2.4. Biomass of Prokaryotes
2.5. Fungal Biomass
2.6. Statistical Analysis
3. Results
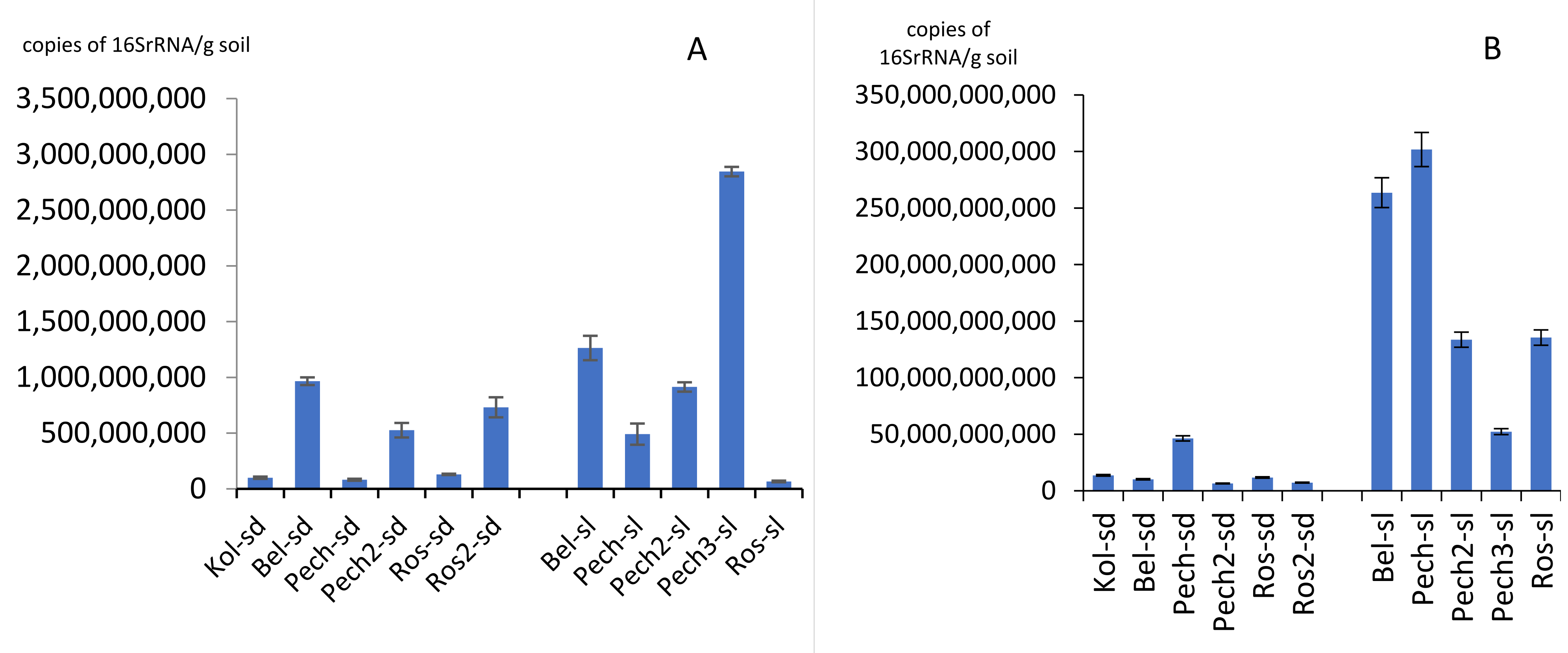
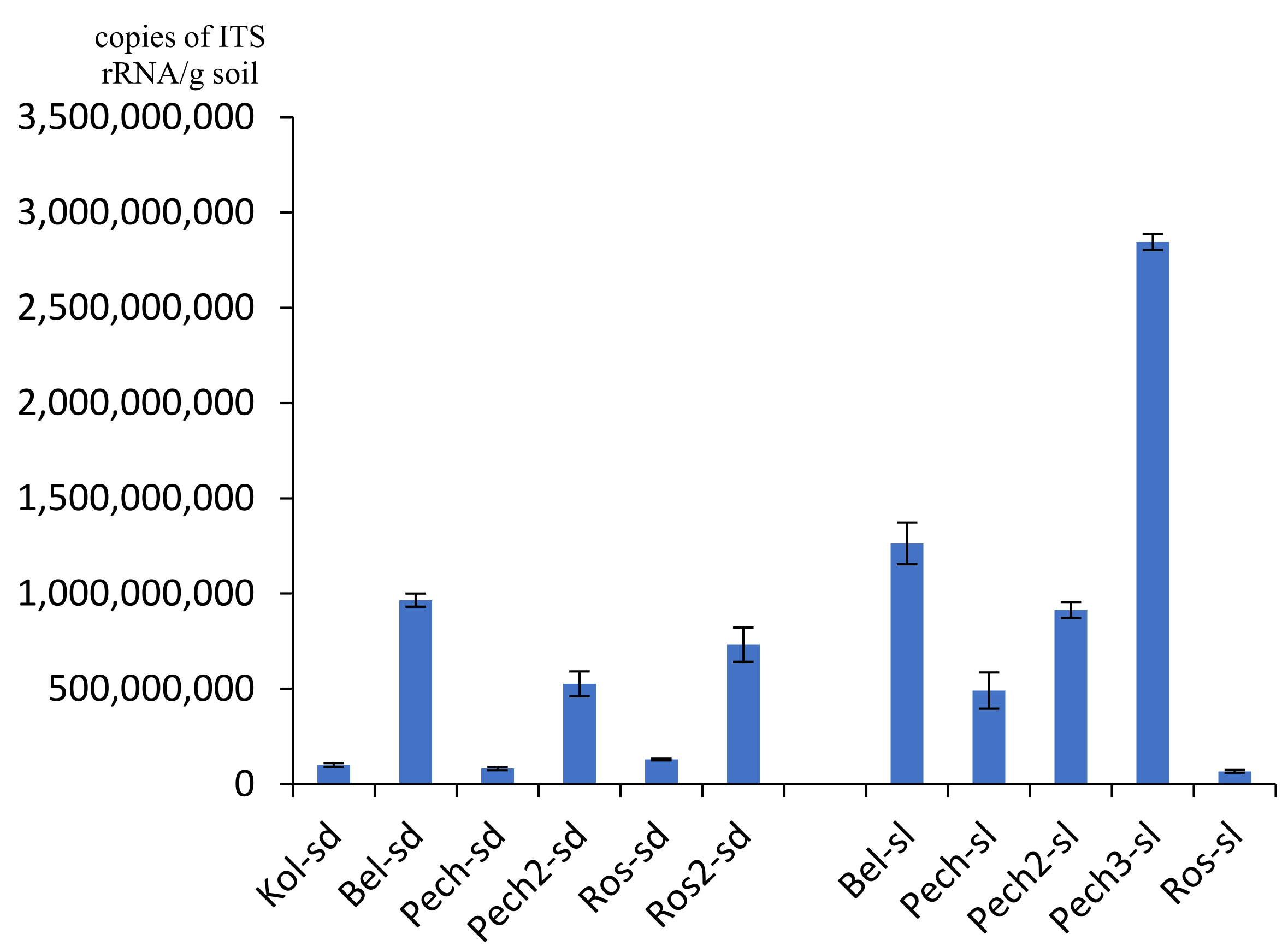
3.1. Number of rRNA Gene Copies
3.2. Microbial Biomass
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Disclaimer
References
- Rühland, K.M.; Paterson, A.M.; Keller, W.; Michelutti, N.; Smol, J.P. Global warming triggers the loss of a key Arctic refugium. Proc. R. Soc. B Biol. Sci. 2013, 280, 20131887. [Google Scholar] [CrossRef]
- Pereverzev, V.N. Soils and soil cover of the Kola Peninsula: History and current state of research. Bull. Kola Sci. Cent. Russ. Acad. Sci. 2011, 1, 39–43. [Google Scholar]
- Evdokimova, G.A.; Mozgova, N.P. Comparative characteristics of microbial biomass of AI-FE-humus podzols of the Kola Peninsula. Eurasian Soil Sci. 2001, 12, 1465–1472. [Google Scholar]
- Pereverzev, V.N. Forest soils of the Kola Peninsula. For. Green Build. West. Sib. 2007, 225–231. Materials of the third international webinar, (In Russian) [Google Scholar]
- Korneykova, M.V. Comparative analysis of the number and structure of the complexes of microscopic fungi in tundra and taiga soils in the north of the Kola Peninsula. Eurasian Soil Sci. 2018, 51, 89–95. [Google Scholar] [CrossRef]
- Nikitin, D.A.; Semenov, M.V.; Chernov, T.I.; Ksenofontova, N.A.; Zhelezova, A.D.; Ivanova, E.A.; Kozlov, D.N.; Khitrov, N.B.; Stepanov, A.L. Microbiological indicators of ecological functions of soils (review). Eurasian Soil Sci. 2021. (in print). [Google Scholar]
- Fierer, N.; Wood, S.A.; de Mesquita, C.P. How microbes can, and cannot, be used to assess soil health. Soil Biol. Biochem. 2021, 153, 108111. [Google Scholar] [CrossRef]
- Marshall, G.J.; Vignols, R.M.; Rees, W.G. Climate change in the Kola Peninsula, Arctic Russia, during the last 50 years from meteorological observations. J. Clim. 2016, 29, 6823–6840. [Google Scholar] [CrossRef]
- Hättestrand, C.; Clark, C.D. The glacial geomorphology of Kola Peninsula and adjacent areas in the Murmansk Region, Russia. J. Maps 2006, 2, 30–42. [Google Scholar] [CrossRef]
- Jacoby, R.; Peukert, M.; Succurro, A.; Koprivova, A.; Kopriva, S. The role of soil microorganisms in plant mineral nutrition—current knowledge and future directions. Front. Plant Sci. 2017, 8, 1617. [Google Scholar] [CrossRef] [Green Version]
- Koroleva, N.E.; Chinenko, S.V.; Sortland, E.B. Communities of marches, beaches and coastal floodplain ephemeretum of the Murmansk, Tersk and Vostok of the Kandalaksha coast (Murmansk region). Phyto-Divers. East. Eur. 2011, 9, 26–62. [Google Scholar]
- Gorin, S.L.; Leman, V.N. Hydrological regime and pollution in the basin and in the water area of the Pechenga Bay (Varanger Fjord of the Barents Sea) according to long-term observations of the hydrometeorological service. Tr. VNIRO 2017, 165, 66–82. [Google Scholar]
- Polyanskaya, L.M.; Zvyagintsev, D.G. The content and composition of microbial biomass as an index of the ecological status of soil. Eurasian Soil Sci. 2005, 38, 625–633. [Google Scholar]
- Zvyagintsev, D.G. Methods of Soil Microbiology and Biochemistry; Moscow State University Publishing House: Moscow, Russia, 1991; Volume 60. [Google Scholar]
- Semenov, M.V.; Krasnov, G.S.; Semenov, V.M.; Ksenofontova, N.; Zinyakova, N.B.; van Bruggen, A.H. Does fresh farmyard manure introduce surviving microbes into soil or activate soil-borne microbiota? J. Environ. Manag. 2021, 294, 113018. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Goodfellow, M. Nucleic Acid Techniques in Bacterial Systematics; Wiley: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Yu, Y.; Lee, C.; Kim, J.; Hwang, S. Group-specific primer and probe sets to detect methanogenic communities using quantitative real-time polymerase chain reaction. Biotechnol. Bioeng. 2005, 89, 670–679. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, J.A.; Vilgalys, R.; Jacksson, R.B. Assessment of soil microbial community structure by use of taxon-specific quantitative PCR assays. Appl. Environ. Microbiol. 2005, 71, 4117–4120. [Google Scholar] [CrossRef] [Green Version]
- Korneikova, M.V.; Nikitin, D.A. Qualitative and quantitative characteristics of the soil microbiome in the zone of influence of emissions from the Kandalaksha aluminum plant. Eurasian Soil Sci. 2021, 6, 725–734. [Google Scholar] [CrossRef]
- McLean, M.A.; Angilletta, M.J., Jr.; Williams, K.S. If you can’t stand the heat, stay out of the city: Thermal reaction norms of chitinolytic fungi in an urban heat island. J. Therm. Biol. 2005, 30, 384–391. [Google Scholar] [CrossRef]
- Korneikova, M.V.; Nikitin, D.A.; Dolgikh, A.V.; Soshina, A.S. Soil mycobiota of the city of Apatity (Murmansk region). Mikol. Fitopatol. 2020, 54, 264–277. [Google Scholar] [CrossRef]
- Vodyanitsky, Y.N. Organic matter in urban soils (literature review). Eurasian Soil Sci. 2015, 8, 921. [Google Scholar] [CrossRef]
- Vasenev, V.; Kuzyakov, Y. Urban soils as hot spots of anthropogenic carbon accumulation: Review of stocks, mechanisms and driving factors. Land Degrad. Dev. 2018, 29, 1607–1622. [Google Scholar] [CrossRef]
- Lysak, L.V.; Lapygina, E.V. The diversity of bacterial communities in urban soils. Eurasian Soil Sci. 2018, 51, 1050–1056. [Google Scholar] [CrossRef]
- Hui, N.; Jumpponen, A.; Francini, G.; Kotze, D.J.; Liu, X.; Romantschuk, M.; Strömmer, R.; Setälä, H. Soil microbial communities are shaped by vegetation type and park age in cities under cold climate. Environ. Microbiol. 2017, 19, 1281–1295. [Google Scholar] [CrossRef]
- Shoemaker, W.R.; Lennon, J.T. Evolution with a seed bank: The population genetic consequences of microbial dormancy. Evol. Appl. 2018, 11, 60–75. [Google Scholar] [CrossRef] [PubMed]
- Zhelezova, A.; Chernov, T.; Tkhakakhova, A.; Xenofontova, N.; Semenov, M.; Kutovaya, O. Prokaryotic community shifts during soil formation on sands in the tundra zone. PLoS ONE 2019, 14, e0206777. [Google Scholar]
- Demyanenko, D.A. Analysis of the state of the environment in the Murmansk region. In Proceedings of the Current Problems and Prospects for the Development of State Statistics in Modern Conditions, Saratov, Russia, 20–22 January 2020. [Google Scholar]
- Evdokimova, G.A.; Mozgova, N.P.; Korneikova, M.V. Content and toxicity of heavy metals in soils of zone affected by gas-air emissions from the Pechenganikel plant. Eurasian Soil Sci. 2014, 5, 531–625. [Google Scholar] [CrossRef]
- Korneikova, M.V.; Redkina, V.V.; Myazin, V.A.; Fokina, N.V.; Shalygina, R.R. Microorganisms of soils of the Rybachiy Peninsula. Proc. Kola Sci. Cent. Russ. Acad. Sci. USA 2019, 10, 108–122. [Google Scholar] [CrossRef]
- Nikitin, D.A.; Lysak, L.V.; Kholod, S.S.; Mergelov, N.S.; Dolgikh, A.V.; Goryachkin, S.V. Biological activity of soils on Northern (Novaya Zemlya). Eurasian Soil Sci. 2021, 10, 1207–1230. [Google Scholar] [CrossRef]
- Fomicheva, O.A.; Polyanskaya, L.M.; Nikonov, V.V.; Lukina, N.V.; Orlova, M.A.; Isaeva, L.G.; Zvyagintsev, D.G. Population and biomass of soil microorganisms in old-growth primary spruce forests in the Northern Taiga. Eurasian Soil Sci. 2006, 39, 1323–1331. [Google Scholar] [CrossRef]
- Garrett, S.D. Soil Fungi and Soil Fertility: An Introduction to Soil Mycology; Elsevier: Exeter, UK, 2016. [Google Scholar]
- Nikitin, D.A.; Marfenina, O.E.; Maksimova, I.A. Using the succession approach in studying the species composition of microscopic fungi and the content of fungal biomass in Antarctic soils. Mikol. Fitopatol. 2017, 51, 211–219. (In Russian) [Google Scholar]
- Rozali, S.N.; Milani, E.A.; Deed, R.C.; Silva, F.V. Bacteria, mold and yeast spore inactivation studies by scanning electron microscope observations. Int. J. food Microbiol. 2017, 263, 17–25. [Google Scholar] [CrossRef]
- Nikitin, D.A.; Lysak, L.V.; Mergelov, N.S.; Dolgikh, A.V.; Zazovskaya, E.P.; Goryachkin, S.V. Microbial Biomass, Carbon Stocks, and CO2 Emission in Soils of Franz Josef Land: High-Arctic Tundra or Polar Deserts? Eurasian Soil Sci. 2020, 53, 467–484. [Google Scholar] [CrossRef]
- Popova, O.N.; Shcherbina, Y.F. Climatogeophysical characteristics of the Kola Polar region. Ekol. Cheloveka 2012, 5, 3–7. (In Russian) [Google Scholar]
- Schmidt, N.; Bölter, M. Fungal and bacterial biomass in tundra soils along an arctic transect from Taimyr Peninsula, central Siberia. Polar Biol. 2002, 25, 871–877. [Google Scholar] [CrossRef]
- Nikitin, D.A.; Semenov, M.V.; Semikolennykh, A.A.; Kachalkin, A.V.; Ivanova, A.E. Biomass of fungi and species diversity of the cultivated mycobiota of soils and substrates in Northbrook Island (Franz Josef Land). Mikol. Fitopatol. 2019, 53, 210–222. [Google Scholar] [CrossRef]
- Khabibullina, F.M.; Kuznetsova, E.G.; Vaseneva, I.Z. Micromycetes in podzolic and bog-podzolic soils in the middle taiga subzone of northeastern European Russia. Eurasian Soil Sci. 2014, 47, 1027–1032. [Google Scholar] [CrossRef]
- Nikitin, D.A.; Chernov, T.V.; Zhelezova, A.D.; Tkhakakhova, A.K.; Nikitina, S.A.; Semenov, M.V.; Xenofontova, N.A.; Kutovaya, O.V. Seasonal Dynamics of Microbial Biomass in Soddy-Podzolic Soil. Eurasian Soil Sci. 2019, 52, 1414–1421. [Google Scholar] [CrossRef]
- Polyanskaya, L.M.; Sukhanova, N.I.; Chakmazyan, K.V.; Zvyagintsev, D.G. Changes in the structure of soil microbial biomass under fallow. Eurasian Soil Sci. 2012, 45, 710–716. [Google Scholar] [CrossRef]
- Bakermans, C.; Emili, L.A. Terrestrial systems of the Arctic as a model for growth and survival at low temperatures. In Model Ecosystems in Extreme Environments; Academic Press: Cambridge, MA, USA, 2019; pp. 1–21. [Google Scholar] [CrossRef]
- Pastor, A.; Freixa, A.; Skovsholt, L.J.; Wu, N.; Romaní, A.M.; Riis, T. Microbial Organic Matter Utilization in High-Arctic Streams: Key Enzymatic Controls. Microb. Ecol. 2019, 78, 539–554. [Google Scholar] [CrossRef] [PubMed]
- Dubrova, M.S.; Lubsanova, D.A.; Makarova, E.P.; Kozhevin, P.A.; Manucharova, N.A.; Zenova, G.M. Psychrotolerant actinomycetes in soils of the tundra and northern taiga. Mosc. Univ. Soil Sci. Bull. 2011, 66, 45–49. [Google Scholar] [CrossRef]
- Millán-Aguiñaga, N.; Soldatou, S.; Brozio, S.; Munnoch, J.T.; Howe, J.; Hoskisson, P.A.; Duncan, K.R. Awakening ancient polar Actinobacteria: Diversity, evolution and specialized metabolite potential. Microbiology 2019, 165, 1169–1180. [Google Scholar] [CrossRef] [PubMed]
- Kudinova, A.G.; Lysak, L.V.; Soina, V.S.; Mergelov, N.S.; Dolgikh, A.V.; Shorkunov, I.G. Bacterial communities in the soils of cryptogamic barrens of East Antarctica (the Larsemann Hills and Thala Hills oases). Eurasian Soil Sci. 2015, 48, 276–287. [Google Scholar] [CrossRef]
- Ivashchenko, K.; Ananyeva, N.; Vasenev, V.; Sushko, S.; Seleznyova, A.; Kudeyarov, V. Microbial C-availability and organic matter decomposition in urban soils of megapolis depend on functional zoning. Soil Environ. 2019, 38, 31–41. [Google Scholar] [CrossRef]
- Grosse, G.; Harden, J.; Turetsky, M.; McGuire, A.D.; Camill, P.; Tarnocai, C.; Frolking, S.; Schuur, E.A.G.; Jorgenson, T.; Marchenko, S.; et al. Vulnerability of high-latitude soil organic carbon in North America to disturbance. J. Geophys Res. Biogeosci. 2011, 116, G00K06. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, K.; Lal, R. Managing soil carbon stocks to enhance the resilience of urban ecosystems. Carbon Manag. 2015, 6, 35–50. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality | Coordinates | Sample Code | TOC,% | pH | Petroleum Hydrocarbon Content, mg/kg | Description of the Site |
|---|---|---|---|---|---|---|
| Kola | 68°53′07.3″ N 33°02′22.6″ E | Kol-sd | 0.81 | 6.81 | 711 | The foot of the coastal ledge; rocky beach with deposits of sand between stones |
| Belokamenka | 69°04′35.7″ N 33°10′12.8″ E | Bel-sl | 13.25 | 5.57 | 568 | Soil: 0–7 cm—organogenic, deeper than 7 cm—sand; Leymus arenarius, Lathyrus aleuticus, Mertensia maritima, Festuca arenaria, Ligusticum scoticum, Achillea apiculata, Atriplex glabriuscula, Chamaenerion angustifolium |
| Bel-sd | 0.24 | 6.43 | 211 | Sandy and gravelly beach with a lot of stones | ||
| Pechenga | 69°34′38.6″ N 31°13′58.5″ E | Pech-sl | 22.26 | 5.94 | 130 | Soil: 0–3 cm—organogenic, deeper than 3 cm—sand; Rumex confertus, Caltha palustris, Chamaenerion angustifolium, Festuca arenaria |
| Pech-sd | 1.25 | 5.42 | 59 | Sandy and gravelly beach | ||
| 69°34′09.0″ N 31°13′50.4″ E | Pech2-sl | 4.74 | 5.11 | 97 | Soil: 0–2 cm—organogenic, deeper than 2 cm—medium loam (hydrogen sulfide decay is in the loam); Carex sp. | |
| Pech2-sd | 0.10 | 7.19 | 46 | Sandy beach | ||
| Pech3-sl | 9.74 | 5.70 | 174 | Soil: 0–5 cm—organogenic, deeper than 5 cm—sand; Solidago lapponica, Chamaenerion angustifolium, Salix sp., Festuca arenaria | ||
| Roslyakovo | 69°03′22.4″ N 33°13′39.3″ E | Ros-sl | 17.44 | 6.86 | 203 | Soil: 0–5 cm—organogenic, deeper than 5 cm—sand and gravel; Achillea apiculate, Chamaenerion angustifolium, Salix sp., Taraxacum officinale, Festuca sp. |
| Ros-sd | 1.32 | 7.22 | 252 | The foot of the coastal ledge; rocky beach with deposits of sand and gravel between stones | ||
| Ros2-sd | 1.25 | 7.20 | 245 | The foot of the coastal ledge; rocky beach with deposits of gravel between stones |
| Sample Description | Mycelium (Mainly d = 3 μm) | Spores (Diameter, μm) | Total Spore Biomass, mg/g Soil | The Portion of Small (2–3 μm) Spores by Weight,% | Total Biomass of Fungi, mg/g Soil | Average Biomass of Fungi by Location, mg/g Soil | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Biomass, mg/g | Length, m | Proportion of Mycelium in Total Biomass,% | 2 | 3 | 5 | |||||||||
| Number, Cells/g × 104 | Biomass, mg/g | Number, Cells/g × 104 | Biomass, mg/g | Number, Cells /g × 103 | Biomass, mg/g | |||||||||
| Kola | Kol-sd | 0.027 ± 0.004 | 21.59 ± 2.96 | 54.0 | 4.32 ± 0.59 | 0.015 ± 0.002 | 0.65 ± 0.09 | 0.008 ± 0.001 | - | - | 0.023 ± 0.004 | 100 | 0.050 ± 0.009 | 0.050 ± 0.009 |
| Belokamenka | Bel-sl | 0.024 ± 0.003 | 190.21 ± 26.08 | 32.9 | 3.88 ± 0.53 | 0.013 ± 0.002 | 1.62 ± 0.22 | 0.019 ± 0.003 | 1.74 ± 0.34 | 0.017 ± 0.002 | 0.049 ± 0.007 | 65.3 | 0.073 ± 0.011 | 0.058 ± 0.010 |
| Bel-sd | 0.015 ± 0.002 | 11.54 ± 1.58 | 34.9 | 2.59 ± 0.35 | 0.009 ± 0.001 | 1.62 ± 0.22 | 0.019 ± 0.003 | - | - | 0.028 ± 0.004 | 100 | 0.043 ± 0.006 | ||
| Pechenga | Pech-sl | 0.303 ± 0.045 | 240.02 ± 32.91 | 86.3 | 6.04 ± 0.83 | 0.020 ± 0.003 | 2.43 ± 0.34 | 0.028 ± 0.004 | - | - | 0.048 ± 0.007 | 100 | 0.351 ± 0.053 | 0.342 ± 0.051 |
| Pech2-sl | 0.637 ± 0.094 | 504.22 ± 69.13 | 89.1 | 9.06 ± 1.25 | 0.031 ± 0.004 | 3.24 ± 0.45 | 0.037 ± 0.005 | 1.04 ± 0.20 | 0.010 ± 0.001 | 0.078 ± 0.012 | 87.2 | 0.715 ± 0.107 | ||
| Pech-sd | 0.018 ± 0.002 | 14.51 ± 1.99 | 43.9 | 3.45 ± 0.47 | 0.012 ± 0.002 | 0.97 ± 0.13 | 0.011 ± 0.001 | - | - | 0.023 ± 0.004 | 100 | 0.041 ± 0.006 | ||
| Pech2-sd | 0.008 ± 0.001 | 6.33 ± 0.87 | 25.0 | 2.59 ± 0.36 | 0.009 ± 0.001 | 1.30 ± 0.17 | 0.015 ± 0.002 | - | - | 0.024 ± 0.004 | 100 | 0.032 ± 0.004 | ||
| Pech3-sl | 0.488 ± 0.072 | 386.55 ± 53.00 | 85.6 | 4.74 ± 0.65 | 0.016 ± 0.002 | 4.86 ± 0.65 | 0.056 ± 0.008 | 1.04 ± 0.20 | 0.010 ± 0.001 | 0.082 ± 0.012 | 87.8 | 0.570 ± 0.086 | ||
| Roslyakovo | Ros-sl | - | - | 0.0 | 3.02 ± 0.42 | 0.010 ± 0.001 | 0.97 ± 0.13 | 0.011 ± 0.001 | - | - | 0.021 ± 0.003 | 100 | 0.021 ± 0.003 | 0.120 ± 0.018 |
| Ros-sd | 0.250 ± 0.037 | 195.30 ± 26.78 | 85.6 | 4.74 ± 0.65 | 0.016 ± 0.002 | 2.27 ± 0.30 | 0.026 ± 0.004 | - | - | 0.042 ± 0.006 | 100 | 0.292 ± 0.044 | ||
| Ros2-sd | - | - | 0.0 | 3.02 ± 0.42 | 0.010 ± 0.001 | 1.94 ± 0.26 | 0.023 ± 0.003 | 1.39 ± 0.27 | 0.013 ± 0.002 | 0.046 ± 0.007 | 71.7 | 0.046 ± 0.007 | ||
| Description Sample | Number of Prokaryotic Cells, × 108 Cells/g | Biomass of Unicellular Prokaryotes, μg/g Soil | Actinomycete Mycelium Length, m/g | Biomass of Actinomycete Mycelium, μg/g | The Share of Mycelium in the Total Biomass, % | Total Biomass of Prokaryotes, μg/g Soil | Average Biomass of Prokaryotes by Location, μg/g Soil | |
|---|---|---|---|---|---|---|---|---|
| Kola | Kol-sd | 2.96 ± 0.43 | 0.63 ± 0.10 | - | - | 0.0 | 0.63 ± 0.10 | 0.63 ± 0.10 |
| Belokamenka | Bel-sl | 10.00 ± 1.47 | 2.12 ± 0.32 | 8.22 ± 1.24 | 0.30 ± 0.05 | 12.4 | 2.42 ± 0.38 | 1.53 ± 0.24 |
| Bel-sd | 2.48 ± 0.36 | 0.53 ± 0.08 | 2.84 ± 0.43 | 0.10 ± 0.01 | 15.9 | 0.63 ± 0.10 | ||
| Pechenga | Pech-sl | 13.99 ± 2.05 | 2.97 ± 0.44 | 25.24 ± 3.82 | 0.92 ± 0.16 | 23.8 | 3.90 ± 0.62 | 2.40 ± 0.38 |
| Pech2-sl | 11.49 ± 1.68 | 2.43 ± 0.36 | 88.18 ± 13.33 | 3.22 ± 0.54 | 57.3 | 5.69 ± 0.90 | ||
| Pech-sd | 1.87 ± 0.27 | 0.40 ± 0.06 | 22.12 ± 3.34 | 0.81 ± 0.14 | 66.4 | 1.22 ± 0.19 | ||
| Pech2-sd | 3.80 ± 0.56 | 0.81 ± 0.12 | 4.17 ± 0.63 | 0.15 ± 0.03 | 15.6 | 0.96 ± 0.15 | ||
| Pech3-sl | 9.99 ± 1.47 | 2.10 ± 0.31 | 0.77 ± 0.12 | 0.03 | 1.4 | 2.13 ± 0.34 | ||
| Roslakovo | Ros-sl | 10.40 ± 1.53 | 2.18 ± 0.32 | 3.04 ± 0.46 | 0.11 ± 0.01 | 4.8 | 2.29 ± 0.36 | 2.09 ± 0.33 |
| Ros-sd | 10.05 ± 1.48 | 2.11 ± 0.31 | 1.97 ± 0.30 | 0.72 ± 0.14 | 25.4 | 2.84 ± 0.45 | ||
| Ros2-sd | 5.27 ± 0.78 | 1.06 ± 0.16 | 2.67 ± 0.40 | 0.09 ± 0.01 | 7.8 | 1.15 ± 0.18 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korneykova, M.; Nikitin, D.; Myazin, V. Qualitative and Quantitative Characteristics of Soil Microbiome of Barents Sea Coast, Kola Peninsula. Microorganisms 2021, 9, 2126. https://doi.org/10.3390/microorganisms9102126
Korneykova M, Nikitin D, Myazin V. Qualitative and Quantitative Characteristics of Soil Microbiome of Barents Sea Coast, Kola Peninsula. Microorganisms. 2021; 9(10):2126. https://doi.org/10.3390/microorganisms9102126
Chicago/Turabian StyleKorneykova, Maria, Dmitry Nikitin, and Vladimir Myazin. 2021. "Qualitative and Quantitative Characteristics of Soil Microbiome of Barents Sea Coast, Kola Peninsula" Microorganisms 9, no. 10: 2126. https://doi.org/10.3390/microorganisms9102126
APA StyleKorneykova, M., Nikitin, D., & Myazin, V. (2021). Qualitative and Quantitative Characteristics of Soil Microbiome of Barents Sea Coast, Kola Peninsula. Microorganisms, 9(10), 2126. https://doi.org/10.3390/microorganisms9102126