Biodegradation of Tetrahydrofuran by the Newly Isolated Filamentous Fungus Pseudallescheria boydii ZM01

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals, Media, and Growth Conditions
2.2. Isolation and Identification of the THF-Degrading Fungus
2.3. Biodegradation of THF by Strain ZM01
2.4. Effects of Different Factors on THF Biodegradation of Strain ZM01
2.5. Utilization of Different Substrates by Strain ZM01
2.6. Detection and Identification of THF and Related Metabolites
3. Results and Discussion
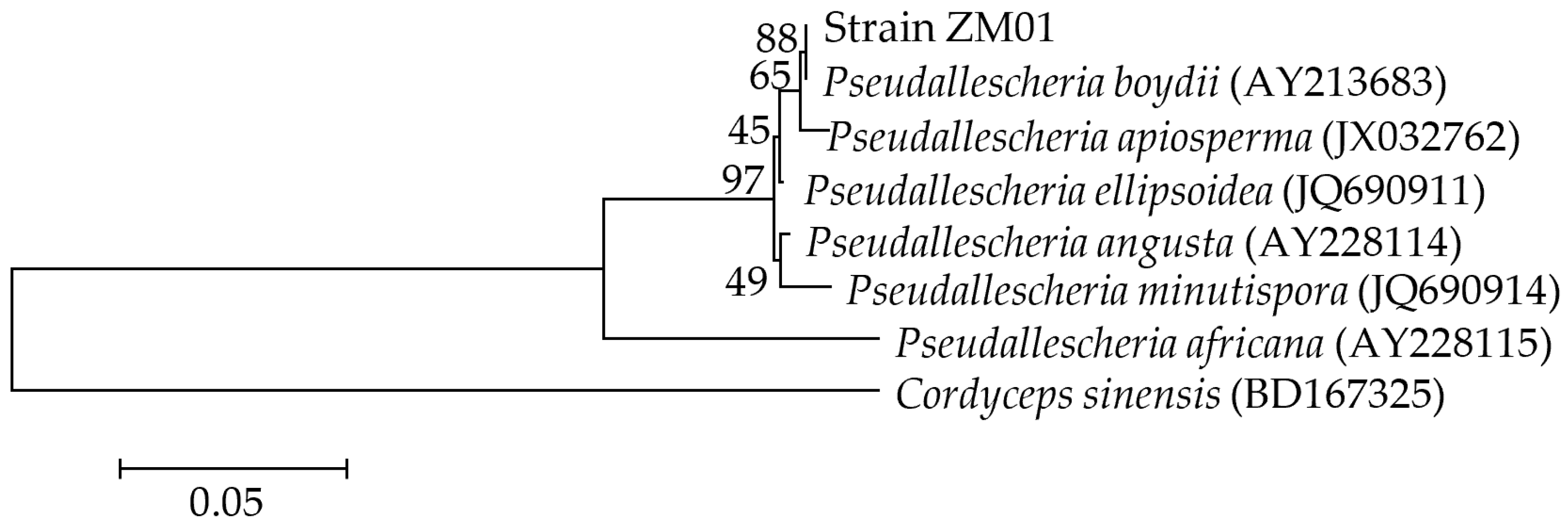
3.1. Isolation and Identification of the THF-Degrading Fungus
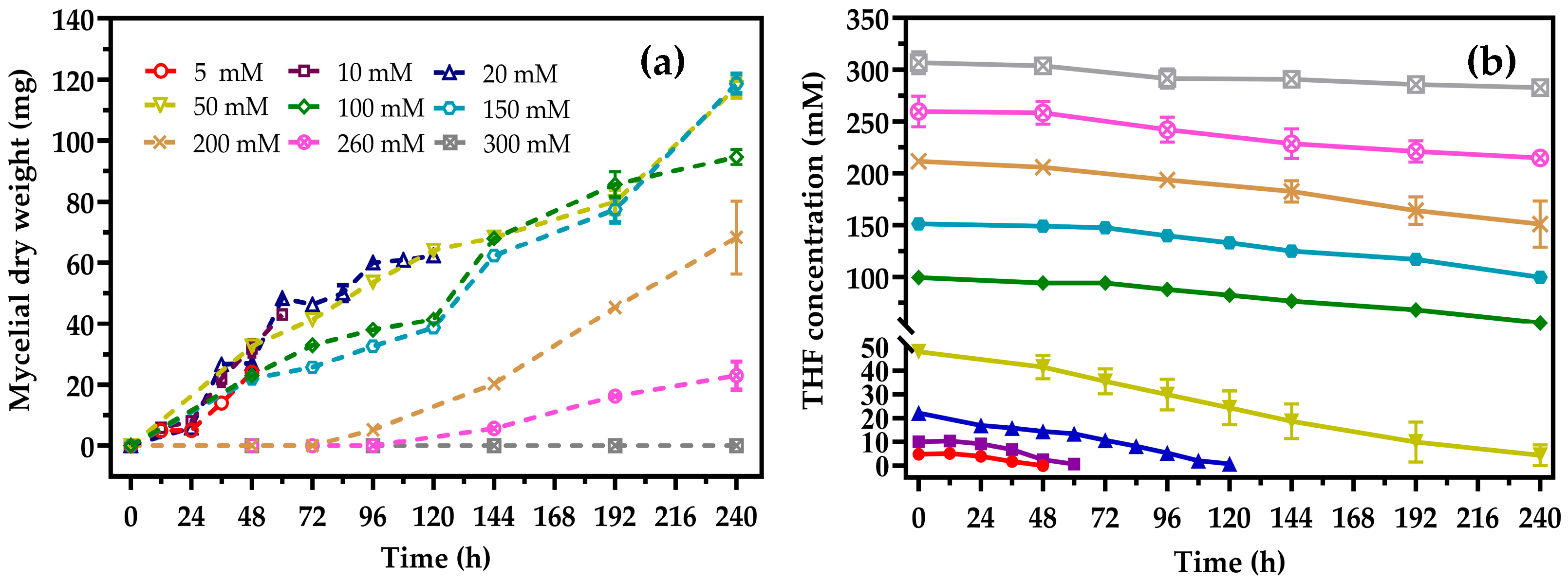
3.2. Degradation Characteristics of Strain ZM01
3.3. Effects of pH, Temperature, and Inoculum Size on THF Biodegradation
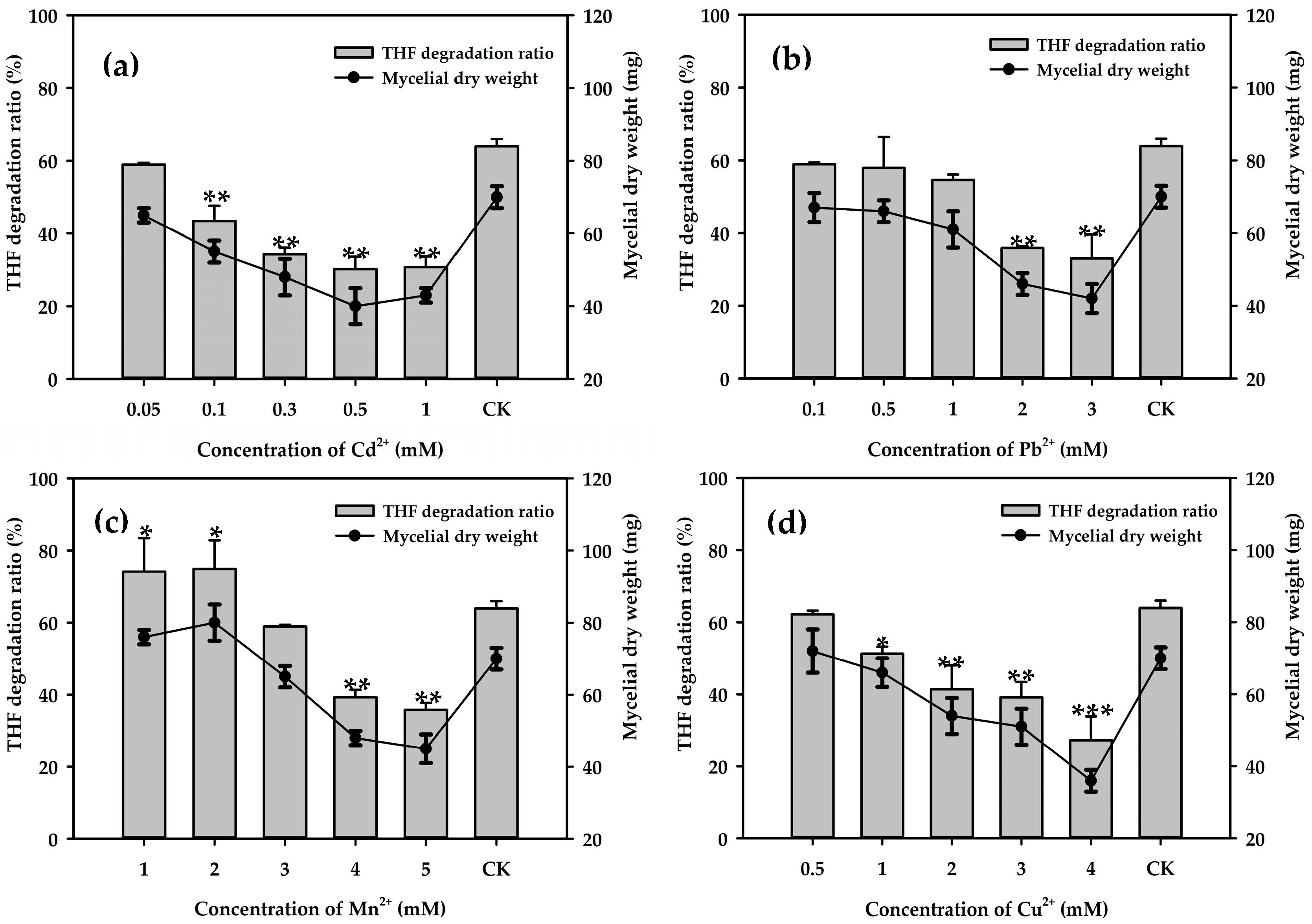
3.4. Effects of Metal Ions on THF Degradation
3.5. Degradation of Different Substrates by Strain ZM01
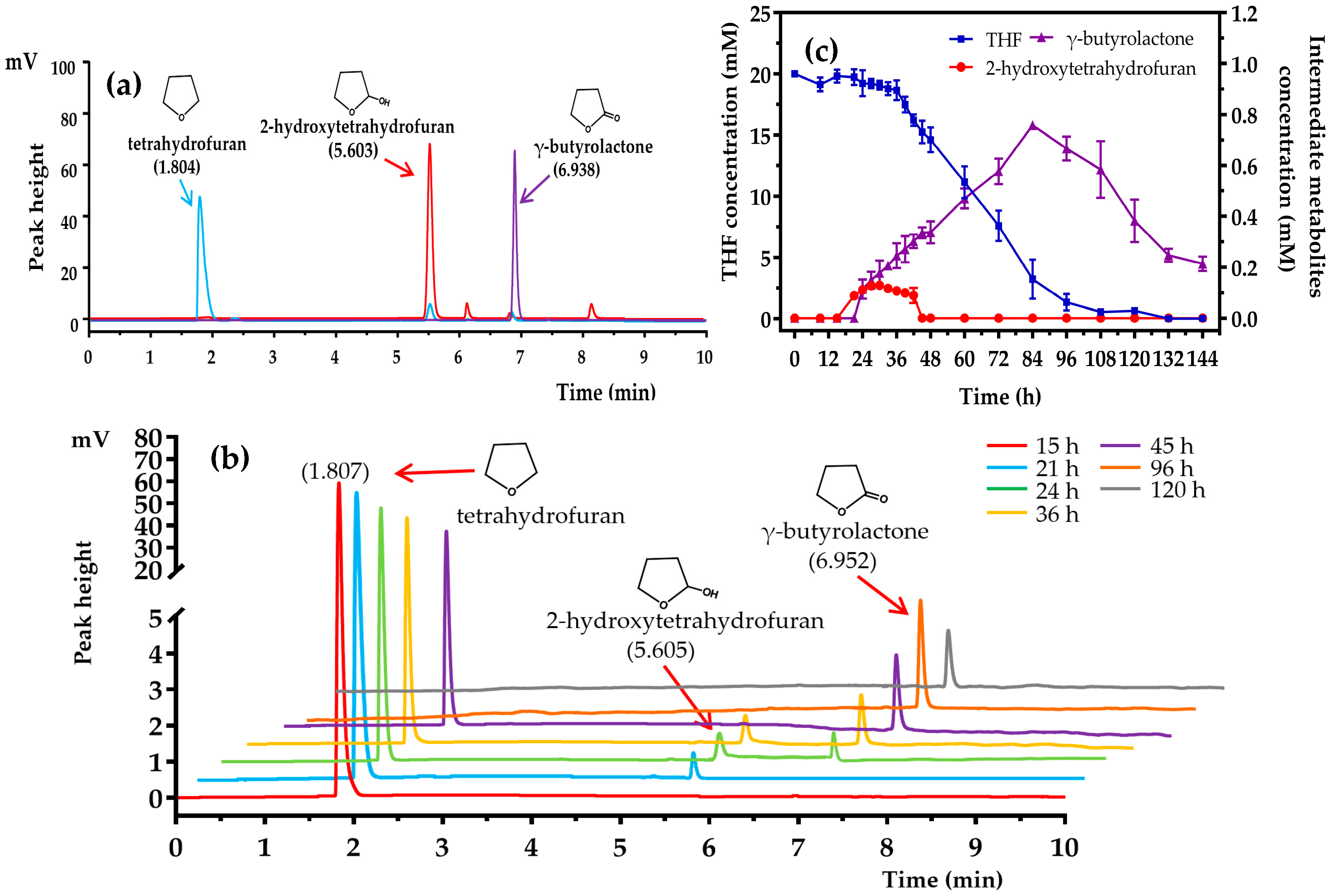
3.6. Proposed THF Degradation Pathway in Strain ZM01
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, J.M.; Zhou, Y.Y.; Chen, D.Z.; Jin, X.J. A newly isolated strain capable of effectively degrading tetrahydrofuran and its performance in a continuous flow system. Bioresour Technol. 2010, 101, 6461–6467. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Liu, C.; Li, H.; Lei, A. Direct functionalization of tetrahydrofuran and 1,4-dioxane: Nickel-catalyzed oxidative C(sp3)-H arylation. Angew. Chem. Int. Ed. Engl. 2013, 52, 4453–4456. [Google Scholar] [CrossRef] [PubMed]
- Isaacson, C.; Mohr, T.K.; Field, J.A. Quantitative determination of 1, 4-dioxane and tetrahydrofuran in groundwater by solid phase extraction GC/MS/MS. Environ. Sci. Technol. 2006, 40, 7305–7311. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Guan, J.; Tang, P.; Jiao, H.; Lin, C.; Wang, J.; Lu, Z.; Min, H.; Gao, H. Assessment of toxicity of tetrahydrofuran on the microbial community in activated sludge. Bioresour. Technol. 2010, 101, 5213–5221. [Google Scholar] [CrossRef]
- Gamer, A.O.; Jaeckh, R.; Leibold, E.; Kaufmann, W.; Gembardt, C.; Bahnemann, R.; van Ravenzwaay, B. Investigations on cell proliferation and enzyme induction in male rat kidney and female mouse liver caused by tetrahydrofuran. Toxicol. Sci. 2002, 70, 140–149. [Google Scholar] [CrossRef] [Green Version]
- Hermida, S.A.S.; Possari, E.P.M.; Souza, D.B.; Campos, I.P.D.; Gomes, O.F.; Di Mascio, P.; Medeiros, M.H.G.; Loureiro, A.P.M. 2′-Deoxyguanosine, 2′-deoxycytidine, and 2′-deoxyadenosine adducts resulting from the reaction of tetrahydrofuran with DNA bases. Chem. Res. Toxicol. 2006, 19, 927–936. [Google Scholar] [CrossRef]
- Malley, L.A.; Christoph, G.R.; Stadler, J.C.; Hansen, J.F.; Biesemeier, J.A.; Jasti, S.L. Acute and subchronic neurotoxicological evaluation of tetrahydrofuran by inhalation in rats. Drug Chem. Toxicol. 2001, 24, 201–219. [Google Scholar] [CrossRef]
- Asgher, M.; Bhatti, H.N.; Ashraf, M.; Legge, R.L. Recent developments in biodegradation of industrial pollutants by white rot fungi and their enzyme system. Biodegradation 2008, 19, 771–783. [Google Scholar] [CrossRef]
- Varjani, S.J. Microbial degradation of petroleum hydrocarbons. Bioresour. Technol. 2017, 223, 277–286. [Google Scholar] [CrossRef]
- Zhou, Y.Y.; Chen, D.Z.; Zhu, R.Y.; Chen, J.M. Substrate interactions during the biodegradation of BTEX and THF mixtures by Pseudomonas oleovorans DT4. Bioresour. Technol. 2011, 102, 6644–6649. [Google Scholar] [CrossRef]
- Sei, K.; Miyagaki, K.; Kakinoki, T.; Fukugasako, K.; Inoue, D.; Ike, M. Isolation and characterization of bacterial strains that have high ability to degrade 1,4-dioxane as a sole carbon and energy source. Biodegradation 2013, 24, 665–674. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Mathieu, J.; Yang, Y.; Yu, P.F.; da Silva, M.L.B.; Alvarez, P.J.J. 1,4-Dioxane biodegradation by Mycobacterium dioxanotrophicus PH-06 is associated with a group-6 soluble di-iron monooxygenase. Environ. Sci. Tech. Let. 2017, 4, 494–499. [Google Scholar] [CrossRef] [Green Version]
- Daye, K.J.; Groff, J.C.; Kirpekar, A.C.; Mazumder, R. High efficiency degradation of tetrahydrofuran (THF) using a membrane bioreactor: Identification of THF-degrading cultures of Pseudonocardia sp. strain M1 and Rhodococcus ruber isolate M2. J. Ind. Microbiol. Biot. 2003, 30, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.Z.; Ko, K.; Ramsay, J.A. Biodegradation of 1,4-dioxane by a Flavobacterium. Biodegradation 2011, 22, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Inoue, D.; Tsunoda, T.; Sawada, K.; Yamamoto, N.; Saito, Y.; Sei, K.; Ike, M. 1,4-Dioxane degradation potential of members of the genera Pseudonocardia and Rhodococcus. Biodegradation 2016, 27, 277–286. [Google Scholar] [CrossRef]
- Yao, Y.L.; Lv, Z.M.; Min, H.; Lv, Z.H.; Jiao, H.P. Isolation, identification and characterization of a novel Rhodococcus sp. strain in biodegradation of tetrahydrofuran and its medium optimization using sequential statistics-based experimental designs. Bioresour. Technol. 2009, 100, 2762–2769. [Google Scholar] [CrossRef]
- Tajima, T.; Hayashida, N.; Matsumura, R.; Omura, A.; Nakashimada, Y.; Kato, J. Isolation and characterization of tetrahydrofuran-degrading Rhodococcus aetherivorans strain M8. Process. Biochem. 2012, 47, 1665–1669. [Google Scholar] [CrossRef]
- Niu, L.H.; Li, Y.; Xu, L.L.; Wang, P.F.; Zhang, W.L.; Wang, C.; Cai, W.; Wang, L.Q. Ignored fungal community in activated sludge wastewater treatment plants: Diversity and altitudinal characteristics. Environ. Sci. Pollut. Res. 2017, 24, 4185–4193. [Google Scholar] [CrossRef]
- Stajich, J.E.; Berbee, M.L.; Blackwell, M.; Hibbett, D.S.; James, T.Y.; Spatafora, J.W.; Taylor, J.W. The Fungi. Curr. Biol. 2009, 19, 840–845. [Google Scholar] [CrossRef] [Green Version]
- Li, X.J.; Li, Y.; Zhao, X.D.; Zhang, X.L.; Zhao, Q.; Wang, X.; Li, Y.T. Restructured fungal community diversity and biological interactions promote metolachlor biodegradation in soil microbial fuel cells. Chemosphere 2019, 221, 735–749. [Google Scholar] [CrossRef]
- More, T.T.; Yan, S.; Tyagi, R.D.; Surampalli, R.Y. Potential use of filamentous fungi for wastewater sludge treatment. Bioresour. Technol. 2010, 101, 7691–7700. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Zheng, G.Y.; Zhou, L.X. Degradation of slime extracellular polymeric substances and inhibited sludge flocs destruction contribute to sludge dewaterability enhancement during fungal treatment of sludge using filamentous fungus Mucor sp. GY-1. Bioresour. Technol. 2015, 192, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Das, B.K.; Roy, A.; Koschorreck, M.; Mandal, S.M.; Wendt-Potthoff, K.; Bhattacharya, J. Occurrence and role of algae and fungi in acid mine drainage environment with special reference to metals and sulfate immobilization. Water Res. 2009, 43, 883–894. [Google Scholar] [PubMed]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Dijksterhuis, J. Fungal spores: Highly variable and stress-resistant vehicles for distribution and spoilage. Food Microbiol. 2019, 81, 2–11. [Google Scholar] [CrossRef] [PubMed]
- Nakamiya, K.; Hashimoto, S.; Ito, H.; Edmonds, J.S.; Morita, M. Degradation of 1,4-dioxane and cyclic ethers by an isolated fungus. Appl. Environ. Microbiol. 2005, 71, 1254–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, K.; Cuiffetti, L.; Hyman, M. Metabolism and cometabolism of cyclic ethers by a filamentous fungus, a Graphium sp. Appl. Environ. Microbiol. 2009, 75, 5514–5522. [Google Scholar] [CrossRef] [Green Version]
- Trippe, K.M.; Wolpert, T.J.; Hyman, M.R.; Ciuffetti, L.M. RNAi silencing of a cytochrome P450 monoxygenase disrupts the ability of a filamentous fungus, Graphium sp., to grow on short-chain gaseous alkanes and ethers. Biodegradation 2014, 25, 137–151. [Google Scholar] [CrossRef]
- Moreno-Horn, M.; Garbe, L.A.; Tressl, R.; Gorisch, H. Transient accumulation of gamma-butyrolactone during degradation of bis(4-chloro-n-butyl) ether by diethylether-grown Rhodococcus sp. strain DTB. Appl. Microbiol. Biotechnol. 2005, 69, 335–340. [Google Scholar] [CrossRef]
- Kohlweyer, U.; Thiemer, B.; Schrader, T.; Andreesen, J.R. Tetrahydrofuran degradation by a newly isolated culture of Pseudonocardia sp. strain K1. FEMS Microbiol. Lett. 2000, 186, 301–306. [Google Scholar] [CrossRef]
- Kim, Y.M.; Jeon, J.R.; Murugesan, K.; Kim, E.J.; Chang, Y.S. Biodegradation of 1,4-dioxane and transformation of related cyclic compounds by a newly isolated Mycobacterium sp. PH-06. Biodegradation 2009, 20, 511–519. [Google Scholar] [CrossRef] [PubMed]
- Sales, C.M.; Grostern, A.; Parales, J.V.; Parales, R.E.; Alvarez-Cohen, L. Oxidation of the cyclic ethers 1,4-dioxane and tetrahydrofuran by a monooxygenase in two Pseudonocardia species. Appl. Environ. Microbiol. 2013, 79, 7702–7708. [Google Scholar] [CrossRef] [Green Version]
- Curry, S.; Ciuffetti, L.; Hyman, M. Inhibition of growth of a Graphium sp. on gaseous n-alkanes by gaseous n-alkynes and n-alkenes. Appl. Environ. Microbiol. 1996, 62, 2198–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.B.; Huang, H.; Qi, M.B.; Wang, X.J.; Adebanjo, O.O.; Lu, Z.M. Metabolite cross-feeding between Rhodococcus ruber YYL and Bacillus cereus MLY1 in the biodegradation of tetrahydrofuran under pH stress. Appl. Environ. Microb. 2019, 85, e01196-19. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.J.; Choi, J.W.; Choi, N.C.; Mahendran, B.; Lee, C.E. Modeling of growth kinetics for Pseudomonas spp. during benzene degradation. Appl. Microbiol. Biot. 2005, 69, 456–462. [Google Scholar] [CrossRef]
- Chen, D.Z.; Jin, X.J.; Chen, J.; Ye, J.X.; Jiang, N.X.; Chen, J.M. Intermediates and substrate interaction of 1,4-dioxane degradation by the effective metabolizer Xanthobacter flavus DT8. Int. Biodeter. Biodegr. 2016, 106, 133–140. [Google Scholar] [CrossRef]
- Liu, Z.B.; He, Z.X.; Huang, H.; Ran, X.B.; Oluwafunmilayo, A.O.; Lu, Z.M. pH Stress-induced cooperation between Rhodococcus ruber YYL and Bacillus cereus MLY1 in biodegradation of tetrahydrofuran. Front. Microbiol. 2017, 8, 2297. [Google Scholar] [CrossRef] [Green Version]
- Ji, J.; Kakade, A.; Zhang, R.L.; Zhao, S.; Khan, A.; Liu, P.; Li, X.K. Alcohol ethoxylate degradation of activated sludge is enhanced by bioaugmentation with Pseudomonas sp. LZ-B. Ecotoxicol. Environ. Saf. 2019, 169, 335–343. [Google Scholar] [CrossRef]
- Manai, I.; Miladi, B.; El Mselmi, A.; Hamdi, M.; Bouallagui, H. Improvement of activated sludge resistance to shock loading by fungal enzyme addition during textile wastewater treatment. Environ. Technol. 2017, 38, 880–890. [Google Scholar] [CrossRef]
- Inoue, D.; Tsunoda, T.; Yamamoto, N.; Ike, M.; Sei, K. 1,4-Dioxane degradation characteristics of Rhodococcus aetherivorans JCM 14343. Biodegradation 2018, 29, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Fiorenza, S.; Chatham, J.R.; Mahendra, S.; Alvarez, P.J.J. 1,4-Dioxane biodegradation at low temperatures in arctic groundwater samples. Water Res. 2010, 44, 2894–2900. [Google Scholar] [PubMed] [Green Version]
- Kuo, C.W.; Genthner, B.R.S. Effect of added heavy metal ions on biotransformation and biodegradation of 2-chlorophenol and 3-chlorobenzoate in anaerobic bacterial consortia. Appl. Environ. Microb. 1996, 62, 2317–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandrin, T.R.; Maier, R.M. Impact of metals on the biodegradation of organic pollutants. Environ. Health Persp. 2003, 111, 1093–1101. [Google Scholar] [CrossRef]
- Pornwongthong, P.; Mulchandani, A.; Gedalanga, P.B.; Mahendra, S. Transition metals and organic ligands influence biodegradation of 1,4-dioxane. Appl. Biochem. Biotech. 2014, 173, 291–306. [Google Scholar] [CrossRef]
- Zhao, L.D.; Lu, X.; Polasko, A.; Johnson, N.W.; Miao, Y.; Yang, Z.M.; Mahendra, S.; Gu, B.H. Co-contaminant effects on 1,4-dioxane biodegradation in packed soil column flow-through systems. Environ. Pollut. 2018, 243, 573–581. [Google Scholar] [CrossRef]
- Huang, H.; Yu, H.; Qi, M.; Liu, Z.; Wang, H.; Lu, Z. Enrichment and characterization of a highly efficient tetrahydrofuran-degrading bacterial culture. Biodegradation 2019, 30, 467–479. [Google Scholar] [CrossRef]
- Parales, R.E.; Adamus, J.E.; White, N.; May, H.D. Degradation of 1,4-dioxane by an actinomycete in pure culture. Appl. Environ. Microb. 1994, 60, 4527–4530. [Google Scholar] [CrossRef] [Green Version]
- Masuda, H.; McClay, K.; Steffan, R.J.; Zylstra, G.J. Biodegradation of tetrahydrofuran and 1,4-dioxane by soluble diiron monooxygenase in Pseudonocardia sp. strain ENV478. J. Mol. Microb Biotech. 2012, 22, 312–316. [Google Scholar] [CrossRef]
- Xiong, Y.; Mason, O.U.; Lowe, A.; Zhou, C.; Chen, G.; Tang, Y. Microbial community analysis provides insights into the effects of tetrahydrofuran on 1,4-dioxane biodegradation. Appl. Environ. Microbiol. 2019, 85, e00244-19. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | Growth a (mg) | Substrate | Growth a (mg) |
|---|---|---|---|
| Tetrahydrofuran | 63.3 ± 2.5 | 1,4-Dioxane | 8.0 ± 1.0 |
| 3-Hydroxytetrahydrofuran | 8.0 ± 1.0 | Pyridine | NG |
| Dimethylfuran | 6.2 ± 0.5 | Ethyl acetate | 17.7 ± 1.5 |
| γ-Butyrolactone | 53.7 ± 1.5 | Methyl alcohol | NG |
| 1,4-Butanediol | 71.7 ± 1.5 | n-Hexane | 8.7 ± 0.6 |
| Benzene | 8.3 ± 0.6 | Acetone | NG |
| Methylbenzene | 7.7 ± 0.6 | 2-hydroxytetrahydrofuran | 58.9 ± 2.1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ren, H.; Li, H.; Wang, H.; Huang, H.; Lu, Z. Biodegradation of Tetrahydrofuran by the Newly Isolated Filamentous Fungus Pseudallescheria boydii ZM01. Microorganisms 2020, 8, 1190. https://doi.org/10.3390/microorganisms8081190
Ren H, Li H, Wang H, Huang H, Lu Z. Biodegradation of Tetrahydrofuran by the Newly Isolated Filamentous Fungus Pseudallescheria boydii ZM01. Microorganisms. 2020; 8(8):1190. https://doi.org/10.3390/microorganisms8081190
Chicago/Turabian StyleRen, Hao, Hanbo Li, Haixia Wang, Hui Huang, and Zhenmei Lu. 2020. "Biodegradation of Tetrahydrofuran by the Newly Isolated Filamentous Fungus Pseudallescheria boydii ZM01" Microorganisms 8, no. 8: 1190. https://doi.org/10.3390/microorganisms8081190
APA StyleRen, H., Li, H., Wang, H., Huang, H., & Lu, Z. (2020). Biodegradation of Tetrahydrofuran by the Newly Isolated Filamentous Fungus Pseudallescheria boydii ZM01. Microorganisms, 8(8), 1190. https://doi.org/10.3390/microorganisms8081190