Fatty Acids from Hermetia illucens Larvae Fat Inhibit the Proliferation and Growth of Actual Phytopathogens




Abstract
1. Introduction
2. Materials and Methods
2.1. BSFL Sample, Reagents, and Supply
2.2. Bacterial Strains and Culture Growth
2.3. BSFL Fat Extraction
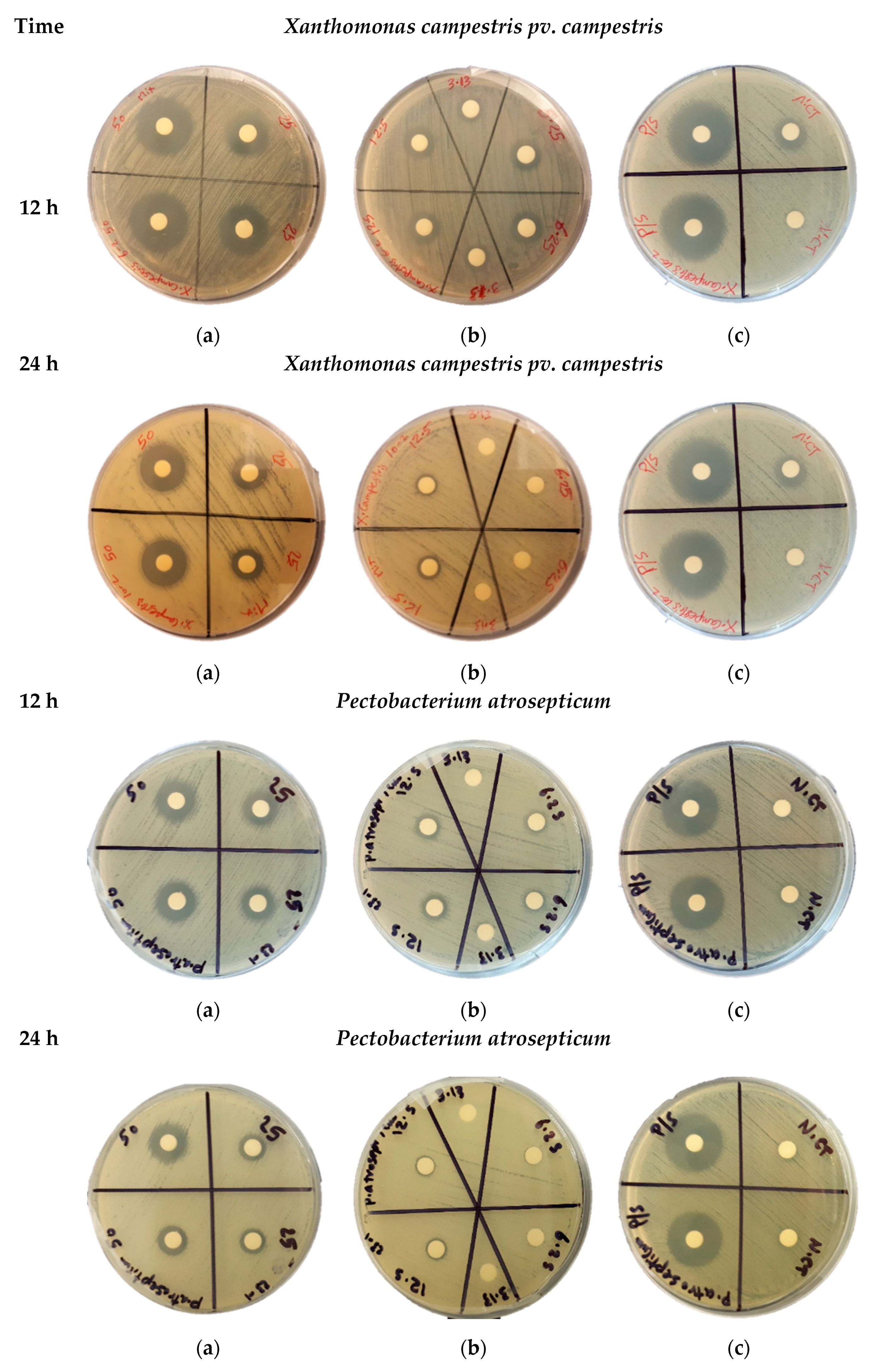
2.4. Agar Disk Diffusion Assay
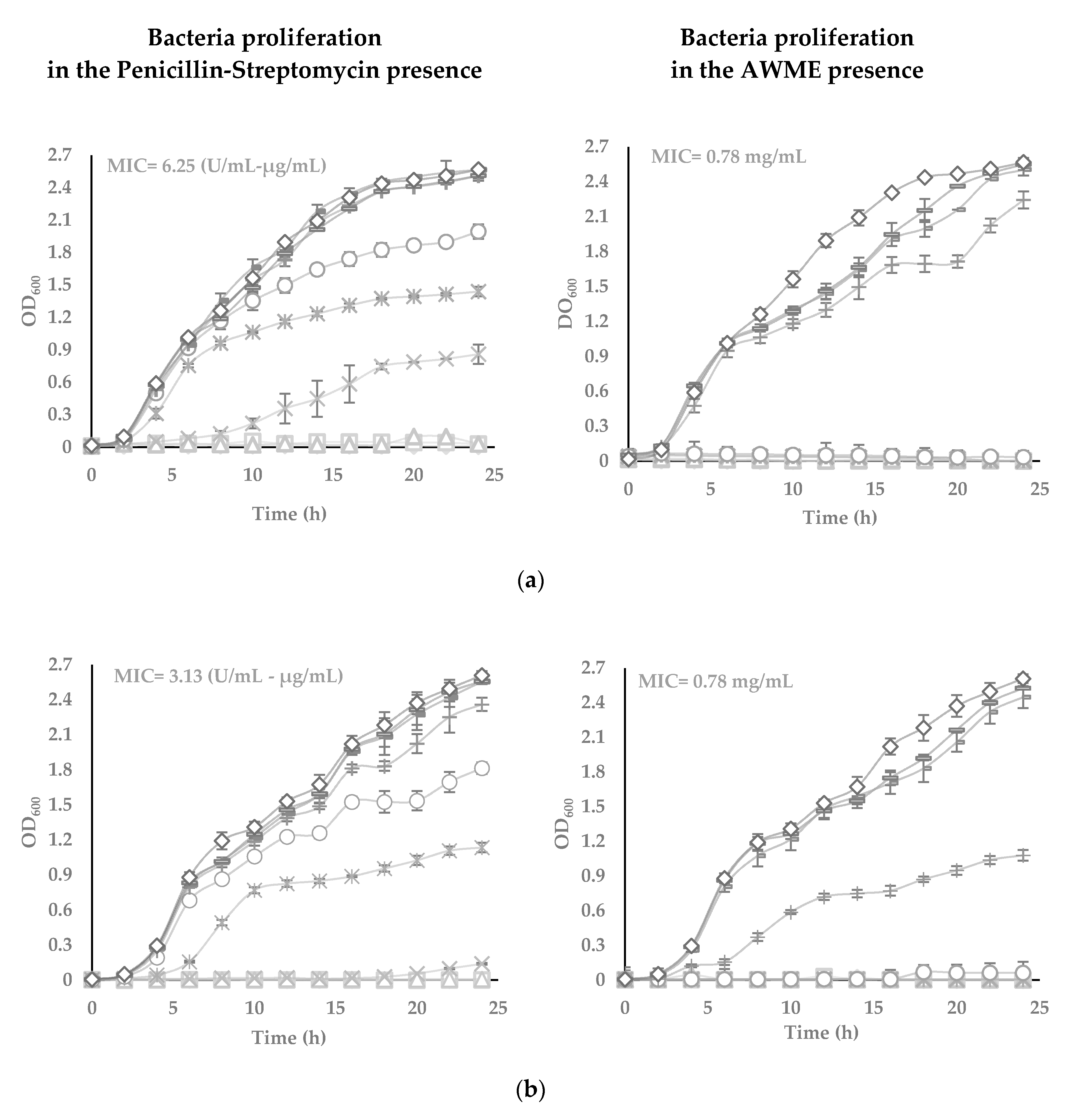
2.5. Turbidimetric Assay
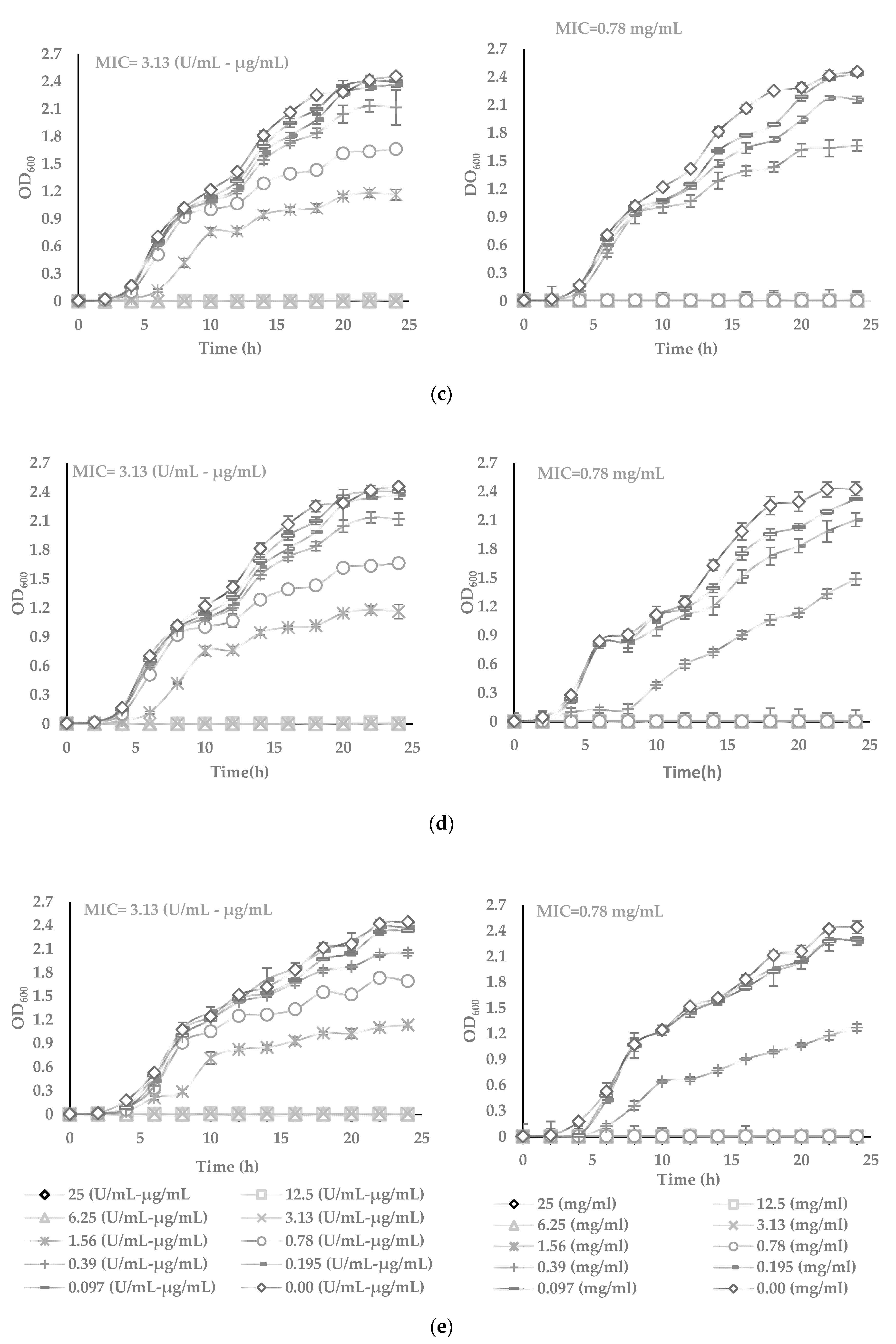
2.6. Determination of Minimum Inhibitory Concentrations by Turbidimetric Assay
2.7. Determination of Half of Inhibitory Concentration (IC50)
2.8. Determination of Minimum Bactericidal Concentration (MBC)
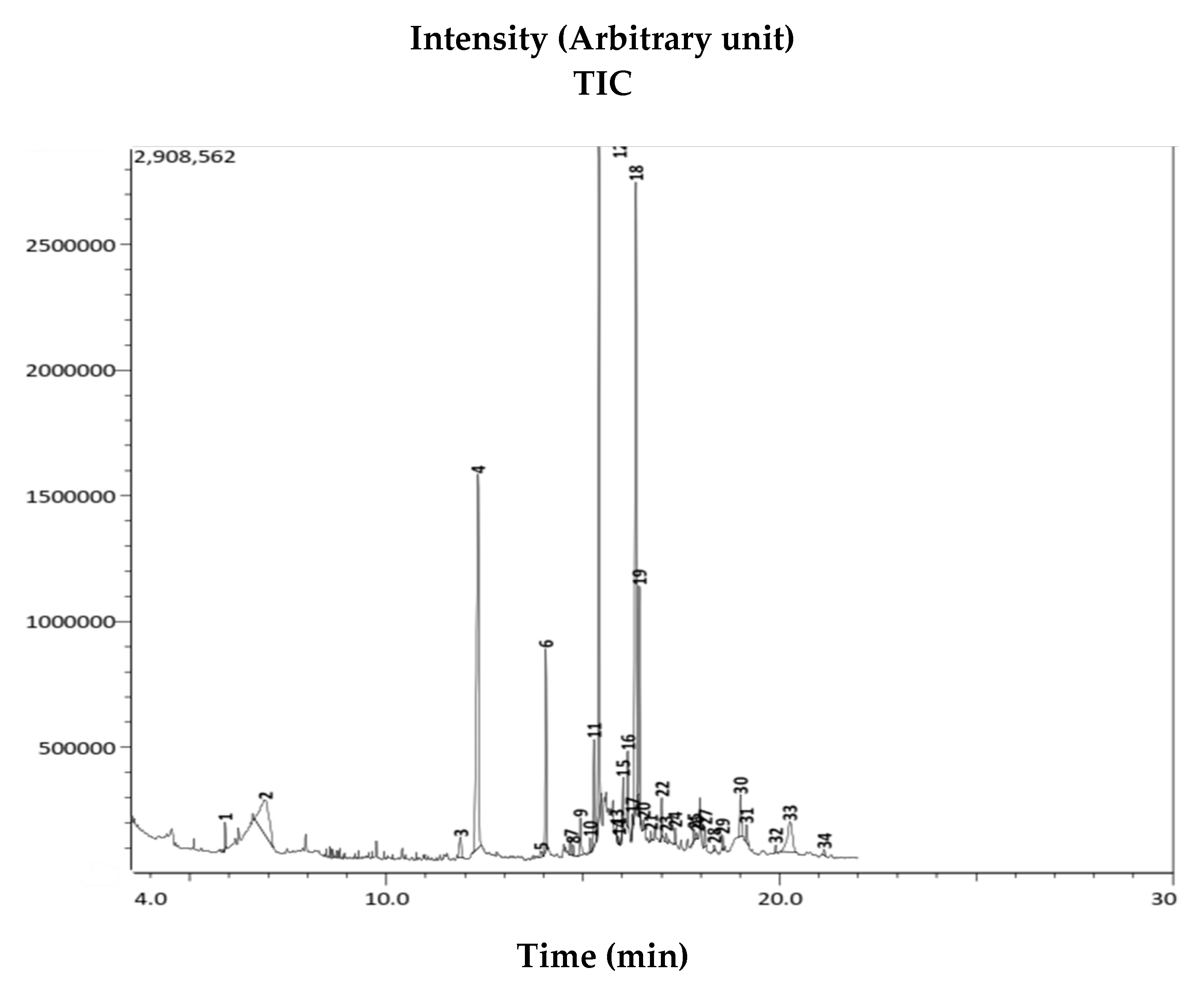
2.9. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
2.10. Statistical Analysis
3. Results
3.1. Extraction of Bioactive Compounds from the Hermetia illucens Larvae Fat
3.2. Examination of Antibacterial Activity against Phytopathogens
3.3. The Potency of AWME from Larvae Fat on Bacteria Growth and Proliferation in Turbidimetric Assay (TB)
3.4. Determination of the 50% Inhibitory Concentration (IC50) of the Plant Pathogenic Bacteria
3.5. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis of AWME
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kannan, V.; Bastas, K.; Devi, R. Scientific and Economic Impact of Plant Pathogenic Bacteria. In Sustainable Approaches to Controlling Plant Pathogenic Bacteria; Kannan, V., Bastas, K., Devi, R., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 369–392. [Google Scholar]
- Strange, R.N.; Scott, P.R. Plant Disease: A Threat to Global Food Security. Annu. Rev. Phytopathol. 2005, 43, 83–116. [Google Scholar] [CrossRef]
- Czajkowski, R.; Pérombelon, M.C.M.; Jafra, S.; Lojkowska, E.; Potrykus, M.; Van Der Wolf, J.M.; Sledz, W. Detection, identification and differentiation of Pectobacterium and Dickeya species causing potato blackleg and tuber soft rot: A review. Ann. Appl. Biol. 2015, 166, 18–38. [Google Scholar] [CrossRef]
- Pritchard, L.; Glover, R.H.; Humphris, S.; Elphinstone, J.G.; Toth, I.K. Genomics and taxonomy in diagnostics for food security: Soft-rotting enterobacterial plant pathogens. Anal. Methods 2016, 8, 12–24. [Google Scholar] [CrossRef]
- Anajjar, B.; Aitmhand, R.; Timinouni, M.; Ennaji, M.M. Characterization by PCR of two strains of Erwinia carotovora isolated from the potato rhizosphere in the region of greater Casablanca Casablanca in Morocco. EPPO Bull. 2007, 37, 175–180. [Google Scholar] [CrossRef]
- Cui, Y.; Chatterjee, A.; Yang, H.; Chatterjee, A.K. Regulatory network controlling extracellular proteins in Erwinia carotovora subsp. carotovora: FlhDC, the master regulator of flagellar genes, activates rsmB regulatory RNA production by affecting gacA and hexA (lrhA) expression. J. Bacteriol. 2008, 190, 4610–4623. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, A.R.; Potnis, N.; Timilsina, S.; Wilson, M.; Patané, J.; Martins, J.; Minsavage, G.V.; Dahlbeck, D.; Akhunova, A.; Almeida, N.; et al. Phylogenomics of Xanthomonas field strains infecting pepper and tomato reveals diversity in effector repertoires and identifies determinants of host specificity. Front. Microbiol. 2015, 6, 1–17. [Google Scholar] [CrossRef]
- Wulff, E.G.; Mguni, C.M.; Mortensen, C.N.; Keswani, C.L.; Hockenhull, J. Biological control of black rot (Xanthomonas campestris pv. campestris) of brassicas with an antagonistic strain of Bacillus subtilis in Zimbabwe. Eur. J. Plant Pathol. 2002, 108, 317–325. [Google Scholar] [CrossRef]
- Motyka, A.; Zoledowska, S.; Sledz, W.; Lojkowska, E. Molecular methods as tools to control plant diseases caused by Dickeya and Pectobacterium spp: A minireview. New Biotechnol. 2017, 39, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Czajkowski, R.; Pérombelon, M.C.M.; Van Veen, J.A.; Van der Wolf, J.M. Control of blackleg and tuber soft rot of potato caused by Pectobacterium and Dickeya species: A review. Plant Pathol. 2011, 60, 999–1013. [Google Scholar] [CrossRef]
- Zimnoch-Guzowska, E.; Lojkowska, M. Perombelon Resistance to Bacterial Pathogens. In Genetic Improvement of Solanaceous Crops Volume I: Potato; Razdan, M.K., Mattoo, A.K., Eds.; Science Publishers: Enfield, NH, USA, 2005; pp. 339–395. [Google Scholar]
- Aćimović, S.G.; Zeng, Q.; McGhee, G.C.; Sundin, G.W.; Wise, J.C. Control of fire blight (Erwinia amylovora) on apple trees with trunk-injected plant resistance inducers and antibiotics and assessment of induction of pathogenesis-related protein genes. Front. Plant Sci. 2015, 6, 1–10. [Google Scholar] [CrossRef]
- Hippler, F.W.R.; Boaretto, R.M.; Dovis, V.L.; Quaggio, J.A.; Azevedo, R.A.; Mattos-Jr, D. Oxidative stress induced by Cu nutritional disorders in Citrus depends on nitrogen and calcium availability. Sci. Rep. 2018, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Purschke, B.; Scheibelberger, R.; Axmann, S.; Adler, A.; Jäger, H. Impact of substrate contamination with mycotoxins, heavy metals and pesticides on the growth performance and composition of black soldier fly larvae (Hermetia illucens) for use in the feed and food value chain. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess 2017, 34, 1410–1420. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.S.; Tran, G.; Heuzé, V.; Ankers, P. State-of-the-art on use of insects as animal feed. Anim. Feed Sci. Technol. 2014, 197, 1–33. [Google Scholar] [CrossRef]
- Park, K.H.; Kwak, K.W.; Nam, S.H.; Choi, J.Y.; Hyun, S.; Kim, H.G.; Kim, S.H. Antibacterial activity of larval extract from the black soldier fly Hermetia illucens (Diptera: Stratiomyidae) against plant pathogens. J. Entomol. Zool. Stud. 2015, 3, 176–179. [Google Scholar] [CrossRef]
- Teh, C.H.; Nazni, W.A.; Lee, H.L.; Fairuz, A.; Tan, S.B.; Sofian-Azirun, M. In vitro antibacterial activity and physicochemical properties of a crude methanol extract of the larvae of the blow fly Lucilia cuprina. Med. Vet. Entomol. 2013, 27, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Ji, H.; Zhang, B.; Tian, J.; Zhou, J.; Yu, H. Influence of black soldier fly (Hermetia illucens) larvae oil on growth performance, body composition, tissue fatty acid composition and lipid deposition in juvenile Jian carp (Cyprinus carpio var. Jian). Aquaculture 2016, 465, 43–52. [Google Scholar] [CrossRef]
- Bauer, A.W.; Kirby, W.M.M.; Sherris, J.C.; Turck, M. Antibiotic Susceptibility Testing by a Standardized Single Disk Method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Thornsberry, C. Antimicrobial susceptibility testing: General considerations. In Manual of Clinical Microbiology, 5th ed.; BALOWS, A., Microbiology, A.S., Hausler, W.J., Jr., Herrmann, K.L., Isenberg, H.D., Shadomy, H.J., Eds.; American Society for Microbiology: Washington, DC, USA, 1991; pp. 1059–1201. ISBN 1555810292. [Google Scholar]
- Ordóñez, R.M.; Zampini, I.C.; Moreno, M.I.N.; Isla, M.I. Potential application of Northern Argentine propolis to control some phytopathogenic bacteria. Microbiol. Res. 2011, 166, 578–584. [Google Scholar] [CrossRef]
- Sledz, W.; Los, E.; Paczek, A.; Rischka, J.; Motyka, A.; Zoledowska, S.; Piosik, J.; Lojkowska, E. Antibacterial activity of caffeine against plant pathogenic bacteria. Acta Biochim. Pol. 2015, 62, 605–612. [Google Scholar] [CrossRef]
- Scrimgeour, C. Chemistry of fatty acids. In Bailey’s Industrial Oil and Fat Products; Shamsi, I.H., Shamsi, B.H., Jiang, L., Eds.; John Wiley and Sons, Inc.: New York, NY, USA, 2005; Volume 1, pp. 1–43. [Google Scholar]
- Ramos-Bueno, R.P.; González-Fernández, M.J.; Sánchez-Muros-Lozano, M.J.; García-Barroso, F.; Guil-Guerrero, J.L. Fatty acid profiles and cholesterol content of seven insect species assessed by several extraction systems. Eur. Food Res. Technol. 2016, 242, 1471–1477. [Google Scholar] [CrossRef]
- Caligiani, A.; Marseglia, A.; Sorci, A.; Bonzanini, F.; Lolli, V.; Maistrello, L.; Sforza, S. Influence of the killing method of the black soldier fly on its lipid composition. Food Res. Int. 2019, 116, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Agoramoorthy, G.; Chandrasekaran, M.; Venkatesalu, V.; Hsu, M.J. Antibacterial and antifungal activities of fatty acid methyl esters of the blind-your-eye mangrove from India. Braz. J. Microbiol. 2007, 38, 739–742. [Google Scholar] [CrossRef]
- Sivakumar, R.; Jebanesan, A.; Govindarajan, M.; Rajasekar, P. Larvicidal and repellent activity of tetradecanoic acid against Aedes aegypti (Linn.) and Culex quinquefasciatus (Say.) (Diptera:Culicidae). Asian Pac. J. Trop. Med. 2011, 4, 706–710. [Google Scholar] [CrossRef]
- Choi, W.H.; Jiang, M. Evaluation of antibacterial activity of hexanedioic acid isolated from Hermetia illucens larvae. J. Appl. Biomed. 2014, 12, 179–189. [Google Scholar] [CrossRef]
- Kim, S.A.; Rhee, M.S. Highly enhanced bactericidal effects of medium chain fatty acids (caprylic, capric, and lauric acid) combined with edible plant essential oils (carvacrol, eugenol, b-resorcylic acid, trans-cinnamaldehyde, thymol, and vanillin) against Escherichia coli O15. Food Control 2016, 60, 447–454. [Google Scholar] [CrossRef]
- Sugumaran, M. Comparative Biochemistry of Eumelanogenesis and the Protective Roles of Phenoloxidase and Melanin in Insects. Pigment Cell Res. 2002, 15, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Zeon, S.-R.; Lee, J.-G.; Choi, S.-H.; Shin, Y.K.; Park, K.-I. In vitro and in vivo efficacy of drugs against the protozoan parasite Azumiobodo hoyamushi that causes soft tunic syndrome in the edible ascidian Halocynthia roretzi (Drasche). J. Fish Dis. 2014, 37, 309–317. [Google Scholar] [CrossRef] [PubMed]
- Soetemans, L.; Uyttebroek, M.; D’Hondt, E.; Bastiaens, L. Use of organic acids to improve fractionation of the black soldier fly larvae juice into lipid- and protein-enriched fractions. Eur. Food Res. Technol. 2019, 245, 2257–2267. [Google Scholar] [CrossRef]
- Choi, W.H.; Yun, J.H.; Chu, J.P.; Chu, K.B. Antibacterial effect of extracts of hermetia illucens (diptera: Stratiomyidae) larvae against gram-negative bacteria. Entomol. Res. 2012, 42, 219–226. [Google Scholar] [CrossRef]
- Ushakova, N.A.; Brodskii, E.S.; Kovalenko, A.A.; Bastrakov, A.I.; Kozlova, A.A.; Pavlov, D.S. Characteristics of lipid fractions of larvae of the black soldier fly Hermetia illucens. Dokl. Biochem. Biophys. 2016, 468, 209–212. [Google Scholar] [CrossRef]
- Ewald, N.; Vidakovic, A.; Langeland, M.; Kiessling, A.; Sampels, S.; Lalander, C. Fatty acid composition of black soldier fly larvae (Hermetia illucens)—Possibilities and limitations for modification through diet. Waste Manag. 2020, 102, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Barroso, F.G.; Sánchez-Muros, M.J.; Segura, M.; Morote, E.; Torres, A.; Ramos, R.; Guil, J.L. Insects as food: Enrichment of larvae of Hermetia illucens with omega 3 fatty acids by means of dietary modifications. J. Food Compos. Anal. 2017, 62, 8–13. [Google Scholar] [CrossRef]
- Liland, N.S.; Biancarosa, I.; Araujo, P.; Biemans, D.; Bruckner, C.G.; Waagbø, R.; Torstensen, B.E.; Lock, E.-J.J. Modulation of nutrient composition of black soldier fly (Hermetia illucens) larvae by feeding seaweed-enriched media. PLoS ONE 2017, 12, 3188. [Google Scholar] [CrossRef] [PubMed]
- Meneguz, M.; Schiavone, A.; Gai, F.; Dama, A.; Lussiana, C.; Renna, M.; Gasco, L. Effect of rearing substrate on growth performance, waste reduction efficiency and chemical composition of black soldier fly (Hermetia illucens) larvae. J. Sci. Food Agric. 2018, 98, 5776–5784. [Google Scholar] [CrossRef]
- Spranghers, T.; Ottoboni, M.; Klootwijk, C.; Ovyn, A.; Deboosere, S.; De Meulenaer, B.; Michiels, J.; Eeckhout, M.; De Clercq, P.; De Smet, S. Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 2017, 97, 2594–2600. [Google Scholar] [CrossRef]
- Alanis, A.J. Resistance to antibiotics: Are we in the post-antibiotic era? Arch. Med. Res. 2005, 36, 697–705. [Google Scholar] [CrossRef]
- Kotan, R.; Cakir, A.; Dadasoglu, F.; Aydin, T.; Cakmakci, R.; Ozer, H.; Kordali, S.; Mete, E.; Dikbas, N. Antibacterial activities of essential oils and extracts of Turkish Achillea, Satureja and Thymus species against plant pathogenic bacteria. J. Sci. Food Agric. 2010, 90, 145–160. [Google Scholar] [CrossRef]
- Shea, K.M. Antibiotic resistance: What is the impact of agricultural uses of antibiotics on children’s health? Pediatrics 2003, 112, 253–258. [Google Scholar]
- Sumayo, M.S.; Kwon, D.K.; Ghim, S.Y. Linoleic acid-induced expression of defense genes and enzymes in tobacco. J. Plant Physiol. 2014, 171, 1757–1762. [Google Scholar] [CrossRef]
- Schlievert, P.M.; Peterson, M.L. Glycerol Monolaurate Antibacterial Activity in Broth and Biofilm Cultures. PLoS ONE 2012, 7, 350. [Google Scholar] [CrossRef]
- Skřivanová, E.; Molatová, Z.; Marounek, M. Effects of caprylic acid and triacylglycerols of both caprylic and capric acid in rabbits experimentally infected with enteropathogenic Escherichia coli O103. Vet. Microbiol. 2008, 126, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Y.; Gisi, U.; Mosinger, E. Systemic resistance of potato plants against Phytophthora infestans induced by unsaturated fatty acids. Physiol. Mol. Plant Pathol. 1991, 38, 255–263. [Google Scholar] [CrossRef]
- Blechert, S.; Brodschelm, W.; Hölder, S.; Kammerer, L.; Kutchan, T.M.; Mueller, M.J.; Xia, Z.Q.; Zenk, M.H. The octadecanoic pathway: Signal molecules for the regulation of secondary pathways. Proc. Natl. Acad. Sci. USA 1995, 92, 4099–4105. [Google Scholar] [CrossRef]
- Farmer, E.E.; Ryan, C.A. Octadecanoid Precursors of Jasmonic Acid Activate the Synthesis of Wound-Inducible Proteinase Inhibitors. Plant Cell 1992, 4, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.P.; Kumaravel, S.; Lalitha, C. Screening of antioxidant activity, total phenolics and GC-MS study of Vitex negundo. Afr. J. Biochem. Res. 2010, 4, 191–195. [Google Scholar]
- Awa, E.P.; Ibrahim, S.; Ameh, D.A. GC/MS Analysis and Antimicrobial activity of Diethyl ether fraction of Methanoolic extract from the Stem Bark of Annona senegalensis Pers. Int. J. Pharm. Sci. Res. 2012, 3, 4213–4218. [Google Scholar] [CrossRef]
- Ouattara, B.; Simard, R.E.; Holley, R.A.; Piette, G.J.P.; Bégin, A. Antibacterial activity of selected fatty acids and essential oils against six meat spoilage organisms. Int. J. Food Microbiol. 1997, 37, 155–162. [Google Scholar] [CrossRef]
- Nair, R.R. Agnihotra Yajna: A Prototype of South Asian Traditional Medical Knowledge. JAMS J. Acupunct. Meridian Stud. 2017, 10, 143–150. [Google Scholar] [CrossRef]
- Rahuman, A.A.; Gopalakrishnan, G.; Ghouse, B.S.; Arumugam, S.; Himalayan, B. Effect of Feronia limonia on mosquito larvae. Fitoterapia 2000, 71, 553–555. [Google Scholar] [CrossRef]
- Khalil, A.S.; Rahim, A.A.; Taha, K.K.; Abdallah, K.B. Characterization of Methanolic Extracts of Agarwood Leaves. J. Appl. Ind. Sci. 2013, 1, 78–88. [Google Scholar]
- Duke, J.A. Handbook of Biologically Active Phytochemicals and Their Activities; CRC Press, Inc.: Boca Raton, FL, USA, 1992; ISBN 9780849336706. [Google Scholar]
- Chandrasekaran, M.; Kannathasan, K.; Venkatesalu, V. Antimicrobial activity of fatty acid methyl esters of some members of chenopodiaceae. Z. Naturforsch. Sect. C J. Biosci. 2008, 63, 331–336. [Google Scholar] [CrossRef] [PubMed]
- Enig, M.G. Lauric oils as antimicrobial agents: Theory of effect, scientific rationale, and dietary application as adjunct nutritional support for HIV-infected individuals. In Nutrients and Foods in AIDS; Watson, R.R., Ed.; AI12/3/04; CRC Press: Boca Raton, FL, USA, 1998; pp. 81–97. [Google Scholar] [CrossRef]
- Pinto, M.E.A.; Araújo, S.G.; Morais, M.I.; Sá, N.P.; Lima, C.M.; Rosa, C.A.; Siqueira, E.P.; Johann, S.; Lima, L.A.R.S. Antifungal and antioxidant activity of fatty acid methyl esters from vegetable oils. An. Acad. Bras. Cienc. 2017, 89, 1671–1681. [Google Scholar] [CrossRef]
- Sahin, N.; Kula, I.; Erdogan, Y. Investigation of antimicrobial activities of nonanoic acid derivatives. Fresenius Environ. Bull. 2006, 15, 141–143. [Google Scholar]
- Chandrasekaran, M.; Senthilkumar, A.; Venkatesalu, V. Antibacterial and antifungal efficacy of fatty acid methyl esters from the leaves of Sesuvium portulacastrum L. Eur. Rev. Med. Pharmacol. Sci. 2011, 15, 775–780. [Google Scholar] [PubMed]
- Abou-Elela, G.M.; Abd-Elnaby, H.; Ibrahim, H.A.H.; Okbah, M.A. Marine Natural Products and Their Potential Applications as Anti-Infective Agents. World Appl. Sci. J. 2009, 7, 872–880. [Google Scholar]
- McGaw, L.J.; Jäger, A.K.; Van Staden, J. Antibacterial effects of fatty acids and related compounds from plants. S. Afr. J. Bot. 2002, 68, 417–423. [Google Scholar] [CrossRef]
- Kabara, J.J.; Scherr, G.H. Advances in Human Nutrition; Scherr, G.H., Ed.; Chem-Orbital: Park Forest, IL, USA, 1986; ISBN 0930376390. [Google Scholar]
- Feldlaufer, M.F.; Knox, D.A.; Lusby, W.R.; Shimanuki, H. Antimicrobial activity of fatty acids against Bacillus larvae, the causative agent of American foulbrood disease. Apidologie 1993, 24, 95–99. [Google Scholar] [CrossRef]
- Teh, C.H.; Nazni, W.A.; Nurulhusna, A.H.; Norazah, A.; Lee, H.L. Determination of antibacterial activity and minimum inhibitory concentration of larval extract of fly via resazurin-based turbidometric assay. BMC Microbiol. 2017, 17, 36. [Google Scholar] [CrossRef]
- Meziani, S.; Oomah, B.D.; Zaidi, F.; Simon-Levert, A.; Bertrand, C.; Zaidi-Yahiaoui, R. Antibacterial activity of carob (Ceratonia siliqua L.) extracts against phytopathogenic bacteria Pectobacterium atrosepticum. Microb. Pathog. 2015, 78, 95–102. [Google Scholar] [CrossRef]
- Canche-Escamilla, G.; Colli-Acevedo, P.; Borges-Argaez, R.; Quintana-Owen, P.; May-Crespo, J.F.; Cáceres-Farfan, M.; Yam Puc, J.A.; Sansores-Peraza, P.; Vera-Ku, B.M. Extraction of phenolic components from an Aloe vera (Aloe barbadensis Miller) crop and their potential as antimicrobials and textile dyes. Sustain. Chem. Pharm. 2019, 14, 1–8. [Google Scholar] [CrossRef]
- Soberón, J.R.; Sgariglia, M.A.; Dip Maderuelo, M.R.; Andina, M.L.; Sampietro, D.A.; Vattuone, M.A. Antibacterial activities of Ligaria cuneifolia and Jodina rhombifolia leaf extracts against phytopathogenic and clinical bacteria. J. Biosci. Bioeng. 2014, 118, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Hong, H.; Lee, J.H.; Kim, S.K. Phytochemicals and antioxidant capacity of some tropical edible plants. Asian Australas. J. Anim. Sci. 2018, 31, 1677–1684. [Google Scholar] [CrossRef] [PubMed]
- Halldor, T. Hilmarsson Hilmar Antimicrobial Lipids as Disinfectants, Antiseptics and Sanitizers. In Lipids and Essential Oils as Antimicrobial Agents; Thormar, H., Ed.; John Wiley & Sons Ltd.: West Sussex, UK, 2011; pp. 179–201. [Google Scholar]
- Stulnig, T.M.; Huber, J.; Leitinger, N.; Imre, E.M.; Angelisová, P.; Nowotny, P.; Waldhäusl, W. Polyunsaturated Eicosapentaenoic Acid Displaces Proteins from Membrane Rafts by Altering Raft Lipid Composition. J. Biol. Chem. 2001, 276, 37335–37340. [Google Scholar] [CrossRef] [PubMed]
- Zheng, C.J.; Yoo, J.S.; Lee, T.G.; Cho, H.Y.; Kim, Y.H.; Kim, W.G. Fatty acid synthesis is a target for antibacterial activity of unsaturated fatty acids. FEBS Lett. 2005, 579, 5157–5162. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BSFL Dilution Solvent | 10% Fat +H2O | 15% Fat +H2O | 20% Fat +H2O | 25% Fat +H2O | 30% Fat +H2O | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| CCl4 | + | ± | + | ± | + | ± | + | - - - | + | - - - |
| C6H14 | + | + | + | + | + | ± | + | - - - | ± | - - - |
| CH2Cl2 | + | ± | + | + | + | + | ± | - - - | ± | - - - |
| DMSO | ± | + | ± | + | ± | + | ± | - - - | - - - | - - - |
| CHCl3 | + | - - - | ± | ± | ± | ± | ± | - - - | ± | - - - |
| DMFA | + | ± | ± | ± | ± | ± | ± | - - - | ± | - - - |
| CH3CN | + | ± | ± | ± | ± | ± | ± | - - - | ± | - - - |
| CH3OH | ± | ± | - - - | - - - | - - - | - - - | - - - | - - - | - - - | - - - |
| C5H11OH | + | ± | + | ± | + | ± | + | - - - | + | - - - |
| C2H5OH | ± | + | ± | ± | ± | ± | ± | - - - | ± | - - - |
| C3H6O | ± | + | ± | + | ± | + | ± | - - - | ± | - - - |
| CHCl3 + DMFA (9:1, v/v) | + | ± | + | ± | + | ± | + | - - - | + | - - - |
| C6H14 + C3H6O (9:1, v/v) | + | ± | + | ± | + | ± | ± | - - - | ± | - - - |
| C6H14 + CH3CN (9:1, v/v) | + | ± | + | ± | + | ± | ± | - - - | ± | - - - |
| DMSO + C3H6O (9:1, v/v) | + | + | ± | + | ± | ± | ± | - - - | ± | - - - |
| DMSO + CH3CN (9:1, v/v) | + | + | ± | + | ± | + | ± | - - - | ± | - - - |
| CHCl3 + CH3CN (9:1, v/v) | + | ± | + | ± | + | ± | ± | - - - | ± | - - - |
| CHCl3 + C3H6O (9:1, v/v) | + | + | ± | ± | ± | ± | ± | - - - | ± | - - - |
| C5H11OH + C3H6O (9:1, v/v) | + | ± | + | ± | ± | ± | ± | - - - | ± | - - - |
| C5H11OH + DMSO (9:1, v/v) | ± | ± | ± | ± | ± | ± | - - - | - - - | - - - | - - - |
| Xcc | Pagg | Dsol | Pcc | Pba | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| AWME | 12 h | 24 h | 12 h | 24 h | 12 h | 24 h | 12 h | 24 h | 12 h | 24 h |
| 50 | 23.5 ± 035 | 19.0 ± 0.28 | 15.4 ± 0.28 | 12.5 ± 0.7 | 18.5 ± 0.7 | 14.0 ± 0.7 | 20.25 ± 1.76 | 14.75 ± 0.35 | 14.75 ± 0.75 | 12.2 ± 0.56 |
| 25 | 16.5 ± 0.42 | 12.5 ± 0.7 | 10.5 ± 0.7 | 8.5 ± 0.35 | 14.2 ± 0.28 | 11.0 ± 0.7 | 13.25 ± 0.35 | 10.12 ± 0.18 | 12.25 ± 0.35 | 9.5 ± 0.7 |
| 12.5 | 12.0 ± 0.35 | 8.0 ± 0.7 | 9.3 ± 0.21 | 7.0 ± 0.0 | 11.0 ± 0.7 | 9.0 ± 0.7 | 8.25 ± 0.35 | 7.0 ± 0.0 | 9.12 ± 0.18 | 7.12 ± 0.18 |
| 6.25 | 7.25 ± 0.28 | ND | 8.0 ± 0.7 | ND | 9.0 ± 0.7 | 7.0 ± 0.0 | 7.25 ± 0.35 | ND | 7.37 ± 0.18 | ND |
| 3.13 | ND | ND | 7.0 ± 0.0 | ND | 7.0 ± 0.0 | ND | ND | ND | ND | ND |
| P/S | 23.75 ± 0.35 | 24.0 ± 0.35 | 20.75 ± 0.35 | 21.0 ± 0.7 | 23.75 ± 0.7 | 23.75 ± 0.7 | 21.12 ± 0.18 | 21.0 ± 0.7 | 21.0 ± 0.7 | 20.63 ± 053 |
| AWM | 7.0 ± 0.0 | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| IC50 (µg/mL) of AWME against Treated Bacteria | IC50 (µg/mL) of Positive Control against Treated Bacteria | |||||
|---|---|---|---|---|---|---|
| Bacteria species | 6 h | 12 h | 24 h | 6 h | 12 h | 24 h |
| Xcc | 535.1 ± 0.16 | 477 ± 0.25 | 485.4 ± 0.4 | 1.941 ± 0.15 | 1.844 ± 0.26 | 1.893 ± 0.36 |
| Pagg | 299.5 ± 0.12 | 383.6 ± 0.22 | 366.8 ± 0.38 | 1.103 ± 0.13 | 1.638 ± 0.22 | 1.283 ± 0.37 |
| Dsol | 451.8 ± 0.07 | 455.1 ± 0.2 | 441.8 ± 0.36 | 1.060 ± 0.1 | 1.675 ± 0.19 | 1.306 ± 0.35 |
| Pcc | 317.6 ± 0.07 | 386.6 ± 0.22 | 431 ± 0.35 | 1.099 ± 0.13 | 1.782 ± 0.19 | 1.412 ± 0.35 |
| Pba | 290.3 ± 0.07 | 376.1 ± 0.22 | 400.2 ± 0.35 | 1.088 ± 0.07 | 1.630 ± 0.22 | 1.326 ± 0.35 |
| Peak Number | Retention Time | Content (%) | Compound Name (NIST Library) | Chemical Formula | Molecular Weight (g/Mol) | Similarity (%) |
|---|---|---|---|---|---|---|
| 1 | 5.9 | 0.61 | 1,2-Propanediol, 3-chloro | C3H7ClO2 | 110 | 93 |
| 2 | 6.92 | 6.88 | 1,2,3-Propantriol | C3H8O3 | 92 | 97 |
| 3 | 11.888 | 1.02 | beta.-D-Glucopyranose, 1,6-anhydro | C6H10O5 | 162 | 92 |
| 4 | 12.335 | 18.48 | Dodecanoic acid (lauric acid) | C12H24O2 | 200 | 97 |
| 5 | 13.335 | 0.3 | 2,4-Dodecadienal, (E,E)- (aromatic substance) | C12H20O | 180 | 80 |
| 6 | 14.059 | 5.59 | Tetradecanoic acid (myristic acid) | C14H28O2 | 228 | 97 |
| 7 | 14.682 | 0.41 | Dodecanoic acid, ethenyl ester (lauric acid vinyl ester | C14H26O2 | 226 | 81 |
| 8 | 14.753 | 0.24 | Pentadecanoic acid (saturated fatty acid) | C15H30O2 | 242 | 83 |
| 9 | 14.937 | 1.17 | 3-Cyclopentylpropionic acid, 2-dimethylaminoethyl ester | C12H23NO2 | 213 | 91 |
| 10 | 15.186 | 0.35 | Hexadecanoic acid methyl ester (palmetic acid methyl ester) | C17H34O2 | 270 | 88 |
| 11 | 15.286 | 3.02 | cis-9-Hexadecenoic acid (palmetoleic acid) | C16H30O2 | 254 | 96 |
| 12 | 15.412 | 20.34 | n-Hexadecanoic acid (palmetic acid) | C16H32O2 | 256 | 96 |
| 13 | 15.858 | 0.18 | Dodecanoic acid, 2-hydroxy-1-(hydroxymethyl) ethyl ester | C15H30O4 | 274 | 73 |
| 14 | 15.918 | 0.14 | Hexadecanoic acid | C16H32O2 | 256 | 78 |
| 15 | 16.02 | 1.32 | Dodecanoic acid, 2,3-dihydroxypropyl ester (monolaurin) | C15H30O4 | 274 | 83 |
| 16 | 16.138 | 1.62 | 9-Octadecenoic acid (Z)-, methyl ester (oleic acid methyl ester) | C19H36O2 | 296 | 87 |
| 17 | 16.257 | 0.34 | Octadecanoic acid, methyl ester (stearic acid methyl ester) | C19H38O2 | 298 | 86 |
| 18 | 16.347 | 22.22 | Octadec-9-eonic acid (oleic acid | C18H34O2 | 282 | 95 |
| 19 | 16.437 | 5.34 | Octadecanoic acid (stearic acid) | C18H36O2 | 284 | 94 |
| 20 | 16.537 | 0.27 | Linoleic acid ethyl ester | C20H36O2 | 308 | 75 |
| 21 | 16.721 | 0.23 | 9,12-Octadecadienoic acid (Z,Z)-(linoleic acid) | C18H32O2 | 280 | 89 |
| 22 | 17.002 | 0.97 | Fumaric acid, 2-dimethylaminoethyl heptadecyl ester | C25H47NO4 | 425 | 82 |
| 23 | 17.114 | 0.24 | Tetradecanoic acid, 2-hydroxy-1-(hydroxymethyl) ethyl ester | C17H34O4 | 304 | 86 |
| 24 | 17.346 | 0.36 | Eicosanoic acid (Arachidic acid) | C20H40O2 | 312 | 92 |
| 25 | 17.821 | 0.35 | Octanoic acid, 2-dimethylaminoethyl ester | C12H25NO2 | 215 | 87 |
| 26 | 17.902 | 0.15 | cis-9-Hexadecenal | C16H30O | 238 | 83 |
| 27 | 18.1 | 0.51 | Hexadecanoic acid, 2-hydroxy-1-(hydroxymethyl) ethyl ester | C19H38O4 | 330 | 94 |
| 28 | 18.336 | 0.15 | 9-Octadecanoic acid (Z)- | C18H34O2 | 282 | 83 |
| 29 | 18.551 | 0.33 | Tetradecanamide | C14H29NO | 227 | 91 |
| 30 | 19.011 | 1.45 | 9-Octadecenoic acid, 1,2,3-propanetriyl ester, (E,E,E)- | C57H104O6 | 884 | 93 |
| 31 | 19.162 | 0.68 | Oleoyl chloride | C18H33ClO | 300 | 90 |
| 32 | 19.905 | 0.22 | Octadecanamide | C18H37NO | 283 | 92 |
| 33 | 20.262 | 3.52 | Dodecanoic acid, 1,2,3-propanetriyl ester | C39H74O6 | 638 | 85 |
| 34 | 21.134 | 0.2 | Cholesterol, pentafluoropropionate | C30H45F5O2 | 532 | 88 |
| Name of Compounds | Content (%) | Biological Activity |
|---|---|---|
| Octadec-9-eonic acid (oleic acid) | 22.22 | Antibacterial [50] |
| n-Hexadecanoic acid (palmitic acid) | 20.34 | Antimicrobial [51] |
| Dodecanoic acid (lauric acid) | 18.48 | Antibacterial [52] |
| Tetradecanoic acid (myristic acid) | 5.59 | Antibacterial [26] |
| Octadecanoic acid (stearic acid) | 5.34 | Antimicrobial [53] |
| cis-9-Hexadecenoic acid (palmitoleic acid) | 3.02 | Antibacterial [54] |
| 1,2,3-Propantriol Hexadecanoic acid, 2-hydroxy-1-(hydroxymethyl) ethyl ester cis-9-Hexadecenal | 6.88 0.51 0.15 | Antimicrobial and antiseptic [55] Antimicrobial [55] Antimicrobial [55] |
| 9-Octadecenoic acid (Z)-, methyl ester (oleic acid methyl ester) | 1.62 | Antimicrobial [56] |
| Dodecanoic acid, 2,3-dihydroxypropyl ester (monolaurin) | 1.32 | Antimicrobial [57] |
| Eicosanoic acid (arachidic acid) | 0.36 | Antibacterial, antifungal, antioxidant [58,59] |
| Hexadecanoic acid methyl ester (palmitic acid methyl ester) | 0.35 | Antibacterial and antifungal [60] |
| Octadecanoic acid, methyl ester (stearic acid methyl ester) | 0.34 | Antimicrobial [61] |
| 9,12-Octadecadienoic acid (Z, Z)-(linoleic acid) | 0.23 | Antibacterial [62] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marusich, E.; Mohamed, H.; Afanasev, Y.; Leonov, S. Fatty Acids from Hermetia illucens Larvae Fat Inhibit the Proliferation and Growth of Actual Phytopathogens. Microorganisms 2020, 8, 1423. https://doi.org/10.3390/microorganisms8091423
Marusich E, Mohamed H, Afanasev Y, Leonov S. Fatty Acids from Hermetia illucens Larvae Fat Inhibit the Proliferation and Growth of Actual Phytopathogens. Microorganisms. 2020; 8(9):1423. https://doi.org/10.3390/microorganisms8091423
Chicago/Turabian StyleMarusich, Elena, Heakal Mohamed, Yuriy Afanasev, and Sergey Leonov. 2020. "Fatty Acids from Hermetia illucens Larvae Fat Inhibit the Proliferation and Growth of Actual Phytopathogens" Microorganisms 8, no. 9: 1423. https://doi.org/10.3390/microorganisms8091423
APA StyleMarusich, E., Mohamed, H., Afanasev, Y., & Leonov, S. (2020). Fatty Acids from Hermetia illucens Larvae Fat Inhibit the Proliferation and Growth of Actual Phytopathogens. Microorganisms, 8(9), 1423. https://doi.org/10.3390/microorganisms8091423