Biocontrol and Plant-Growth-Promoting Traits of Talaromyces apiculatus and Clonostachys rosea Consortium against Ganoderma Basal Stem Rot Disease of Oil Palm

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolation and Maintenance
2.2. Molecular and Phylogenetic Analyses of AAT0115 and AAB0114
2.3. Dual-Culture Bioassays
2.4. Plant-Growth-Promoting Analysis and Determination of Leaf Chlorophyll Contents
2.5. Plant Nutrient Analyses
2.6. Nursery Biocontrol Experiment
2.7. Statistical Analyses
3. Results
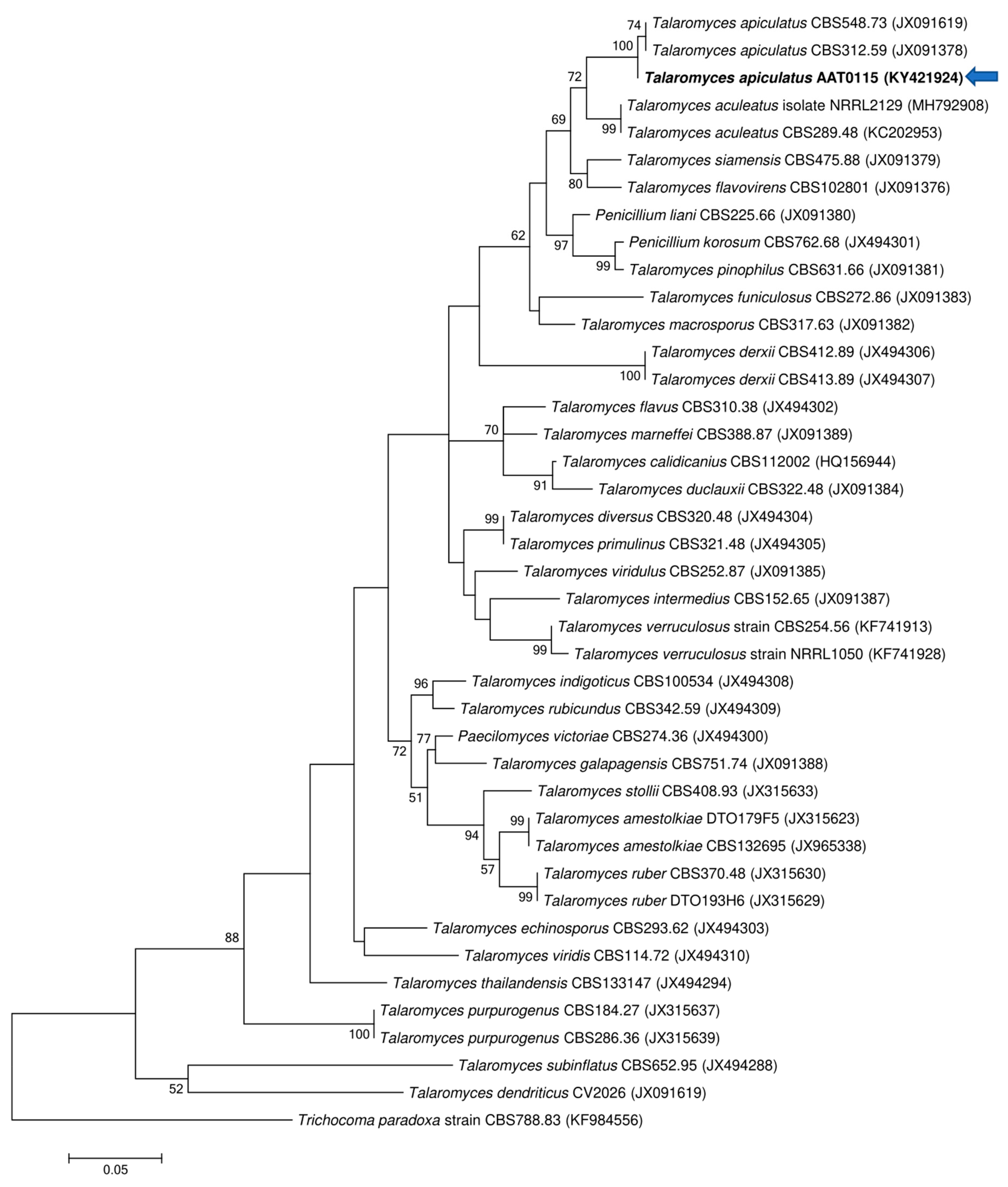
3.1. Molecular Identification of AAT0115 and AAB0114 Isolates
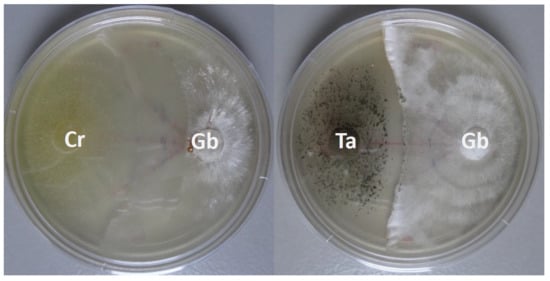
3.2. In Vitro Ganoderma Boninense Growth Suppression
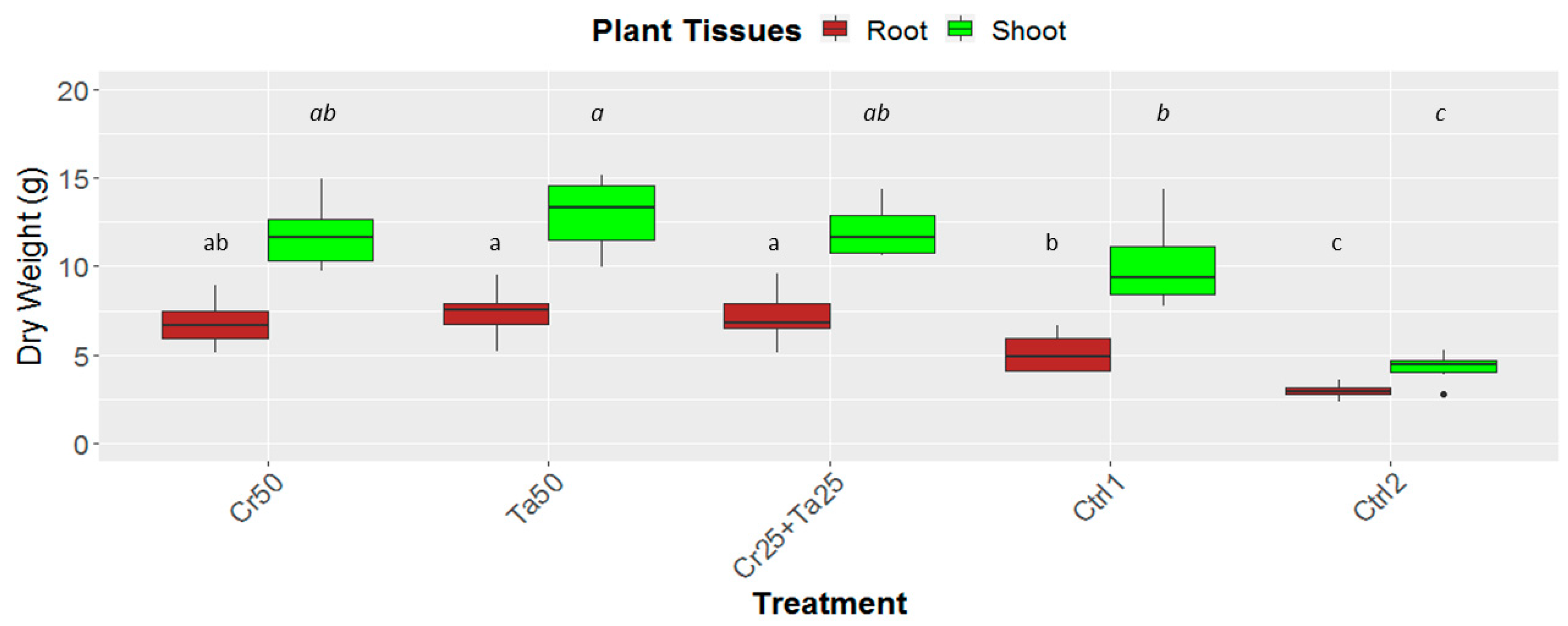
3.3. Plant-Growth-Promoting Abilities of Clonostachys rosea and Talaromyces apiculatus
3.4. Efficacy of Clonostachys rosea and Talaromyces apiculatus in controlling Ganoderma Basal Stem Rot Disease of Oil Palm
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Idris, A.; Kushairi, A.; Ismail, S.; Ariffin, D. Selection for partial resistance in oil palm progenies to Ganoderma basal stem rot. J. Oil Palm Res. 2004, 16, 12–18. [Google Scholar]
- Hushiarian, R.; Yusof, N.A.; Dutse, S.W. Detection and control of Ganoderma boninense: Strategies and perspectives. Springerplus 2013, 2, 555. [Google Scholar] [CrossRef] [PubMed]
- Roslan, A.; Idris, A.S. Economic impact of Ganoderma incidence on Malaysian oil palm plantation—A case study in Johor. Oil Palm Ind. Econ. J. 2012, 12, 24–30. [Google Scholar]
- Cooper, R.M.; Flood, J.; Rees, R.W. Ganoderma boninense in oil palm plantations: Current thinking on epidemiology, resistance and pathology. Planter 2011, 87, 515–526. [Google Scholar]
- Singh, G. Ganoderma-the scourge of oil palms in the coastal areas. Planter 1991, 67, 421–444. [Google Scholar]
- Turner, P.D. Oil Palm Diseases and Disorders; Oxford University Press: Oxford, UK, 1981. [Google Scholar]
- Idris, A. Basal Stem Rot (BSR) of Oil Palm (Elaeis Guineensis Jacq.) in Malaysia: Factors Associated with Variation in Disease Severity; Wye College, University of London: Wye, UK, 1999. [Google Scholar]
- Chung, G.F. Management of Ganoderma diseases in oil palm plantations. Planter 2011, 87, 325–339. [Google Scholar]
- Naher, L.; Yusuf, U.K.; Ismail, A.; Tan, S.G.; Mondal, M.M.A. Ecological status of Ganoderma and basal stem rot disease of oil palms (Elaeis guineensis Jacq.). Aust. J. Crop Sci. 2013, 7, 1723–1727. [Google Scholar]
- Saldajeno, M.G.B.; Ito, M.; Hyakumachi, M. Interaction between the plant growth-promoting fungus Phoma sp. GS8-2 and the arbuscular mycorrhizal fungus Glomus mosseae: Impact on biocontrol of soil-borne diseases, microbial population, and plant growth. Australas. Plant Pathol. 2012, 41, 271–281. [Google Scholar] [CrossRef]
- Fravel, D.R. Commercialization and implementation of biocontrol. Annu. Rev. Phytopathol. 2005, 43, 337–359. [Google Scholar] [CrossRef]
- Guetsky, R.; Shtienberg, D.; Elad, Y.; Dinoor, A. Combining biocontrol agents to reduce the variability of biological control. Phytopathology 2001, 91, 621–627. [Google Scholar] [CrossRef]
- Haggag, W.M. Improving the biological control of Botryodiplodia disease on some Annona cultivars using single or multi-bioagents in Egypt. Biol. Control 2006, 38, 341–349. [Google Scholar] [CrossRef]
- Roberts, D.P.; Lohrke, S.M.; Meyer, S.L.F.; Buyer, J.S.; Bowers, J.H.; Jacyn Baker, C.; Li, W.; de Souza, J.T.; Lewis, J.A.; Chung, S. Biocontrol agents applied individually and in combination for suppression of soilborne diseases of cucumber. Crop Prot. 2005, 24, 141–155. [Google Scholar] [CrossRef]
- Elead, Y.; Köhl, J.; Fokkema, N.J. Control of infection and sporulation of Botrytis cinerea on bean and tomato by saprophytic bacteria and fungi. Eur. J. Plant Pathol. 1994, 100, 315–336. [Google Scholar] [CrossRef]
- Fukui, R.; Fukui, H.; Alvarez, A.M. Comparisons of single versus multiple bacterial species on biological control of Anthurium blight. Phytopathology 1999, 89, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Guetsky, R.; Shtienberg, D.; Elad, Y.; Fischer, E.; Dinoor, A. Improving biological control by combining biocontrol agents each with several mechanisms of disease suppression. Phytopathology 2002, 92, 976–985. [Google Scholar] [CrossRef] [PubMed]
- Guetskyl, R.; Shtienberg, D.; Dinoor, A.; Elad, Y. Establishment, survival and activity of the biocontrol agents Pichia guilermondii and Bacillus mycoides applied as a mixture on strawberry plants. Biocontrol Sci. Technol. 2002, 12, 705–714. [Google Scholar] [CrossRef]
- Nusaibah, S.A.; Saad, G.; Tan, G.H. Antagonistic Efficacy of Trichoderma harzianum and Bacillus cereus against Ganoderma Disease of Oil Palm via Dip, Place and Drench (DPD) Artificial Inoculation Technique. Int. J. Agric. Biol. 2017, 19, 299–306. [Google Scholar] [CrossRef]
- Sundram, S.; Meon, S.; Seman, I.A.; Othman, R. Symbiotic interaction of endophytic bacteria with arbuscular mycorrhizal fungi and its antagonistic effect on Ganoderma boninense. J. Microbiol. 2011, 49, 551. [Google Scholar] [CrossRef]
- Sundram, S.; Meon, S.; Seman, I.A.; Othman, R. Application of arbuscular mycorrhizal fungi with Pseudomonas aeruginosa UPMP3 reduces the development of Ganoderma basal stem rot disease in oil palm seedlings. Mycorrhiza 2015, 25, 387–397. [Google Scholar] [CrossRef]
- Wan Zakaria, W.N.F. Screening Potential Biological Control Agents and Efficacy of Mixed-Microorganisms in Controlling Oil Palm Basal Stem Rot Caused by Ganoderma boninense. Master’s Thesis, Universiti Malaysia Sarawak (UNIMAS), Kota Samarahan, Malaysia, 2014. [Google Scholar]
- Kirschner, R.; Arnold, G.R.W.; Chen, C.-J. Cladobotryum semicirculare sp. nov. (hyphomycetes) from commercially grown Ganoderma tsugae in Taiwan and other Basidiomycota in Cuba. Sydowia 2007, 59, 114–124. [Google Scholar]
- Marzuki, N.F.; Goh, Y.K.; Tung, H.J.; Goh, Y.K.; Goh, K.J. Evaluation on the cultural characteristics and antagonistic activities of Cladobotryum semicirculare against Ganoderma boninense in vitro. J. Oil Palm Res. 2015, 27, 326–338. [Google Scholar]
- Kang, H.-J.; Sigler, L.; Lee, J.; Gibas, C.F.C.; Yun, S.-H.; Lee, Y.-W. Xylogone ganodermophthora sp. nov., an ascomycetous pathogen causing yellow rot on cultivated mushroom Ganoderma lucidum in Korea. Mycologia 2010, 102, 1167–1184. [Google Scholar] [CrossRef] [PubMed]
- Goh, Y.K.; Goh, T.K.; Marzuki, N.F.; Tung, H.J.; Goh, Y.K.; Goh, K.J. Scytalidium parasiticum sp. nov., a new species parasitizing on Ganoderma boninense isolated from oil palm in Peninsular Malaysia. Mycobiology 2015, 43, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Agustini, L.; Wahyuno, D.; Indrayadi, H.; Glen, M. In vitro interaction between Phlebiopsis sp. and Ganoderma philippii isolates. Forest Pathol. 2014, 44, 472–476. [Google Scholar] [CrossRef]
- Helfer, W. Pilze auf Pilzfruchtkӧrpern. Libri Botanici 1; IHW-Verlag: Eching, Germany, 1991. [Google Scholar]
- Goh, Y.K.; Marzuki, N.F.; Goh, T.K.; Tan, S.Y.; Goh, Y.K.; Goh, K.J. Mycoparasitic Scytalidium parasiticum as a potential biocontrol agent against Ganoderma boninense basal stem rot in oil palm. Biocontrol Sci. Technol. 2016, 26, 1352–1365. [Google Scholar] [CrossRef]
- Gohel, V.; Singh, A.; Vimal, M.; Ashwini, P.; Chhatpar, H.S. Bioprospecting and Antifungl Potential of Chitinolytic Microorganisms. Afr. J. Biotechnol. 2006, 5, 54–72. [Google Scholar]
- Naraghi, L.; Heydari, A.; Rezaee, S.; Razavi, M. Biocontrol Agent Talaromyces flavus Stimulates the Growth of Cotton and Potato. J. Plant Growth Regul. 2012, 31, 471–477. [Google Scholar] [CrossRef]
- Kok, S.M.; Goh, Y.K.; Tung, H.J.; Goh, K.J.; Wong, W.C.; Goh, Y.K. In vitro growth of Ganoderma boninense isolates on novel palm extract medium and virulence on oil palm (Elaeis guineensis) seedlings. Malays. J. Microbiol. 2013, 9, 33–42. [Google Scholar]
- Ariffin, D.; Idris, A. A selective medium for the isolation of Ganoderma from diseased tissues. In Proceedings of Proceedings of the International Palm Oil Conference, Progress, Prospects and Challenges Towards the 21st Century (Model 1—Agriculture), Kuala Lumpur Hilton, Malaysia, 9–14 September 1991; Palm Oil Research Institute of Malaysia: Bangi, Malaysia, 1991; pp. 517–519. [Google Scholar]
- Rees, R.W.; Flood, J.; Hasan, Y.; Cooper, R.M. Effects of inoculum potential, shading and soil temperature on root infection of oil palm seedlings by the basal stem rot pathogen Ganoderma boninense. Plant Pathol. 2007, 56, 862–870. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.B.; Taylor, J.W. Analysis of phylogenetic relationships by amplification and direct sequencing of ribosomal DNA genes. In PCR Protocols: A guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Vujanovic, V.; Goh, Y.K. Sphaerodes mycoparasitica sp. nov., a new biotrophic mycoparasite on Fusarium avenaceum, F. graminearum and F. oxysporum. Mycol. Res. 2009, 113, 1172–1180. [Google Scholar] [CrossRef]
- Sokolski, S.; Piché, Y.; Bérubé, J.A. Lophodermium macci sp. nov., a new species on senesced foliage of five-needle pines. Mycologia 2004, 96, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz, N.; Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of the genus Talaromyces. Stud. Mycol. 2014, 78, 175–341. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Corley, R.H.V.; Hardon, J.J.; Tan, G.Y. Analysis of growth of the oil palm (Elaeis guineensis Jacq.). Euphytica 1971, 20, 307–315. [Google Scholar] [CrossRef]
- Corley, R.H.V.; Tinker, P.B. The Oil Palm, 5th ed.; Blackwell Publishing: Oxford, UK, 2016. [Google Scholar]
- Hardon, J.J.; Williams, C.; Watson, I. Leaf area and yield in the oil palm in Malaya. Exp. Agric. 1969, 5, 25–32. [Google Scholar] [CrossRef]
- Goh, Y.K.; Ng, F.W.; Kok, S.M.; Goh, Y.K.; Goh, K.J. Aggressive of Ganoderma boninense isolates on the vegetative growth of oil palm (Elaeis guineensis) seedling at different age. Malays. J. Appl. Biol. 2014, 43, 9–16. [Google Scholar]
- Sim, C.C.; Zaharah, A.R.; Tan, M.S.; Goh, K.J. Rapid Determination of Leaf Chlorophyll Concentration, Photosynthetic Activity and NK Concentration of Elaies guineensis Via Correlated SPAD-502 Chlorophyll Index. Asian J. Agric. Res. 2015, 9, 132–138. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids: Measurement and Characterization by UV-VIS Spectroscopy. Curr. Protoc. Food Anal. Chem. 2001, 1, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Sharifuddin, H.A.H.; Fauziah, I.; Zaharah, A.R. Technique of soil testing and plant analysis and their utilization for crop production in malaysia. Commun. Soil Sci. Plant Anal. 1990, 21, 1959–1978. [Google Scholar] [CrossRef]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemiology; John Wiley & Sons: New York, NY, USA, 1990; p. 532. [Google Scholar]
- Sapak, Z.; Meon, S.; Ahmad, Z.M.A. Effect of endophytic bacteria on growth and suppression of Ganoderma infected in oil palm. Int. J. Agric. Biol. 2008, 10, 127–132. [Google Scholar]
- Simko, I.; Piepho, H.P. The area under the disease progress stairs: Calculation, advantage, and application. Phytopathology 2012, 102, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Wickham, H.; Wickham, M.H. The Ggplot Package. 2007. Available online: https://cran.r-project.org/web/packages/ggplot2/index.html (accessed on 15 March 2020).
- RStudio Team. RStudio: Integrated Development for R. Available online: http://www.rstudio.com/ (accessed on 15 March 2020).
- Schroers, H.-J.; Samuels, G.J.; Seifert, K.A.; Gams, W. Classification of the mycoparasite Gliocladium roseum in Clonostachys as C. rosea, its relationship to Bionectria ochroleuca, and notes on other Gliocladium-like fungi. Mycologia 1999, 91, 365–385. [Google Scholar] [CrossRef]
- El-Debaiky, S.A. Antagonistic studies and hyphal interactions of the new antagonist Aspergillus piperis against some phytopathogenic fungi in vitro in comparison with Trichoderma harzianum. Microb. Pathog. 2017, 113, 135–143. [Google Scholar] [CrossRef]
- Yang, D.; Plante, F.; Bernier, L.; Piché, Y.; Dessureault, M.; Laflamme, G.; Ouellette, G.B. Evaluation of a fungal antagonist, Phaeotheca dimorphospora, for biological control of tree diseases. Can. J. Bot. 1993, 71, 426–433. [Google Scholar] [CrossRef]
- Zalila-Kolsi, I.; Ben Mahmoud, A.; Ali, H.; Sellami, S.; Nasfi, Z.; Tounsi, S.; Jamoussi, K. Antagonist effects of Bacillus spp. strains against Fusarium graminearum for protection of durum wheat (Triticum turgidum L. subsp. durum). Microbiol. Res. 2016, 192, 148–158. [Google Scholar] [CrossRef]
- Costa, L.B.; Rangel, D.E.N.; Morandi, M.A.B.; Bettiol, W. Effects of UV-B radiation on the antagonistic ability of Clonostachys rosea to Botrytis cinerea on strawberry leaves. Biol. Control 2013, 65, 95–100. [Google Scholar] [CrossRef]
- Fatema, U.; Broberg, A.; Jensen, D.F.; Karlsson, M.; Dubey, M. Functional analysis of polyketide synthase genes in the biocontrol fungus Clonostachys rosea. Sci. Rep. 2018, 8, 15009. [Google Scholar] [CrossRef]
- Mamarabadi, M.; Jensen, D.F.; Lübeck, M. An N-acetyl-beta-D-glucosaminidase gene, cr-nag1, from the biocontrol agent Clonostachys rosea is up-regulated in antagonistic interactions with Fusarium culmorum. Mycol. Res. 2009, 113, 33–43. [Google Scholar] [CrossRef]
- Goh, Y.K.; Vujanovic, V. Biotrophic mycoparasitic interactions between Sphaerodes mycoparasitica and phytopathogenic Fusarium species. Biocontrol Sci. Technol. 2010, 20, 891–902. [Google Scholar] [CrossRef]
- Kim, S.H.; Vujanovic, V. Changes in mycoparasite-Fusarium hosts interfaces in response to hostile environment as revealed by water contact angle and atomic force microscopy. Biol. Control 2018, 121, 247–255. [Google Scholar] [CrossRef]
- Manoch, L.; Dethoup, T. A potential use of Talaromyces species as biological agents against plant pathogenic fungi. Thai J. Agric. Sci. 2011, 44, 81–91. [Google Scholar]
- Gao, H.; Zhou, L.; Li, D.; Gu, Q.; Zhu, T.-J. New cytotoxic metabolites from the marine-derived fungus Penicillium sp. ZLN29. Helv. Chim. Acta 2013, 96, 514–519. [Google Scholar] [CrossRef]
- Schlingmann, G.; Milne, L.; Carter, G.T. Isolation and identification of antifungal polyesters from the marine fungus Hypoxylon oceanicum LL-15G256. Tetrahedron 2002, 58, 6825–6835. [Google Scholar] [CrossRef]
- Breinholt, J.; Jensen, G.W.; Nielsen, R.I.; Olsen, C.E.; Frisvad, J.C. Antifungal macrocyclic polylactones from Penicillium verruculosum. J. Antibiot. 1993, 46, 1101–1108. [Google Scholar] [CrossRef]
- Yamagiwa, Y.; Inagaki, Y.; Ichinose, Y.; Toyoda, K.; Hyakumachi, M.; Shiraishi, T. Talaromyces wortmannii FS2 emits β-caryphyllene, which promotes plant growth and induces resistance. J. Gen. Plant Pathol. 2011, 77, 336–341. [Google Scholar] [CrossRef]
- Khalmuratova, I.; Kim, H.; Nam, Y.-J.; Oh, Y.; Jeong, M.-J.; Choi, H.-R.; You, Y.-H.; Choo, Y.-S.; Lee, I.-J.; Shin, J.-H.; et al. Diversity and Plant Growth Promoting Capacity of Endophytic Fungi Associated with Halophytic Plants from the West Coast of Korea. Mycobiology 2015, 43, 373–383. [Google Scholar] [CrossRef]
- Johansen, A.; Knudsen, I.M.B.; Binnerup, S.J.; Winding, A.; Johansen, J.E.; Jensen, L.E.; Andersen, K.S.; Svenning, M.M.; Bonde, T.A. Non-target effects of the microbial control agents Pseudomonas fluorescens DR54 and Clonostachys rosea IK726 in soils cropped with barley followed by sugar beet: A greenhouse assessment. Soil Biol. Biochem. 2005, 37, 2225–2239. [Google Scholar] [CrossRef]
- Sutton, J.C.; Liu, W.; Ma, J.; Brown, W.G.; Stewart, J.F.; Walker, G.D. Evaluation of the Fungal Endophyte Clonostachys Rosea as an Inoculant to Enhance Growth, Fitness and Productivity of Crop Plants. Acta Hortic. 2008, 782, 279–286. [Google Scholar] [CrossRef]
- Maity, A.; Pal, R.K.; Chandra, R.; Singh, N.V. Penicillium pinophilum—A novel microorganism for nutrient management in pomegranate (Punica granatum L.). Sci. Hortic. 2014, 169, 111–117. [Google Scholar] [CrossRef]
- Silitonga, N.; Sembiring, M.; Marbun, P.; Rosneli. Application of phosphate solubilizing fungi and various sources of P-Fertilizers toward P-Available and P Nutrient content of soybean (Glycine max L. Merrill) in andisol soil. IOP Conf. Ser. Earth Environ. Sci. 2019, 260, 012159. [Google Scholar] [CrossRef]
- Kamou, N.N.; Cazorla, F.; Kandylas, G.; Lagopodi, A.L. Induction of defense-related genes in tomato plants after treatments with the biocontrol agents Pseudomonas chlororaphis ToZa7 and Clonostachys rosea IK726. Arch. Microbiol. 2020, 202, 257–267. [Google Scholar] [CrossRef] [PubMed]
- Mouekouba, L.D.O.; Zhang, L.; Guan, X.; Chen, X.; Chen, H.; Zhang, J.; Zhang, J.; Li, J.; Yang, Y.; Wang, A. Analysis of Clonostachys rosea-Induced Resistance to Tomato Gray Mold Disease in Tomato Leaves. PLoS ONE 2014, 9, e102690. [Google Scholar] [CrossRef] [PubMed]
- de Boer, M.; Bom, P.; Kindt, F.; Keurentjes, J.J.B.; van der Sluis, I.; van Loon, L.C.; Bakker, P.A.H.M. Control of Fusarium wilt of radish by combining Pseudomonas putida strains that have different disease-suppressive mechanisms. Phytopathology 2003, 93, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Döring, T.F.; Pautasso, M.; Finckh, M.R.; Wolfe, M.S. Concepts of plant health—Reviewing and challenging the foundations of plant protection. Plant Pathol. 2012, 61, 1–15. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Descriptions |
|---|---|
| Cr50 | Inoculation with 50 g of blended AAB0114 (Cr) inoculant |
| Ta50 | Inoculation with 50 g of blended AAT0115 (Ta) inoculant |
| Cr25+Ta25 | Inoculation with consortium of 25 g Cr and 25 g Ta inoculant |
| Ctrl 1 (control 1) | Application of uninoculated blended maize (50 g) only |
| Ctrl 2 (control 2) | Without uninoculated blended maize |
| Ganoderma boninense Isolates | Pathogenicity Level § | Treatment | Linear Mycelial Growth of Ganoderma Colony (mm) a | |
|---|---|---|---|---|
| Ta | Cr | |||
| G. boniense isolate G8 | Low | control | 44.17 (1.77) c | 44.17 (1.77) c |
| With Ta or Cr | 20.47 (1.57) e | 22.73 (1.10) e | ||
| G. boninense isolate G10 | High | Control | 53.30 (0.25) a | 53.30 (0.25) a |
| With Ta or Cr | 26.53 (0.20) d | 32.47 (0.87) d | ||
| G. boninense isolate G14 | High | Control | 53.13 (0.33) a | 53.13 (0.33) a |
| With Ta or Cr | 25.47 (2.01) d | 31.07 (2.11) d | ||
| G. boninense isolate G12 | Moderate | Control | 48.03 (1.36) b | 48.03 (1.36) b |
| With Ta or Cr | 25.60 (0.84) d | 24.73 (2.08) e | ||
| Treatment a | Disease Census | |||||||
|---|---|---|---|---|---|---|---|---|
| DI (%) on MPI b | DSI (%) c on MPI b | AUDPC (%) d | DR (%) e | |||||
| 3 | 4 | 5 | 3 | 4 | 5 | |||
| +G10–Cr–Ta | 40 | 70 | 100 | 16.67 (7.86) a | 40.00 (10.90) a | 75.00 (7.14) a | 54.17 a | – |
| +G10+Cr50 | 10 | 50 | 80 | 5.00 (5.00) ab | 36.67 (14.00) a | 61.67 (13.20) ab | 35.84 ab | 33.84 |
| +G10+Ta50 | 30 | 60 | 80 | 16.67 (9.30) ab | 35.00 (12.50) a | 70.00 (11.90) a | 51.67 a | 4.62 |
| +G10 + Cr25 + Ta25 | 0 | 20 | 60 | 0 b | 11.67 (8.62) a | 51.67 (14.20) ab | 25.8 ab | 52.37 |
| +G10 + Cr50 + Ta50 | 10 | 20 | 50 | 5.00 (5.00) ab | 18.33 (12.30) a | 33.33 (12.90) b | 21.67 b | 60 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Goh, Y.K.; Marzuki, N.F.; Tuan Pa, T.N.F.; Goh, T.-K.; Kee, Z.S.; Goh, Y.K.; Yusof, M.T.; Vujanovic, V.; Goh, K.J. Biocontrol and Plant-Growth-Promoting Traits of Talaromyces apiculatus and Clonostachys rosea Consortium against Ganoderma Basal Stem Rot Disease of Oil Palm. Microorganisms 2020, 8, 1138. https://doi.org/10.3390/microorganisms8081138
Goh YK, Marzuki NF, Tuan Pa TNF, Goh T-K, Kee ZS, Goh YK, Yusof MT, Vujanovic V, Goh KJ. Biocontrol and Plant-Growth-Promoting Traits of Talaromyces apiculatus and Clonostachys rosea Consortium against Ganoderma Basal Stem Rot Disease of Oil Palm. Microorganisms. 2020; 8(8):1138. https://doi.org/10.3390/microorganisms8081138
Chicago/Turabian StyleGoh, Yit Kheng, Nurul Fadhilah Marzuki, Tuan Nur Fatihah Tuan Pa, Teik-Khiang Goh, Zeng Seng Kee, You Keng Goh, Mohd Termizi Yusof, Vladimir Vujanovic, and Kah Joo Goh. 2020. "Biocontrol and Plant-Growth-Promoting Traits of Talaromyces apiculatus and Clonostachys rosea Consortium against Ganoderma Basal Stem Rot Disease of Oil Palm" Microorganisms 8, no. 8: 1138. https://doi.org/10.3390/microorganisms8081138
APA StyleGoh, Y. K., Marzuki, N. F., Tuan Pa, T. N. F., Goh, T.-K., Kee, Z. S., Goh, Y. K., Yusof, M. T., Vujanovic, V., & Goh, K. J. (2020). Biocontrol and Plant-Growth-Promoting Traits of Talaromyces apiculatus and Clonostachys rosea Consortium against Ganoderma Basal Stem Rot Disease of Oil Palm. Microorganisms, 8(8), 1138. https://doi.org/10.3390/microorganisms8081138