
Sex-Specific Associations between Gut Microbiome and Non-Alcoholic Fatty Liver Disease among Urban Chinese Adults

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. NAFLD Assessment
2.3. Stool Sample Collection and 16S rRNA Gene Sequencing
2.4. Statistical Analysis
3. Results
3.1. Characteristics of the Study Subjects
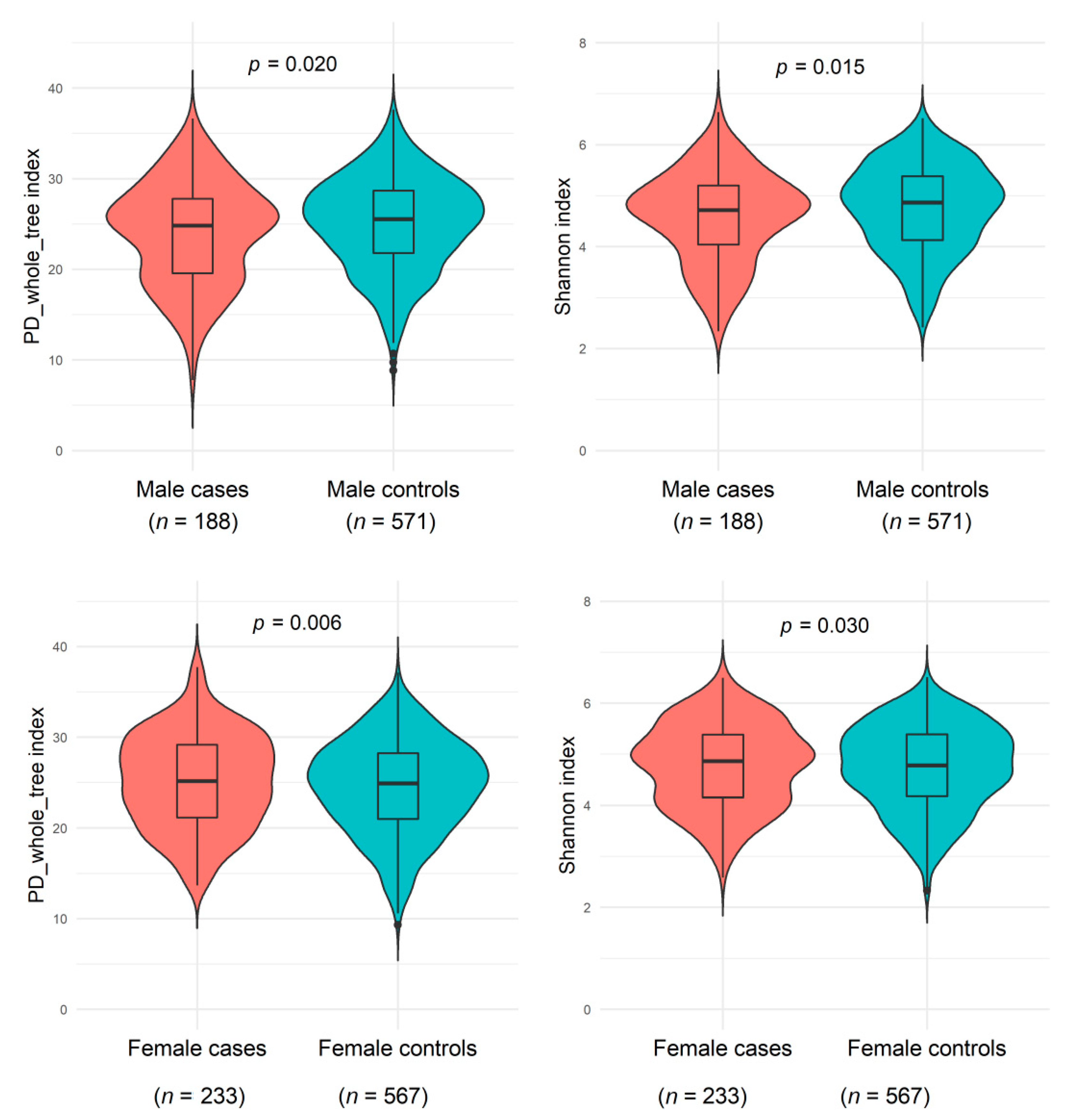
3.2. Associations of NAFLD History with Gut Microbiome Alpha and Beta Diversity
3.3. Associations of NAFLD History with Individual Gut Microbial Taxa
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yki-Järvinen, H. Non-Alcoholic Fatty Liver Disease as a Cause and a Consequence of Metabolic Syndrome. Lancet Diabetes Endocrinol. 2014, 2, 901–910. [Google Scholar] [CrossRef]
- Younossi, Z.M. Non-Alcoholic Fatty Liver Disease—A Global Public Health Perspective. J. Hepatol. 2019, 70, 531–544. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Zhou, J.; Wang, W.; Zhang, X.-J.; Ji, Y.-X.; Zhang, P.; She, Z.-G.; Zhu, L.; Cai, J.; Li, H. Unexpected Rapid Increase in the Burden of NAFLD in China From 2008 to 2018: A Systematic Review and Meta-Analysis. Hepatology 2019, 70, 1119–1133. [Google Scholar] [CrossRef] [PubMed]
- Safari, Z.; Gérard, P. The Links between the Gut Microbiome and Non-Alcoholic Fatty Liver Disease (NAFLD). Cell. Mol. Life Sci. CMLS 2019, 76, 1541–1558. [Google Scholar] [CrossRef]
- Albillos, A.; de Gottardi, A.; Rescigno, M. The Gut-Liver Axis in Liver Disease: Pathophysiological Basis for Therapy. J. Hepatol. 2020, 72, 558–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, F.; Grander, C.; Effenberger, M.; Adolph, T.E.; Tilg, H. Gut Dysfunction and Non-Alcoholic Fatty Liver Disease. Front. Endocrinol. 2019, 10, 611. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.D.; Behary, J.; Zekry, A. Non-Alcoholic Fatty Liver Disease: A Review of Epidemiology, Risk Factors, Diagnosis and Management. Intern. Med. J. 2020, 50, 1038–1047. [Google Scholar] [CrossRef]
- Le Roy, T.; Llopis, M.; Lepage, P.; Bruneau, A.; Rabot, S.; Bevilacqua, C.; Martin, P.; Philippe, C.; Walker, F.; Bado, A.; et al. Intestinal Microbiota Determines Development of Non-Alcoholic Fatty Liver Disease in Mice. Gut 2013, 62, 1787–1794. [Google Scholar] [CrossRef] [PubMed]
- Henao-Mejia, J.; Elinav, E.; Jin, C.; Hao, L.; Mehal, W.Z.; Strowig, T.; Thaiss, C.A.; Kau, A.L.; Eisenbarth, S.C.; Jurczak, M.J.; et al. Inflammasome-Mediated Dysbiosis Regulates Progression of NAFLD and Obesity. Nature 2012, 482, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boursier, J.; Mueller, O.; Barret, M.; Machado, M.; Fizanne, L.; Araujo-Perez, F.; Guy, C.D.; Seed, P.C.; Rawls, J.F.; David, L.A.; et al. The Severity of Nonalcoholic Fatty Liver Disease Is Associated with Gut Dysbiosis and Shift in the Metabolic Function of the Gut Microbiota. Hepatology 2016, 63, 764–775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loomba, R.; Seguritan, V.; Li, W.; Long, T.; Klitgord, N.; Bhatt, A.; Dulai, P.S.; Caussy, C.; Bettencourt, R.; Highlander, S.K.; et al. Gut Microbiome-Based Metagenomic Signature for Non-Invasive Detection of Advanced Fibrosis in Human Nonalcoholic Fatty Liver Disease. Cell Metab. 2017, 25, 1054–1062.e5. [Google Scholar] [CrossRef]
- Caussy, C.; Tripathi, A.; Humphrey, G.; Bassirian, S.; Singh, S.; Faulkner, C.; Bettencourt, R.; Rizo, E.; Richards, L.; Xu, Z.Z.; et al. A Gut Microbiome Signature for Cirrhosis Due to Nonalcoholic Fatty Liver Disease. Nat. Commun. 2019, 10, 1406. [Google Scholar] [CrossRef]
- Li, F.; Ye, J.; Shao, C.; Zhong, B. Compositional Alterations of Gut Microbiota in Nonalcoholic Fatty Liver Disease Patients: A Systematic Review and Meta-Analysis. Lipids Health Dis. 2021, 20, 22. [Google Scholar] [CrossRef]
- Zheng, W.; Chow, W.-H.; Yang, G.; Jin, F.; Rothman, N.; Blair, A.; Li, H.-L.; Wen, W.; Ji, B.-T.; Li, Q.; et al. The Shanghai Women’s Health Study: Rationale, Study Design, and Baseline Characteristics. Am. J. Epidemiol. 2005, 162, 1123–1131. [Google Scholar] [CrossRef] [Green Version]
- Shu, X.-O.; Li, H.; Yang, G.; Gao, J.; Cai, H.; Takata, Y.; Zheng, W.; Xiang, Y.-B. Cohort Profile: The Shanghai Men’s Health Study. Int. J. Epidemiol. 2015, 44, 810–818. [Google Scholar] [CrossRef]
- Yu, D.; Nguyen, S.M.; Yang, Y.; Xu, W.; Cai, H.; Wu, J.; Cai, Q.; Long, J.; Zheng, W.; Shu, X.-O. Long-Term Diet Quality Is Associated with Gut Microbiome Diversity and Composition among Urban Chinese Adults. Am. J. Clin. Nutr. 2021, 113, 684–694. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Cai, Q.; Shu, X.-O.; Steinwandel, M.D.; Blot, W.J.; Zheng, W.; Long, J. Prospective Study of Oral Microbiome and Colorectal Cancer Risk in Low-Income and African American Populations. Int. J. Cancer 2019, 144, 2381–2389. [Google Scholar] [CrossRef] [PubMed]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An Improved Greengenes Taxonomy with Explicit Ranks for Ecological and Evolutionary Analyses of Bacteria and Archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing Mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dixon, P. VEGAN, a Package of R Functions for Community Ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Fernandes, A.D.; Reid, J.N.; Macklaim, J.M.; McMurrough, T.A.; Edgell, D.R.; Gloor, G.B. Unifying the Analysis of High-Throughput Sequencing Datasets: Characterizing RNA-Seq, 16S RRNA Gene Sequencing and Selective Growth Experiments by Compositional Data Analysis. Microbiome 2014, 2, 15. [Google Scholar] [CrossRef] [Green Version]
- Knight, R.; Vrbanac, A.; Taylor, B.C.; Aksenov, A.; Callewaert, C.; Debelius, J.; Gonzalez, A.; Kosciolek, T.; McCall, L.-I.; McDonald, D.; et al. Best Practices for Analysing Microbiomes. Nat. Rev. Microbiol. 2018, 16, 410–422. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.-F. Predictive Values of Body Mass Index and Waist Circumference for Risk Factors of Certain Related Diseases in Chinese Adults--Study on Optimal Cut-off Points of Body Mass Index and Waist Circumference in Chinese Adults. Biomed. Environ. Sci. BES 2002, 15, 83–96. [Google Scholar] [PubMed]
- de la Cuesta-Zuluaga, J.; Kelley, S.T.; Chen, Y.; Escobar, J.S.; Mueller, N.T.; Ley, R.E.; McDonald, D.; Huang, S.; Swafford, A.D.; Knight, R.; et al. Age- and Sex-Dependent Patterns of Gut Microbial Diversity in Human Adults. mSystems 2019, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sinha, T.; Vich Vila, A.; Garmaeva, S.; Jankipersadsing, S.A.; Imhann, F.; Collij, V.; Bonder, M.J.; Jiang, X.; Gurry, T.; Alm, E.J.; et al. Analysis of 1135 Gut Metagenomes Identifies Sex-Specific Resistome Profiles. Gut Microbes 2019, 10, 358–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Unno, T.; Kim, B.Y.; Park, M.S. Sex Differences in Gut Microbiota. World J. Mens Health 2020, 38, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Peterson, V.L.; Richards, J.B.; Meyer, P.J.; Cabrera-Rubio, R.; Tripi, J.A.; King, C.P.; Polesskaya, O.; Baud, A.; Chitre, A.S.; Bastiaanssen, T.F.S.; et al. Sex-Dependent Associations between Addiction-Related Behaviors and the Microbiome in Outbred Rats. EBioMedicine 2020, 55, 102769. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Chen, R.; Zhang, Y.; Lin, X.; Yang, X. Sexual Dimorphism of Gut Microbiota at Different Pubertal Status. Microb. Cell Factories 2020, 19, 152. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P.; et al. Human Gut Microbiome Viewed across Age and Geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef]
- Lakshminarayanan, B.; Stanton, C.; O’Toole, P.W.; Ross, R.P. Compositional Dynamics of the Human Intestinal Microbiota with Aging: Implications for Health. J. Nutr. Health Aging 2014, 18, 773–786. [Google Scholar] [CrossRef]
- Odamaki, T.; Kato, K.; Sugahara, H.; Hashikura, N.; Takahashi, S.; Xiao, J.-Z.; Abe, F.; Osawa, R. Age-Related Changes in Gut Microbiota Composition from Newborn to Centenarian: A Cross-Sectional Study. BMC Microbiol. 2016, 16, 90. [Google Scholar] [CrossRef] [Green Version]
- Raman, M.; Ahmed, I.; Gillevet, P.M.; Probert, C.S.; Ratcliffe, N.M.; Smith, S.; Greenwood, R.; Sikaroodi, M.; Lam, V.; Crotty, P.; et al. Fecal Microbiome and Volatile Organic Compound Metabolome in Obese Humans with Nonalcoholic Fatty Liver Disease. Clin. Gastroenterol. Hepatol. Off. Clin. Pract. J. Am. Gastroenterol. Assoc. 2013, 11, 868–875.e1–3. [Google Scholar] [CrossRef]
- Wong, V.W.-S.; Tse, C.-H.; Lam, T.T.-Y.; Wong, G.L.-H.; Chim, A.M.-L.; Chu, W.C.-W.; Yeung, D.K.-W.; Law, P.T.-W.; Kwan, H.-S.; Yu, J.; et al. Molecular Characterization of the Fecal Microbiota in Patients with Nonalcoholic Steatohepatitis—A Longitudinal Study. PLoS ONE 2013, 8, e62885. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of Gut Microbiomes in Nonalcoholic Steatohepatitis (NASH) Patients: A Connection between Endogenous Alcohol and NASH. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef]
- Jiang, W.; Wu, N.; Wang, X.; Chi, Y.; Zhang, Y.; Qiu, X.; Hu, Y.; Li, J.; Liu, Y. Dysbiosis Gut Microbiota Associated with Inflammation and Impaired Mucosal Immune Function in Intestine of Humans with Non-Alcoholic Fatty Liver Disease. Sci. Rep. 2015, 5, 8096. [Google Scholar] [CrossRef] [PubMed]
- Michail, S.; Lin, M.; Frey, M.R.; Fanter, R.; Paliy, O.; Hilbush, B.; Reo, N.V. Altered Gut Microbial Energy and Metabolism in Children with Non-Alcoholic Fatty Liver Disease. FEMS Microbiol. Ecol. 2015, 91, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Jiang, X.; Cao, M.; Ge, J.; Bao, Q.; Tang, L.; Chen, Y.; Li, L. Altered Fecal Microbiota Correlates with Liver Biochemistry in Nonobese Patients with Non-Alcoholic Fatty Liver Disease. Sci. Rep. 2016, 6, 32002. [Google Scholar] [CrossRef]
- Del Chierico, F.; Nobili, V.; Vernocchi, P.; Russo, A.; De Stefanis, C.; Gnani, D.; Furlanello, C.; Zandonà, A.; Paci, P.; Capuani, G.; et al. Gut Microbiota Profiling of Pediatric Nonalcoholic Fatty Liver Disease and Obese Patients Unveiled by an Integrated Meta-Omics-Based Approach. Hepatology 2017, 65, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Shen, F.; Zheng, R.-D.; Sun, X.-Q.; Ding, W.-J.; Wang, X.-Y.; Fan, J.-G. Gut Microbiota Dysbiosis in Patients with Non-Alcoholic Fatty Liver Disease. Hepatobiliary Pancreat. Dis. Int. 2017, 16, 375–381. [Google Scholar] [CrossRef]
- Da Silva, H.E.; Teterina, A.; Comelli, E.M.; Taibi, A.; Arendt, B.M.; Fischer, S.E.; Lou, W.; Allard, J.P. Nonalcoholic Fatty Liver Disease Is Associated with Dysbiosis Independent of Body Mass Index and Insulin Resistance. Sci. Rep. 2018, 8, 1466. [Google Scholar] [CrossRef] [Green Version]
- Nistal, E.; Sáenz de Miera, L.E.; Ballesteros Pomar, M.; Sánchez-Campos, S.; García-Mediavilla, M.V.; Álvarez-Cuenllas, B.; Linares, P.; Olcoz, J.L.; Arias-Loste, M.T.; García-Lobo, J.M.; et al. An Altered Fecal Microbiota Profile in Patients with Non-Alcoholic Fatty Liver Disease (NAFLD) Associated with Obesity. Rev. Espanola Enfermedades Dig. Organo Of. Soc. Espanola Patol. Dig. 2019, 111, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.; Kim, H.-N.; Lee, E.-J.; Ryu, S.; Chang, Y.; Shin, H.; Kim, H.-L.; Kim, T.H.; Yoo, K.; Kim, H.Y. Fecal and Blood Microbiota Profiles and Presence of Nonalcoholic Fatty Liver Disease in Obese versus Lean Subjects. PLoS ONE 2019, 14, e0213692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, M.-C.; Liu, Y.-Y.; Lin, C.-C.; Wang, C.-C.; Wu, Y.-J.; Yong, C.-C.; Chen, K.-D.; Chuah, S.-K.; Yao, C.-C.; Huang, P.-Y.; et al. Gut Microbiota Dysbiosis in Patients with Biopsy-Proven Nonalcoholic Fatty Liver Disease: A Cross-Sectional Study in Taiwan. Nutrients 2020, 12, 820. [Google Scholar] [CrossRef] [Green Version]
- Al-Jashamy, K.; Murad, A.; Zeehaida, M.; Rohaini, M.; Hasnan, J. Prevalence of Colorectal Cancer Associated with Streptococcus Bovis among Inflammatory Bowel and Chronic Gastrointestinal Tract Disease Patients. Asian Pac. J. Cancer Prev. APJCP 2010, 11, 1765–1768. [Google Scholar] [PubMed]
- Gevers, D.; Kugathasan, S.; Denson, L.A.; Vázquez-Baeza, Y.; Van Treuren, W.; Ren, B.; Schwager, E.; Knights, D.; Song, S.J.; Yassour, M.; et al. The Treatment-Naive Microbiome in New-Onset Crohn’s Disease. Cell Host Microbe 2014, 15, 382–392. [Google Scholar] [CrossRef] [Green Version]
- Forbes, J.D.; Chen, C.-Y.; Knox, N.C.; Marrie, R.-A.; El-Gabalawy, H.; de Kievit, T.; Alfa, M.; Bernstein, C.N.; Van Domselaar, G. A Comparative Study of the Gut Microbiota in Immune-Mediated Inflammatory Diseases-Does a Common Dysbiosis Exist? Microbiome 2018, 6, 221. [Google Scholar] [CrossRef]
- Chen, Y.; Yang, F.; Lu, H.; Wang, B.; Chen, Y.; Lei, D.; Wang, Y.; Zhu, B.; Li, L. Characterization of Fecal Microbial Communities in Patients with Liver Cirrhosis. Hepatology 2011, 54, 562–572. [Google Scholar] [CrossRef]
- Ponziani, F.R.; Bhoori, S.; Castelli, C.; Putignani, L.; Rivoltini, L.; Del Chierico, F.; Sanguinetti, M.; Morelli, D.; Paroni Sterbini, F.; Petito, V.; et al. Hepatocellular Carcinoma Is Associated with Gut Microbiota Profile and Inflammation in Nonalcoholic Fatty Liver Disease. Hepatology 2019, 69, 107–120. [Google Scholar] [CrossRef]
- Behari, J.; Graham, L.; Wang, R.; Schirda, C.; Borhani, A.A.; Methé, B.A.; Li, K.; Morris, A.; Luu, H.N.; Palmieri, S.; et al. Dynamics of Hepatic Steatosis Resolution and Changes in Gut Microbiome with Weight Loss in Nonalcoholic Fatty Liver Disease. Obes. Sci. Pract. 2021, 7, 217–225. [Google Scholar] [CrossRef]
- Santos-Marcos, J.A.; Haro, C.; Vega-Rojas, A.; Alcala-Diaz, J.F.; Molina-Abril, H.; Leon-Acuña, A.; Lopez-Moreno, J.; Landa, B.B.; Tena-Sempere, M.; Perez-Martinez, P.; et al. Sex Differences in the Gut Microbiota as Potential Determinants of Gender Predisposition to Disease. Mol. Nutr. Food Res. 2019, 63, e1800870. [Google Scholar] [CrossRef]
- Manor, O.; Dai, C.L.; Kornilov, S.A.; Smith, B.; Price, N.D.; Lovejoy, J.C.; Gibbons, S.M.; Magis, A.T. Health and Disease Markers Correlate with Gut Microbiome Composition across Thousands of People. Nat. Commun. 2020, 11, 5206. [Google Scholar] [CrossRef] [PubMed]
- Goodrich, J.K.; Waters, J.L.; Poole, A.C.; Sutter, J.L.; Koren, O.; Blekhman, R.; Beaumont, M.; Van Treuren, W.; Knight, R.; Bell, J.T.; et al. Human Genetics Shape the Gut Microbiome. Cell 2014, 159, 789–799. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; Bonder, M.J.; Cenit, M.C.; Tigchelaar, E.F.; Maatman, A.; Dekens, J.A.M.; Brandsma, E.; Marczynska, J.; Imhann, F.; Weersma, R.K.; et al. The Gut Microbiome Contributes to a Substantial Proportion of the Variation in Blood Lipids. Circ. Res. 2015, 117, 817–824. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.A.; Jackson, M.; Jeffery, I.B.; Beaumont, M.; Bell, J.T.; Clark, A.G.; Ley, R.E.; O’Toole, P.W.; Spector, T.D.; Steves, C.J. Signatures of Early Frailty in the Gut Microbiota. Genome Med. 2016, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Org, E.; Blum, Y.; Kasela, S.; Mehrabian, M.; Kuusisto, J.; Kangas, A.J.; Soininen, P.; Wang, Z.; Ala-Korpela, M.; Hazen, S.L.; et al. Relationships between Gut Microbiota, Plasma Metabolites, and Metabolic Syndrome Traits in the METSIM Cohort. Genome Biol. 2017, 18, 70. [Google Scholar] [CrossRef]
- Falony, G.; Joossens, M.; Vieira-Silva, S.; Wang, J.; Darzi, Y.; Faust, K.; Kurilshikov, A.; Bonder, M.J.; Valles-Colomer, M.; Vandeputte, D.; et al. Population-Level Analysis of Gut Microbiome Variation. Science 2016, 352, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.A.; Verdi, S.; Maxan, M.-E.; Shin, C.M.; Zierer, J.; Bowyer, R.C.E.; Martin, T.; Williams, F.M.K.; Menni, C.; Bell, J.T.; et al. Gut Microbiota Associations with Common Diseases and Prescription Medications in a Population-Based Cohort. Nat. Commun. 2018, 9, 2655. [Google Scholar] [CrossRef] [Green Version]
- Vich Vila, A.; Collij, V.; Sanna, S.; Sinha, T.; Imhann, F.; Bourgonje, A.R.; Mujagic, Z.; Jonkers, D.M.A.E.; Masclee, A.A.M.; Fu, J.; et al. Impact of Commonly Used Drugs on the Composition and Metabolic Function of the Gut Microbiota. Nat. Commun. 2020, 11, 362. [Google Scholar] [CrossRef]
- Tap, J.; Cools-Portier, S.; Pavan, S.; Druesne, A.; Öhman, L.; Törnblom, H.; Simren, M.; Derrien, M. Effects of the Long-Term Storage of Human Fecal Microbiota Samples Collected in RNAlater. Sci. Rep. 2019, 9, 601. [Google Scholar] [CrossRef]
- Gavriliuc, S.; Stothart, M.R.; Henry, A.; Poissant, J. Long-Term Storage of Feces at −80 °C versus −20 °C Is Negligible for 16S RRNA Amplicon Profiling of the Equine Bacterial Microbiome. PeerJ 2021, 9, e10837. [Google Scholar] [CrossRef] [PubMed]
- Fianchi, F.; Liguori, A.; Gasbarrini, A.; Grieco, A.; Miele, L. Nonalcoholic Fatty Liver Disease (NAFLD) as Model of Gut-Liver Axis Interaction: From Pathophysiology to Potential Target of Treatment for Personalized Therapy. Int. J. Mol. Sci. 2021, 22, 6485. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Characteristics | Shanghai Men’s Health Study | Shanghai Women’s Health Study | ||||
|---|---|---|---|---|---|---|
| Controls (n = 571) | NAFLD (n = 188) | pa | Controls (n = 567) | NAFLD (n = 233) | pa | |
| Age at stool sample collection (years, mean (SD)) | 67.9 (9.4) | 66.7 (8.2) | 0.189 | 69.9 (8.8) | 69.7 (7.4) | 0.750 |
| Recent body mass index (kg/m2, mean (SD)) | 23.9 (3.2) | 25.8 (3.1) | <0.001 | 23.5 (3.6) | 26.1 (3.6) | <0.001 |
| Recent waist-to-hip ratio (mean (SD)) | 0.89 (0.06) | 0.92 (0.05) | <0.001 | 0.81 (0.05) | 0.83 (0.05) | <0.001 |
| Education (%) | 0.280 | 0.171 | ||||
| Elementary school or less | 5.6 | 4.3 | 17.8 | 12.0 | ||
| Middle school | 29.6 | 36.7 | 41.5 | 41.6 | ||
| High school graduate | 37.7 | 32.4 | 28.0 | 30.5 | ||
| Some college and higher | 27.1 | 26.6 | 12.7 | 15.9 | ||
| High income (%) | 8.8 | 10.6 | 0.096 | 15.9 | 22.3 | 0.020 |
| Smoking | ||||||
| Never smoker (%) | 39.9 | 35.6 | 0.248 | 99.1 | 99.6 | 0.500 |
| Former smoker (%) | 13.0 | 17.6 | 0.9 | 0.4 | ||
| Current smoker (%) | 47.1 | 46.8 | ||||
| Pack-year among smokers (mean (SD)) | 24.2 (14.9) | 23.8 (15.4) | 0.815 | - | - | - |
| Alcohol drinking | ||||||
| Ever alcohol drinking (%) | 34.7 | 29.8 | 0.218 | 4.4 | 7.3 | 0.096 |
| Alcohol consumption (g/day, mean (SD)) b | 2.28 (1.82) | 2.65 (2.32) | 0.188 | 0.13 (0.54) | 0.07 (0.13) | 0.110 |
| Stool sample collection season (%) | 0.602 | 0.273 | ||||
| Spring | 10 | 13.3 | 10.4 | 15 | ||
| Summer | 6.8 | 6.9 | 7.4 | 8.2 | ||
| Autumn | 36.4 | 36.7 | 41.1 | 39.9 | ||
| Winter | 46.8 | 43.1 | 41.1 | 36.9 | ||
| Leisure-time physical activity (yes, %) | 55.7 | 53.2 | 0.550 | 61.9 | 56.2 | 0.136 |
| Healthy diet score (mean (SD)) | 25.1 (5.1) | 25.2 (5.5) | 0.655 | 24.1 (5.0) | 23.9 (5.0) | 0.651 |
| Total energy intake (kcal/day, mean (SD)) | 1968 (488) | 1954 (460) | 0.994 | 1508 (307) | 1550 (338) | 0.079 |
| Fiber intake (g/day, mean (SD)) | 12.3 (3.6) | 12.1 (3.5) | 0.508 | 10.0 (2.7) | 10.1 (2.4) | 0.362 |
| Fat intake (g/day, mean (SD)) | 33.9 (10.0) | 34.3 (10.4) | 0.986 | 28.5 (7.1) | 27.3 (7.1) | 0.006 |
| History of hypertension (yes, %) | 13.8 | 18.1 | 0.156 | 18.7 | 26.6 | 0.013 |
| History of dyslipidemia (yes, %) | 6.0 | 19.2 | <0.001 | 14.6 | 44.6 | <0.001 |
| Bowel movement (times/week, mean (SD)) | 7.7 (3.6) | 7.8 (3.6) | 0.926 | 7.3 (3.7) | 7.7 (4.0) | 0.071 |
| SMHS | SWHS | p for Interaction with Sex d | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Non-NAFLD (n = 571) | NAFLD (n = 188) | Non-NAFLD (n = 567) | NAFLD (n = 233) | ||||||
| Taxon b | Median RA (%) | Median RA (%) | Beta (se) c | pc | Median RA (%) | Median RA (%) | Beta (se) c | pc | |
| p_Actinobacteria;c_Actinobacteria;o_Bifidobacteriales;f_Bifidobacteriaceae;g_Bifidobacterium;Other | 0.0075 | 0.0106 | 0.369 (0.171) | 0.031 | 0.0089 | 0.0068 | −0.439 (0.163) | 0.007 | 0.002 |
| p_Actinobacteria;c_Actinobacteria;o_Bifidobacteriales;f_Bifidobacteriaceae;g_Bifidobacterium;s_adolescentis | 0.0441 | 0.0585 | 0.474 (0.218) | 0.03 | 0.0635 | 0.0506 | −0.399 (0.207) | 0.055 | 0.008 |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;f_[Odoribacteraceae];g_Butyricimonas | 0.0906 | 0.032 | −0.238 (0.221) | 0.28 | 0.0463 | 0.1061 | 0.632 (0.214) | 0.003 | 0.007 |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;f_[Odoribacteraceae];g_Butyricimonas;s_unclassified | 0.0906 | 0.032 | −0.23 (0.217) | 0.291 | 0.0463 | 0.1061 | 0.639 (0.212) | 0.003 | 0.005 |
| p_Firmicutes;c_Bacilli;o_Lactobacillales;f_Streptococcaceae;g_Streptococcus | 0.0787 | 0.1144 | 0.363 (0.148) | 0.015 | 0.1027 | 0.0804 | −0.104 (0.139) | 0.454 | 0.012 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_unclassified | 0.0113 | 0.0043 | −0.277 (0.231) | 0.231 | 0.0052 | 0.0127 | 0.674 (0.225) | 0.003 | 0.002 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_unclassified;g_unclassified | 0.0113 | 0.0043 | −0.239 (0.228) | 0.294 | 0.0052 | 0.0127 | 0.66 (0.222) | 0.003 | 0.002 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_unclassified;g_unclassified;s_unclassified | 0.0113 | 0.0043 | −0.23 (0.228) | 0.313 | 0.0052 | 0.0127 | 0.667 (0.222) | 0.003 | 0.002 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Ruminococcaceae;g_Oscillospira;Other | 0.0425 | 0.0093 | −0.307 (0.195) | 0.115 | 0.0146 | 0.0275 | 0.477 (0.183) | 0.009 | 0.005 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Veillonellaceae;g_Dialister | 0.0214 | 0.0554 | 0.788 (0.241) | 0.001 | 0.0467 | 0.0305 | −0.487 (0.241) | 0.044 | 2.4 × 10−4 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Veillonellaceae;g_Dialister;s_unclassified | 0.0214 | 0.0554 | 0.796 (0.243) | 0.001 | 0.0467 | 0.0305 | −0.481 (0.245) | 0.05 | 3.3 × 10−4 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Veillonellaceae;g_Phascolarctobacterium | 1.672 | 0.9446 | −0.555 (0.217) | 0.011 | 1.4603 | 1.6876 | 0.376 (0.238) | 0.116 | 0.007 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Veillonellaceae;g_Phascolarctobacterium;s_unclassified | 1.672 | 0.9446 | −0.546 (0.216) | 0.012 | 1.4603 | 1.6876 | 0.382 (0.238) | 0.108 | 0.006 |
| SMHS | SWHS | p for Interaction with Sex d | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Non-NAFLD (n = 571) | NAFLD (n = 188) | Non-NAFLD (n = 567) | NAFLD (n = 233) | ||||||
| Taxon b | Carrier Frequency (%) | Carrier Frequency (%) | Beta (se) c | pc | Carrier Frequency (%) | Carrier Frequency (%) | Beta (se) c | pc | |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;f_[Paraprevotellaceae] | 50.8 | 42 | −0.314 (0.189) | 0.097 | 39.3 | 47.6 | 0.384 (0.182) | 0.035 | 0.008 |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;f_Rikenellaceae;f\_\_Rikenellaceae_unclassified | 50.3 | 41 | −0.421 (0.187) | 0.024 | 43.7 | 49.8 | 0.400 (0.182) | 0.028 | 0.005 |
| p_Bacteroidetes;c_Bacteroidia;o_Bacteroidales;f_Rikenellaceae;f\_\_Rikenellaceae_unclassified;Other | 50.3 | 41 | −0.421 (0.187) | 0.024 | 43.7 | 49.8 | 0.400 (0.182) | 0.028 | 0.005 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_[Mogibacteriaceae] | 51.3 | 37.8 | −0.551 (0.191) | 0.004 | 48.7 | 49.8 | 0.285 (0.186) | 0.125 | 0.002 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_[Mogibacteriaceae];g_unclassified | 45.9 | 32.4 | −0.551 (0.197) | 0.005 | 42.7 | 44.2 | 0.344 (0.186) | 0.064 | 0.001 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_[Mogibacteriaceae];g_unclassified;s_unclassified | 45.9 | 32.4 | −0.551 (0.197) | 0.005 | 42.7 | 44.2 | 0.344 (0.186) | 0.064 | 0.001 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Christensenellaceae | 35.2 | 26.6 | −0.312 (0.207) | 0.132 | 34.4 | 36.9 | 0.422 (0.193) | 0.029 | 0.007 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Christensenellaceae;g_unclassified | 32.7 | 25.5 | −0.300 (0.210) | 0.154 | 31.9 | 36.1 | 0.516 (0.196) | 0.008 | 0.005 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Christensenellaceae;g_unclassified;s_unclassified | 32.7 | 25.5 | −0.300 (0.210) | 0.154 | 31.9 | 36.1 | 0.516 (0.196) | 0.008 | 0.005 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Peptococcaceae | 24.9 | 16 | −0.481 (0.240) | 0.045 | 24 | 25.3 | 0.256 (0.212) | 0.227 | 0.011 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Peptococcaceae;g_unclassified | 20.3 | 10.6 | −0.572 (0.278) | 0.040 | 19.6 | 21.5 | 0.352 (0.226) | 0.120 | 0.002 |
| p_Firmicutes;c_Clostridia;o_Clostridiales;f_Peptococcaceae;g_unclassified;s_unclassified | 20.3 | 10.6 | −0.572 (0.278) | 0.040 | 19.6 | 21.5 | 0.352 (0.226) | 0.120 | 0.002 |
| p_Tenericutes | 28.2 | 20.2 | −0.323 (0.224) | 0.149 | 24 | 27.5 | 0.327 (0.207) | 0.115 | 0.004 |
| p_Tenericutes;c_Mollicutes | 26.6 | 17 | −0.517 (0.234) | 0.027 | 21.5 | 27 | 0.417 (0.211) | 0.049 | 3.6 × 10−4 |
| p_Tenericutes;c_Mollicutes;o_RF39 | 25.9 | 16.5 | −0.516 (0.237) | 0.029 | 21.5 | 27 | 0.417 (0.211) | 0.049 | 3.8 × 10−4 |
| p_Tenericutes;c_Mollicutes;o_RF39;f_unclassified | 25.9 | 16.5 | −0.516 (0.237) | 0.029 | 21.5 | 27 | 0.417 (0.211) | 0.049 | 3.8 × 10−4 |
| p_Tenericutes;c_Mollicutes;o_RF39;f_unclassified;g_unclassified | 25.9 | 16.5 | −0.516 (0.237) | 0.029 | 21.5 | 27 | 0.417 (0.211) | 0.049 | 3.8 × 10−4 |
| p_Tenericutes;c_Mollicutes;o_RF39;f_unclassified;g_unclassified;s_unclassified | 25.9 | 16.5 | −0.516 (0.237) | 0.029 | 21.5 | 27 | 0.417 (0.211) | 0.049 | 3.8 × 10−4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, J.; Yang, Y.; Xu, W.; Cai, H.; Wu, J.; Long, J.; Cai, Q.; Zheng, W.; Flynn, C.R.; Shu, X.-O.; et al. Sex-Specific Associations between Gut Microbiome and Non-Alcoholic Fatty Liver Disease among Urban Chinese Adults. Microorganisms 2021, 9, 2118. https://doi.org/10.3390/microorganisms9102118
Shi J, Yang Y, Xu W, Cai H, Wu J, Long J, Cai Q, Zheng W, Flynn CR, Shu X-O, et al. Sex-Specific Associations between Gut Microbiome and Non-Alcoholic Fatty Liver Disease among Urban Chinese Adults. Microorganisms. 2021; 9(10):2118. https://doi.org/10.3390/microorganisms9102118
Chicago/Turabian StyleShi, Jiajun, Yaohua Yang, Wanghong Xu, Hui Cai, Jie Wu, Jirong Long, Qiuyin Cai, Wei Zheng, Charles R. Flynn, Xiao-Ou Shu, and et al. 2021. "Sex-Specific Associations between Gut Microbiome and Non-Alcoholic Fatty Liver Disease among Urban Chinese Adults" Microorganisms 9, no. 10: 2118. https://doi.org/10.3390/microorganisms9102118
APA StyleShi, J., Yang, Y., Xu, W., Cai, H., Wu, J., Long, J., Cai, Q., Zheng, W., Flynn, C. R., Shu, X.-O., & Yu, D. (2021). Sex-Specific Associations between Gut Microbiome and Non-Alcoholic Fatty Liver Disease among Urban Chinese Adults. Microorganisms, 9(10), 2118. https://doi.org/10.3390/microorganisms9102118