Mechanisms of Linezolid Resistance Among Enterococci of Clinical Origin in Spain—Detection of optrA- and cfr(D)-Carrying E. faecalis

 , , ,
, , ,  and
and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Bacterial Collection
2.2. Antimicrobial Resistance Phenotype
2.3. Mechanisms of Linezolid Resistance
2.4. Molecular Characterization
2.5. Clonal Relatedness
2.6. Conjugation Assays
3. Results
3.1. Mechanisms of Oxazolidinone Resistance
3.2. Molecular Characterization and Clonal Relatedness
3.3. Resistance to Non-Oxazolidinone Agents
3.4. Virulence Profile
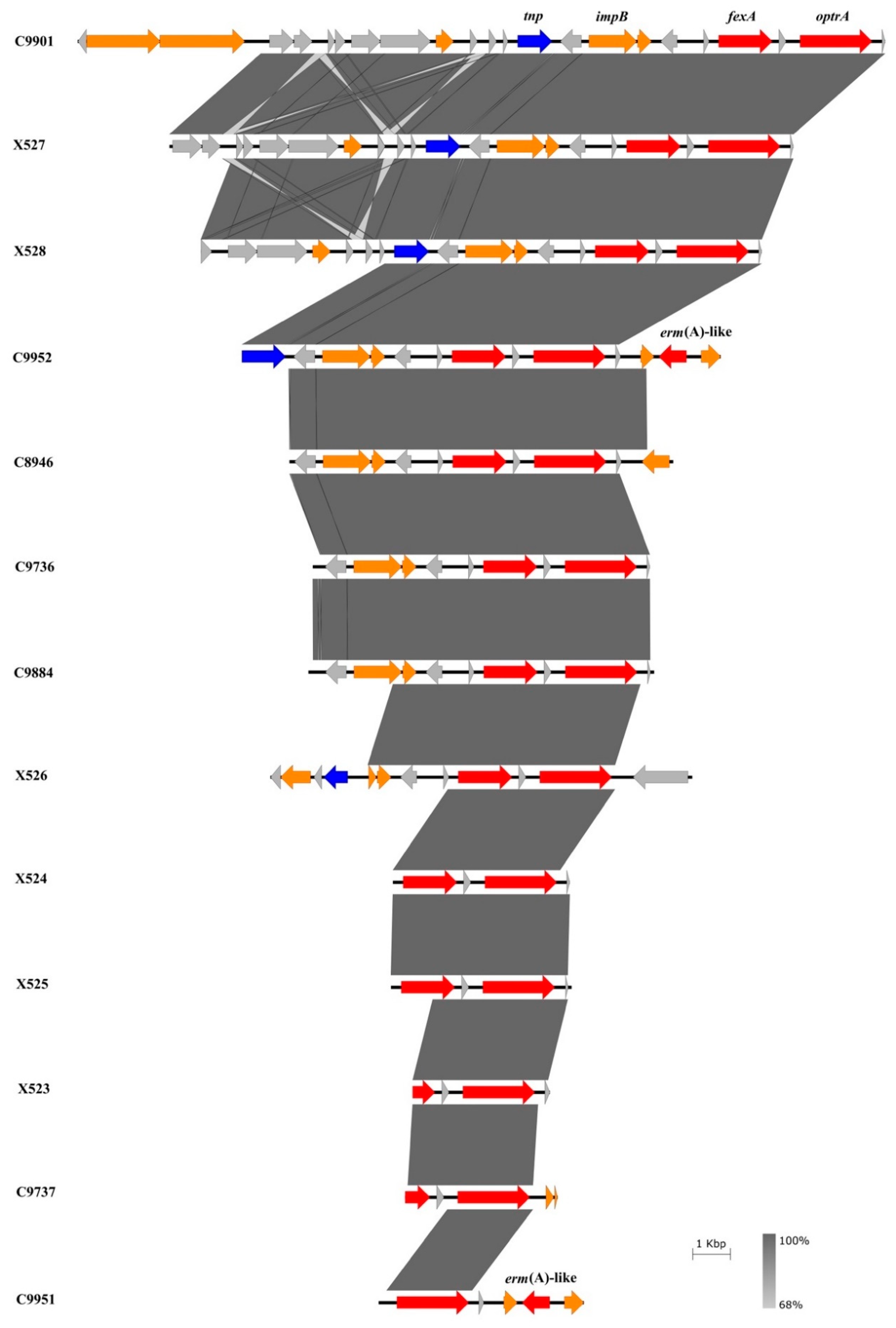
3.5. Genetic Environment of the optrA and cfr(D) Genes
3.6. Transferability of the optrA and cfr(D) Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; León-Sampedro, R.; Del Campo, R.; Coque, T.M. Antimicrobial resistance in Enterococcus spp. of animal origin. Microbiol. Spectr. 2018, 6, 185–227. [Google Scholar] [CrossRef]
- Batts, D.H. Linezolid—A new option for treating gram-positive infections. Oncology 2000, 14, 23–29. [Google Scholar] [PubMed]
- Mendes, R.E.; Deshpande, L.M.; Jones, R.N. Linezolid update: Stable in vitro activity following more than a decade of clinical use and summary of associated resistance mechanisms. Drug Resist. Updat. 2014, 17, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Bi, R.; Qin, T.; Fan, W.; Ma, P.; Gu, B. The emerging problem of linezolid-resistant enterococci. J. Glob. Antimicrob. Resist. 2018, 13, 11–19. [Google Scholar] [CrossRef]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti Infect. Ther. 2014, 12, 1221–1236. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Schwarz, S.; Jacobsen, L.; Hansen, L.H.; Vester, B. A new mechanism for chloramphenicol, florfenicol and clindamycin resistance: Methylation of 23S ribosomal RNA at A2503. Mol. Microbiol. 2005, 57, 1064–1073. [Google Scholar] [CrossRef]
- Schwarz, S.; Werckenthin, C.; Kehrenberg, C. Identification of a plasmid-borne chloramphenicol-florfenicol resistance gene in Staphylococcus sciuri. Antimicrob. Agents Chemother. 2000, 44, 2530–2533. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Wang, Y.; Schwarz, S. Presence and dissemination of the multiresistance gene cfr in Gram-positive and Gram-negative bacteria. J. Antimicrob. Chemother. 2013, 68, 1697–1706. [Google Scholar] [CrossRef]
- Deshpande, L.M.; Ashcraft, D.S.; Kahn, H.P.; Pankey, G.; Jones, R.N.; Farrell, D.J.; Mendes, R.E. Detection of a new cfr-like gene, cfr (B), in Enterococcus faecium isolates recovered from human specimens in the United States as part of the SENTRY Antimicrobial Surveillance Program. Antimicrob. Agents Chemother. 2015, 59, 6256–6261. [Google Scholar] [CrossRef] [Green Version]
- Kuroda, M.; Sekizuka, T.; Matsui, H.; Suzuki, K.; Seki, H.; Saito, M.; Hanaki, H. Complete genome sequence and characterization of linezolid-resistant Enterococcus faecalis clinical isolate KUB3006 carrying a cfr (B)-transposon on its chromosome and optrA-plasmid. Front. Microbiol. 2018, 9, 2576. [Google Scholar] [CrossRef] [Green Version]
- Guerin, F.; Sassi, M.; Dejoies, L.; Zouari, A.; Schutz, S.; Potrel, S.; Auzou, M.; Collet, A.; Lecointe, D.; Auger, G.; et al. Molecular and functional analysis of the novel cfr (D) linezolid resistance gene identified in Enterococcus faecium. J. Antimicrob. Chemother. 2020, 75, 1699–1703. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.K.; Fleige, C.; Klare, I.; Fiedler, S.; Mischnik, A.; Mutters, N.T.; Dingle, K.E.; Werner, G. Detection of a cfr(B) variant in German Enterococcus faecium clinical isolates and the impact on linezolid resistance in Enterococcus spp. PLoS ONE 2016, 11, e0167042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharkey, L.K.; Edwards, T.A.; O’Neill, A.J. ABC-F proteins mediate antibiotic resistance through ribosomal protection. mBio 2016, 7, e01975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Lv, Y.; Cai, J.; Schwarz, S.; Cui, L.; Hun, Z.; Zhang, R.; Li, J.; Zhao, Q.; He, T.; et al. A novel gene, optrA, that confers transferable resistance to oxazolidinones and phenicols and its presence in Enterococcus faecalis and Enterococcus faecium of human and animal origin. J. Antimicrob. Chemother. 2015, 70, 2182–2190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonelli, A.; D’Andrea, M.M.; Brenciani, A.; Galeotti, C.L.; Morroni, G.; Pollini, S.; Varaldo, P.E.; Rossolini, G.M. Characterization of poxtA, a novel phenicol-oxazolidinone-tetracycline resistance gene from an MRSA of clinical origin. J. Antimicrob. Chemother. 2018, 73, 1763–1769. [Google Scholar] [CrossRef] [Green Version]
- Papagiannitsis, C.C.; Tsilipounidaki, K.; Malli, E.; Petinaki, E. Detection in Greece of a clinical Enterococcus faecium isolate carrying the novel oxazolidinone resistance gene poxtA. J. Antimicrob. Chemother. 2019, 74, 2461–2462. [Google Scholar] [CrossRef]
- Mendes, R.E.; Deshpande, L.; Streit, J.M.; Sader, H.S.; Castanheira, M.; Hogan, P.A.; Flamm, R.K. ZAAPS programme results for 2016: An activity and spectrum analysis of linezolid using clinical isolates from medical centres in 42 countries. J. Antimicrob. Chemother. 2018, 73, 1880–1887. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Wang, Y.; Lv, Y.; Wang, S.; Song, Y.; Li, Y.; Liu, J.; Xue, F.; Yang, W.; Zhang, J. Nationwide surveillance of novel oxazolidinone resistance gene optrA in Enterococcus isolates in China from 2004 to 2014. Antimicrob. Agents Chemother. 2016, 60, 7490–7493. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, L.M.; Castanheira, M.; Flamm, R.K.; Mendes, R.E. Evolving oxazolidinone resistance mechanisms in a worldwide collection of enterococcal clinical isolates: Results from the SENTRY Antimicrobial Surveillance Program. J. Antimicrob. Chemother. 2018, 73, 2314–2322. [Google Scholar] [CrossRef] [Green Version]
- Sassi, M.; Guérin, F.; Zouari, A.; Beyrouthy, R.; Auzou, M.; Fines-Guyon, M.; Potrel, S.; Dejoies, L.; Collet, A.; Boukthir, S.; et al. Emergence of optrA-mediated linezolid resistance in enterococci from France, 2006–2016. J. Antimicrob. Chemother. 2019, 74, 1469–1472. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, CLSI Supplement M100, 28th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version, 8th ed.; The European Committee on Antimicrobial Susceptibility Testing: Växjö, Sweden, 2018. [Google Scholar]
- Hong, T.; Li, X.; Wang, J.; Sloan, C.; Cicogna, C. Sequential linezolid-resistant Staphylococcus epidermidis isolates with G2576T mutation. J. Clin. Microbiol. 2007, 45, 3277–3280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Joensen, K.G.; Scheutz, F.; Lund, O.; Hasman, H.; Kaas, R.S.; Nielsen, E.M.; Aarestrup, F.M. Real-time whole-genome sequencing for routine typing, surveillance, and outbreak detection of verotoxigenic Escherichia coli. J. Clin. Micobiol. 2014, 52, 1501–1510. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus sequence typing of total genome sequenced bacteria. J. Clin. Micobiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [Green Version]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The Reference Centre for Bacterial Insertion Sequences. Nucleic Acids Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [Green Version]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the problem of comparing whole bacterial genomes across different sequencing platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef] [Green Version]
- López, M.; Alvarez-Martínez, M.J.; Marco, F.; Torres, C. Glycopeptide-resistant Enterococcus faecium. Analysis of the resistance genotype, virulence and genetic Lines. Enferm. Infecc. Microbiol Clin. 2013, 31, 10–14. [Google Scholar] [CrossRef]
- Tenover, F.C.; Arbet, R.D.; Goering, R.V.; Mickelsen, P.A.; Murray, B.E.; Persing, D.H.; Swaminathan, B. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: Criteria for bacterial strain typing. J. Clin. Microbiol. 1995, 33, 2233–2239. [Google Scholar] [CrossRef] [Green Version]
- Dunny, G.M.; Craig, R.A.; Carron, R.L.; Clewell, D.B. Plasmid transfer in Streptococcus faecalis: Production of multiple sex pheromones by recipients. Plasmid 1979, 2, 454–465. [Google Scholar] [CrossRef] [Green Version]
- He, T.; Shen, Y.; Schwarz, S.; Cai, J.; Lv, Y.; Li, J.; Feßler, A.T.; Zhang, R.; Wu, C.; Shen, J.; et al. Genetic environment of the transferable oxazolidinone/phenicol resistance gene optrA in Enterococcus faecalis isolates of human and animal origin. J. Antimicrob. Chemother. 2016, 71, 1466–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Gómez, J.C.; Fraile-Malmierca, F.; Valverdé-Romero, E.D.; Sánchez, M.; García-Rodríguez, J.A.; García-Sánchez, J.E. Linezolid-resistant Enterococcus faecalis: First report in Spain. J. Chemother. 2006, 18, 440–442. [Google Scholar] [CrossRef]
- Gawryszewska, I.; Żabicka, D.; Hryniewicz, W.; Sadowy, E. Linezolid-resistant enterococci in Polish hospitals: Species, clonality and determinants of linezolid resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 1279–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cámara, J.; Camoez, M.; Tubau, F.; Pujol, M.; Ayats, J.; Ardanuy, C.; Domínguez, M.Á. Detection of the novel optrA gene among linezolid-resistant enterococci in Barcelona, Spain. Microb. Drug Resist. 2019, 25, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moure, Z.; Lara, N.; Marín, M.; Sola-Campoy, P.J.; Bautista, V.; Gómez-Bertomeu, F.; Gómez-Dominguez, C.; Pérez-Vázquez, M.; Aracil, B.; Campos, J.; et al. Interregional spread in Spain of linezolid-resistant Enterococcus spp. isolates carrying the optrA and poxtA genes. Int. J. Antimicrob. Agents 2020, 21, 105977. [Google Scholar] [CrossRef]
- Cai, J.; Wang, Y.; Schwarz, S.; Lv, H.; Li, Y.; Liao, K.; Yu, S.; Zhao, K.; Gu, D.; Wang, X.; et al. Enterococcal isolates carrying the novel oxazolidinone resistance gene optrA from hospitals in Zhejiang, Guangdong, and Henan, China, 2010–2014. Clin. Microbiol. Infect. 2015, 21, 1095.e1–1095.e4. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.; Huh, H.J.; Song, D.J.; Shim, H.J.; Park, K.S.; Kang, C.I.; Ki, C.S.; Lee, N.Y. Resistance mechanisms of linezolid-nonsusceptible enterococci in Korea: Low rate of 23S rRNA mutations in Enterococcus faecium. J. Med. Microbiol. 2017, 66, 1730–1735. [Google Scholar] [CrossRef]
- Chen, M.; Pan, H.; Lou, Y.; Wu, Z.; Zhang, J.; Huang, Y.; Yu, W.; Qiu, Y. Epidemiological characteristics and genetic structure of linezolid-resistant Enterococcus faecalis. Infect. Drug Resist. 2018, 11, 2397–2409. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Dong, G.; Li, J.; Chen, L.; Liu, H.; Bi, W.; Lu, H.; Zhou, T. A high incidence and coexistence of multiresistance genes cfr and optrA among linezolid-resistant enterococci isolated from a teaching hospital in Wenzhou, China. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 1441–1448. [Google Scholar] [CrossRef]
- Hua, R.; Xia, Y.; Wu, W.; Yang, M.; Yan, J. Molecular epidemiology and mechanisms of 43 low-level linezolid-resistant Enterococcus faecalis strains in Chongqing, China. Ann. Lab. Med. 2019, 39, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Schwarz, S.; Chi, D.; Wang, Z.; Zhang, R.; Wang, Y. Faecal carriage of optrA-positive enterococci in asymptomatic healthy humans in Hangzhou, China. Clin. Microbiol. Infect. 2019, 25, 630.e1–630.e6. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Fan, R.; Wang, Y.; Lei, L.; Feßler, A.T.; Wang, Z.; Wu, C.; Schwarz, S.; Wang, Y. Analysis of combined resistance to oxazolidinones and phenicols among bacteria from dogs fed with raw meat/vegetables and the respective food items. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, S.; Boan, P.; Lee, T.; Gangatharan, S.; Tan, S.J.; Daley, D.; Lee, Y.T.; Coombs, G.W. Linezolid-resistant ST872 Enteroccocus faecium harbouring optrA and cfr (D) oxazolidinone resistance genes. Int. J. Antimicrob. Agents 2020, 55, 105831. [Google Scholar] [CrossRef]
- Lee, T.; Pang, S.; Abraham, S.; Coombs, G.W. Antimicrobial-resistant CC17 Enterococcus faecium: The past, the present and the future. J. Glob. Antimicrob. Resist. 2019, 16, 36–47. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
| Isolate | Hospital 1 | Year | Type of Sample | MLST/CC | Linezolid MIC (mg/L) | Mechanisms of Linezolid Resistance 2 | Antimicrobial Resistance Phenotype 3 | Antimicrobial Resistance Genotype | Virulence Genes |
|---|---|---|---|---|---|---|---|---|---|
| C9884 | HSJ | 2017 | Urine | ST585/CC4 | 16 | optrA | ERY-CLI-GEN-KAN-STR-TET-CHL-FFC-LZD-TDZ-SXT-CIP 4-LEV | erm(B), lnu(B), lsa(A), lsa(E), aac(6′)-Ie-aph(2″)-Ia, aph(3″)-III, ant(6)-Ia, str, tet(M), tet(L), optrA, fexA, dfrG | gelE, ace, agg, efaAfs, elrA, srtA, ccf, cob, cad, camE, ebpA, ebpC, fsrB, tpx |
| C8946 | HUG | 2017 | Leg wound exudate | ST480 | 16 | optrA | ERY-CLI-GEN-KAN-STR-TET-CHL-FFC-LZD-TDZ-SXT-CIP 4,5-LEV | erm(B), lnu(B), lsa(A), lsa(E), aac(6′)-Ie-aph(2″)-Ia, aph(3″)-III, ant(6)-Ia, tet(M), tet(L), optrA, fexA, dfrG | hylA, ace, efaAfs, elrA, srtA, ccf, cob, cad, camE, ebpA, ebpB, tpx |
| C9901 | HUG | 2017 | Urine | ST21/CC21 | 16 | optrA | ERY-CLI-KAN-STR-TET-CHL-FFC-LZD-TDZ | erm(B), lnu(B), lsa(A), lsa(E), aph(3″)-III, ant(6)-Ia, tet(M), tet(L), optrA, fexA | gelE, hylA, hylB, ace, agg, cylL, efaAfs, elrA, srtA, ccf, cob, cad, camE, ebpA, ebpC, fsrB, tpx |
| C9951 | HSP | 2017 | Urine | ST16 | 8 | optrA (Y176D, T481P); 23S rDNA (G2576T) | ERY-CLI-GEN-KAN-STR-TET-CHL-FFC-LZD-TDZ-SXT | erm(A)-like, erm(B), lnu(B), lsa(A), lsa(E), aac(6′)-Ie-aph(2″)-Ia, aph(3″)-III, ant(6)-Ia, tet(M), tet(L), tet(O), optrA, fexA, dfrG | ace, agg, cylA, efaAfs, elrA, srtA, ccf, cob, cad, camE, ebpA, ebpC, tpx |
| C9952 | HSP | 2017 | Urine | ST35 | 8 | optrA (Y176D, T481P) | ERY-CLI-GEN-KAN-STR-TET CHL-FFC-LZD-TDZ-CIP 4,6-LEV | erm(A)-like, erm(B), lnu(B), lsa(A), lsa(E), aac(6′)-Ie-aph(2″)-Ia, aph(3″)-III, ant(6)-Ia, tet(M), tet(L), optrA, fexA | gelE, hylB, ace, cylA, cylL, cylM, efaAfs, elrA, srtA, ccf, cob, cad, camE, ebpA, ebpB, fsrB, tpx |
| C9736 | HVC | 2017 | Urine | ST585/CC4 | 16 | optrA | ERY-CLI-GEN-KAN-STR-TET-CHL-FFC-LZD-TDZ-SXT-CIP 4-LEV | erm(B), lnu(B), lsa(A), lsa(E), aac(6′)-Ie-aph(2″)-Ia, aph(3″)-III, ant(6)-Ia, str, tet(M), tet(L), optrA, fexA, dfrG | gelE, ace, agg, cylL, efaAfs, elrA, srtA, ccf, cob, cad, camE, ebpA, ebpC, fsrB, tpx |
| C9737 | HVC | 2017 | Urine | ST896 | 16 | optrA | ERY-CLI-KAN-STR-TET-CHL-FFC-LZD-TDZ-CIP 4,5-LEV | erm(B), lnu(B), lsa(A), lsa(E), aph(3″)-III, ant(6)-Ia, tet(M), optrA, fexA, catpC221 | esp, gelE, hylA, hylB, ace, agg, efaAfs, elrA, srtA, ccf, cob, cad, camE, ebpA, ebpB, tpx |
| X523 | HVC | 2018 | Otic exudate | ST480 | 8 | optrA (T572P) | ERY-CLI-TET-CHL-FFC-LZD-TDZ-SXT-CIP 4,5-LEV | lsa(A), lsa(E), tet(M), tet(L), optrA, fexA, dfrG | hylA, ace, efaAfs, elrA, srtA, ccf, cob, cad, camE, ebpA, ebpB, tpx |
| X524 | HVC | 2018 | Urine | ST480 | 8 | optrA | ERY-CLI-GEN-KAN-STR-TET-CHL-FFC-LZD-TDZ-SXT-CIP 4,5-LEV | erm(B), lsa(A), lsa(E), aac(6′)-Ie-aph(2″)-Ia, aph(3″)-III, ant(6)-Ia, tet(M), tet(L), optrA, fexA, dfrG | hylA, ace, agg, cylL, cylM, efaAfs, elrA, srtA, ccf, cob, cad, camE, ebpA, ebpB, tpx |
| X525 | HVC | 2018 | Ulcer | ST585/CC4 | 8 | optrA | ERY-CLI-GEN-KAN-STR-TET-CHL-FFC-LZD-TDZ-SXT-CIP 4-LEV | erm(B), lnu(B), lsa(A), lsa(E), aac(6′)-Ie-aph(2″)-Ia, aph(3″)-III, ant(6)-Ia, str, tet(M), tet(L), optrA, fexA, catA, dfrG | gelE, ace, agg, cylL, efaAfs, elrA, srtA, cob, cad, camE, ebpA, ebpC, fsrB, tpx |
| X526 | HVC | 2018 | Urine | ST896 | 4 | optrA | ERY-CLI-KAN-STR-TET-CHL-FFC-LZD-TDZ-CIP 4,5-LEV | erm(B), lnu(B), lsa(A), lsa(E), aph(3″)-III, ant(6)-Ia, tet(M), optrA, fexA, catpC221 | gelE, hylA, hylB, ace, cylA, cylL, efaAfs, elrA, srtA, ccf, cob, cad, camE, ebpA, ebpB, fsrB, tpx |
| X527 | HVC | 2018 | Ulcer | ST6/CC2 | 16 | optrA | ERY-CLI-GEN-KAN-STR-TET-CHL-FFC-LZD-TDZ-CIP 4,5-LEV | erm(B), lsa(A), aac(6′)-aph(2″), aph(3″)-III, ant(6)-Ia, tet(M), optrA, fexA | gelE, hylA, hylB, ace, agg, efaAfs, elrA, srtA, cob, cad, camE, ebpA, ebpB, ebpC, fsrB, tpx |
| X528 | HVC | 2018 | Abscess | ST480 | 16 | optrA; cfr(D) | ERY-CLI-KAN-STR-TET-CHL-FFC-LZD-TDZ-SXT-CIP 4,5-LEV | erm(B), lsa(A), aph(3′’)-III, ant(6)-Ia, tet(M), tet(L), cfr(D), optrA, fexA, dfrG | hylA, ace, agg, efaAfs, elrA, srtA, ccf, cob, cad, camE, ebpA, ebpB, tpx |
| Isolate | Hospital 1 | Year | Type of Sample | PFGE Pattern | MLST | LZD MIC (mg/L) | Mechanisms of LZD Resistance 2 | Antimicrobial Resistance Phenotype 3 | Antimicrobial Resistance Genotype 4 | Virulence Genes |
|---|---|---|---|---|---|---|---|---|---|---|
| C9902 | HLB | 2017 | Abscess | A | ST17 | 8 | 23S rDNA (G2576T) | PEN 5-AMP-ERY-CLI-KAN-STR-CIP 8-LEV-LZD-SXT | erm(B), msrC, aac(6′)-Ii, aph(3″)-III, ant(6)-Ia, dfrG | esp, hyl |
| C9903 | HLB | 2017 | Surgical wound | B | ST262 | 16 | 23S rDNA (G2576T) | PEN 5-AMP-ERY-CLI-GEN-KAN-STR-TET-CIP 8-LEV-LZD-SXT | msr(C), lsa(E), aac(6′)-Ie-aph(2″)-Ia, aac(6′)-Ii, ant(6)-Ia, tet(M), tet(L), dfrG | |
| C10262 | HLB | 2017 | Blood | C | ST117 | 32 | 23S rDNA (G2576T) | PEN 6-AMP-ERY-CLI-KAN-STR-CIP 8-LEV-LZD-SXT | erm(B), msr(C), aac(6′)-Ii, aph(3″)-III, ant(6)-Ia, dfrG | esp, hyl |
| C9950 | HSP | 2017 | Blood | D | ST117 | 4 | 23S rDNA (G2576T) | PEN 7-AMP-ERY-CLI-KAN-STR-CIP 8-LEV-LZD-SXT | erm(B), msr(C), aac(6′)-Ii, aph(3″)-III, ant(6)-Ia, dfrG | esp, hyl |
| C9953 | HSP | 2017 | Urine | E | ST117 | 16 | 23S rDNA (G2576T) | PEN I,5-AMP-ERY-CLI-KAN-STR-CIP 8-LEV-LZD-SXT | erm(B), msr(C), aac(6′)-Ii, aph(3″)-III, ant(6)-Ia, dfrG | hyl |
| C10269 | HRV | 2017 | Surgical wound | F | ST117 | 16 | 23S rDNA (G2576T) | PEN 6-AMP-ERY-CLI-KAN-STR-CIP 8-LEV-LZD-SXT | erm(B), msr(C), aac(6′)-Ii, aph(3″)-III, ant(6)-Ia, dfrG | esp, hyl |
| Strain | Genotype | MIC (mg/L) 1 | ||||||
|---|---|---|---|---|---|---|---|---|
| ERY | CLI | TET | CHL | FFC | LZD | TDZ | ||
| E. faecalisJH2-2 (recipient) | - | 1 | 2 | 0.125 | 4 | 4 | 2 | 1 |
| E. faecalisC9884 | erm(B), lnu(B), lsa(A), lsa(E), aac(6′)-aph(2″), aph(3″)-III, ant(6)-Ia, str, tet(M), tet(L), optrA, fexA, dfrG | >256 | 256 | 128 | 64 | 64 | 16 | 1 |
| E. faecalisTC-C9884 | optrA, fexA | 1 | 2 | 0.125 | 32 | 64 | 8 | 1 |
| E. faecalisC9901 | erm(B), lnu(B), lsa(A), lsa(E), aph(3″)-III, ant(6)-Ia, tet(M), tet(L), optrA, fexA | >256 | >256 | >256 | 32 | 64 | 16 | 1 |
| E. faecalisTC-C9901 | optrA, fexA | 1 | 2 | 0.125 | 32 | 64 | 4 | 1 |
| E. faecalisC9951 | erm(A)-like, erm(B), lnu(B), lsa(A), lsa(E), aac(6′)-aph(2″), aph(3″)-III, ant(6)-Ia, tet(M), tet(L), tet(O), optrA, fexA, dfrG | >256 | >256 | 128 | 64 | 64 | 8 | 1 |
| E. faecalisTC-C9951 | erm(A)-like, tet(L), tet(O), optrA, fexA | 256 | >256 | 128 | 64 | 64 | 4 | 1 |
| E. faecalisC9952 | erm(A)-like, erm(B), lnu(B), lsa(A), lsa(E), aac(6′)-aph(2″), aph(3″)-III, ant(6)-Ia, tet(M), tet(L), optrA, fexA | 256 | >256 | >256 | 32 | 64 | 8 | 1 |
| E. faecalisTC-C9952 | erm(A)-like, optrA, fexA | 128 | >256 | 0.125 | 32 | 64 | 4 | 1 |
| E. faecalisC9736 | erm(B), lnu(B), lsa(A), lsa(E), aac(6′)-aph(2″), aph(3″)-III, ant(6)-Ia, str, tet(M), tet(L), optrA, fexA, dfrG | >256 | 128 | 256 | 128 | 32 | 16 | 1 |
| E. faecalisTC-C9736 | optrA, fexA | 1 | 2 | 0.125 | 64 | 32 | 8 | 1 |
| E. faecalisC9737 | erm(B), lnu(B), lsa(A), lsa(E), aph(3″)-III, ant(6)-Ia, tet(M), optrA, fexA, catpC221 | >256 | 128 | 128 | 32 | 64 | 16 | 1 |
| E. faecalisTC-C9737 | optrA, fexA | 1 | 2 | 0.125 | 32 | 32 | 8 | 1 |
| E. faecalisX523 | lsa(A), lsa(E), tet(M), tet(L), optrA, fexA, dfrG | 128 | >256 | 128 | 32 | 64 | 8 | 1 |
| E. faecalisTC-X523 | optrA, fexA | 1 | 2 | 0.125 | 32 | 64 | 8 | 1 |
| E. faecalisX524 | erm(B), lsa(A), lsa(E), aac(6′)-aph(2″), aph(3″)-III, ant(6)-Ia, tet(M), tet(L), optrA, fexA, dfrG | 256 | 256 | 256 | 32 | 64 | 8 | 1 |
| E. faecalisTC-X524 | optrA, fexA | 1 | 2 | 0.125 | 32 | 64 | 8 | 1 |
| E. faecalisX525 | erm(B), lnu(B), lsa(A), lsa(E), aac(6′)-aph(2″), aph(3″)-III, ant(6)-Ia, str, tet(M), tet(L), optrA, fexA, catA, dfrG | >256 | 128 | 128 | 64 | 64 | 8 | 1 |
| E. faecalisTC-X525 | optrA, fexA | 1 | 2 | 0.125 | 32 | 32 | 8 | 1 |
| E. faecalisX526 | erm(B), lnu(B), lsa(A), lsa(E), aph(3″)-III, ant(6)-Ia, tet(M), optrA, fexA, catpC221 | >256 | 128 | 128 | 32 | 64 | 4 | 1 |
| E. faecalisTC-X526 | optrA, fexA | 1 | 2 | 0.125 | 32 | 64 | 4 | 1 |
| E. faecalisX527 | erm(B), lsa(A), aac(6′)-aph(2″), aph(3″)-III, ant(6)-Ia, tet(M), optrA, fexA | >256 | 256 | 128 | 32 | 64 | 16 | 1 |
| E. faecalisTC-X527 | optrA, fexA | 1 | 2 | 0.125 | 32 | 64 | 8 | 1 |
| E. faecalisX528 | erm(B), lsa(A), aph(3″)-III, ant(6)-Ia, tet(M), tet(L), cfr(D), optrA, fexA, dfrG | >256 | >256 | >256 | 32 | 64 | 16 | 1 |
| E. faecalisTC-X528 | optrA, fexA | 1 | 2 | 0.125 | 32 | 64 | 8 | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruiz-Ripa, L.; Feßler, A.T.; Hanke, D.; Eichhorn, I.; Azcona-Gutiérrez, J.M.; Pérez-Moreno, M.O.; Seral, C.; Aspiroz, C.; Alonso, C.A.; Torres, L.; et al. Mechanisms of Linezolid Resistance Among Enterococci of Clinical Origin in Spain—Detection of optrA- and cfr(D)-Carrying E. faecalis. Microorganisms 2020, 8, 1155. https://doi.org/10.3390/microorganisms8081155
Ruiz-Ripa L, Feßler AT, Hanke D, Eichhorn I, Azcona-Gutiérrez JM, Pérez-Moreno MO, Seral C, Aspiroz C, Alonso CA, Torres L, et al. Mechanisms of Linezolid Resistance Among Enterococci of Clinical Origin in Spain—Detection of optrA- and cfr(D)-Carrying E. faecalis. Microorganisms. 2020; 8(8):1155. https://doi.org/10.3390/microorganisms8081155
Chicago/Turabian StyleRuiz-Ripa, Laura, Andrea T. Feßler, Dennis Hanke, Inga Eichhorn, José Manuel Azcona-Gutiérrez, Mar Olga Pérez-Moreno, Cristina Seral, Carmen Aspiroz, Carla Andrea Alonso, Luis Torres, and et al. 2020. "Mechanisms of Linezolid Resistance Among Enterococci of Clinical Origin in Spain—Detection of optrA- and cfr(D)-Carrying E. faecalis" Microorganisms 8, no. 8: 1155. https://doi.org/10.3390/microorganisms8081155
APA StyleRuiz-Ripa, L., Feßler, A. T., Hanke, D., Eichhorn, I., Azcona-Gutiérrez, J. M., Pérez-Moreno, M. O., Seral, C., Aspiroz, C., Alonso, C. A., Torres, L., Alós, J.-I., Schwarz, S., & Torres, C. (2020). Mechanisms of Linezolid Resistance Among Enterococci of Clinical Origin in Spain—Detection of optrA- and cfr(D)-Carrying E. faecalis. Microorganisms, 8(8), 1155. https://doi.org/10.3390/microorganisms8081155