Transferable Extended-Spectrum β-Lactamase (ESBL) Plasmids in Enterobacteriaceae from Irrigation Water

Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. MALDI Biotyping
2.3. Antibiotic Susceptibility Testing
2.4. DNA Extractions, Sequencing, and Bioinformatics
2.5. Plasmid Transfer Assays
3. Results
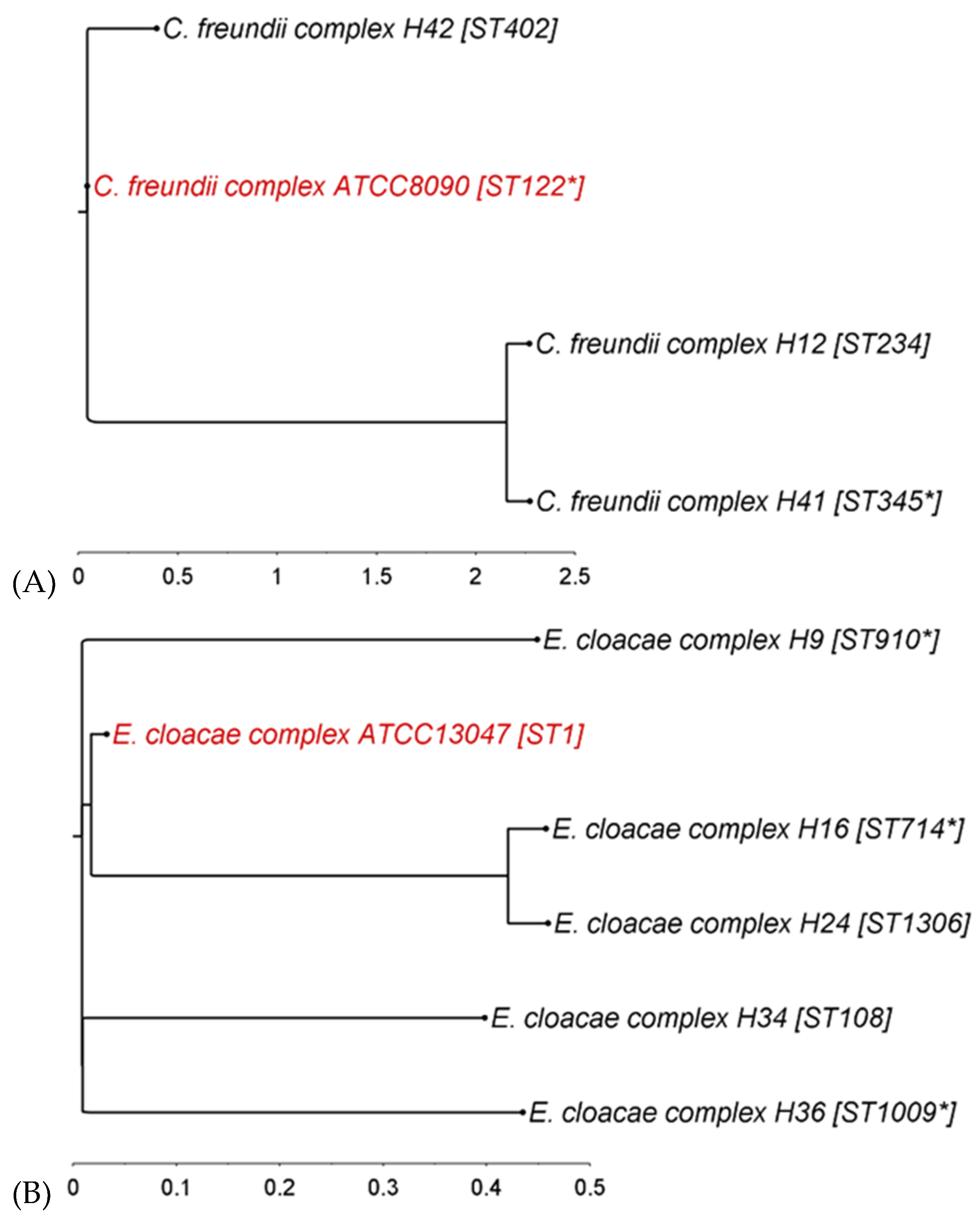
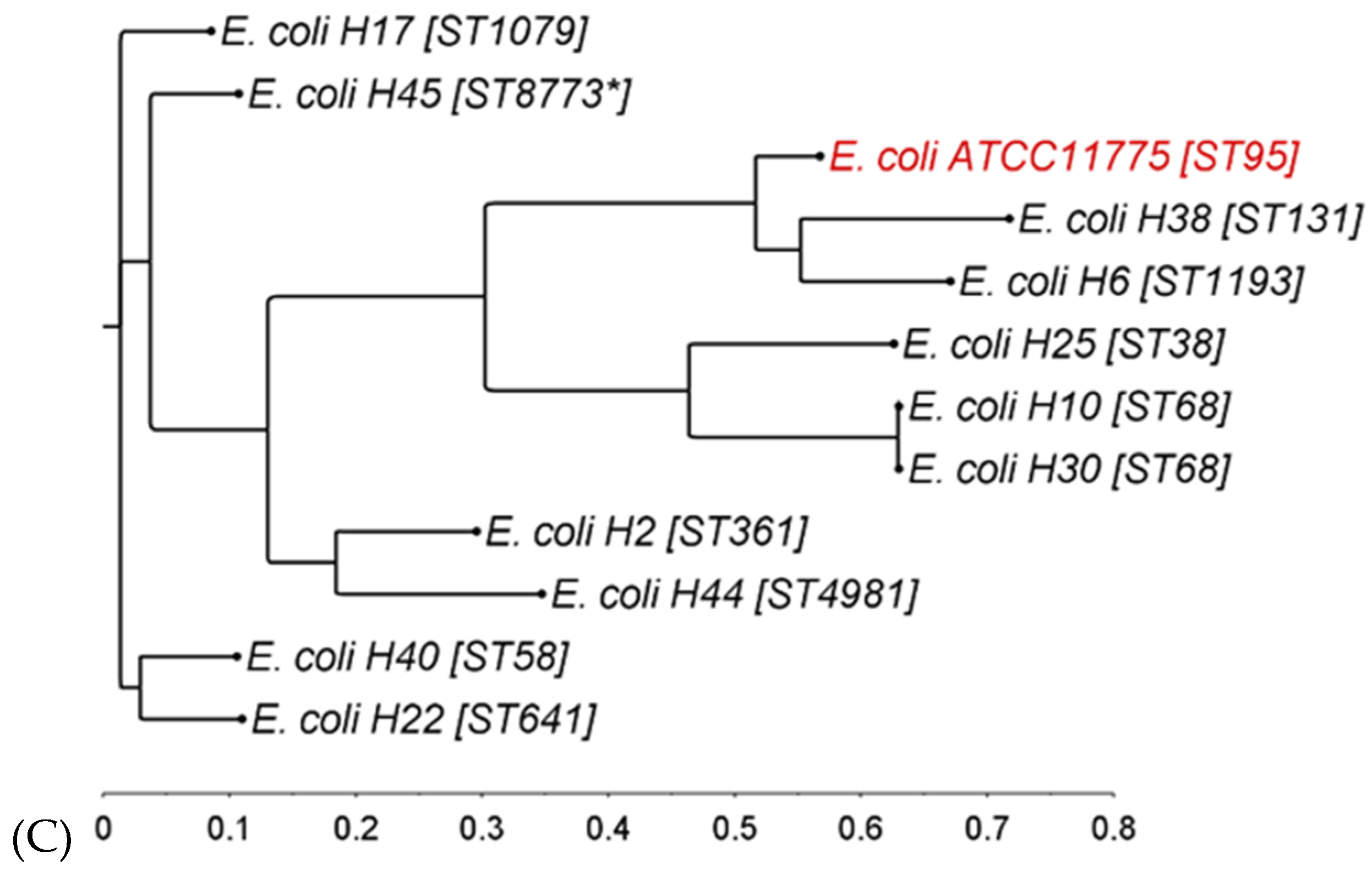
3.1. Phylogenetic Diversity of Donor Strains
3.2. Antibiotic Resistance and Plasmid Profiles of Donor Strains
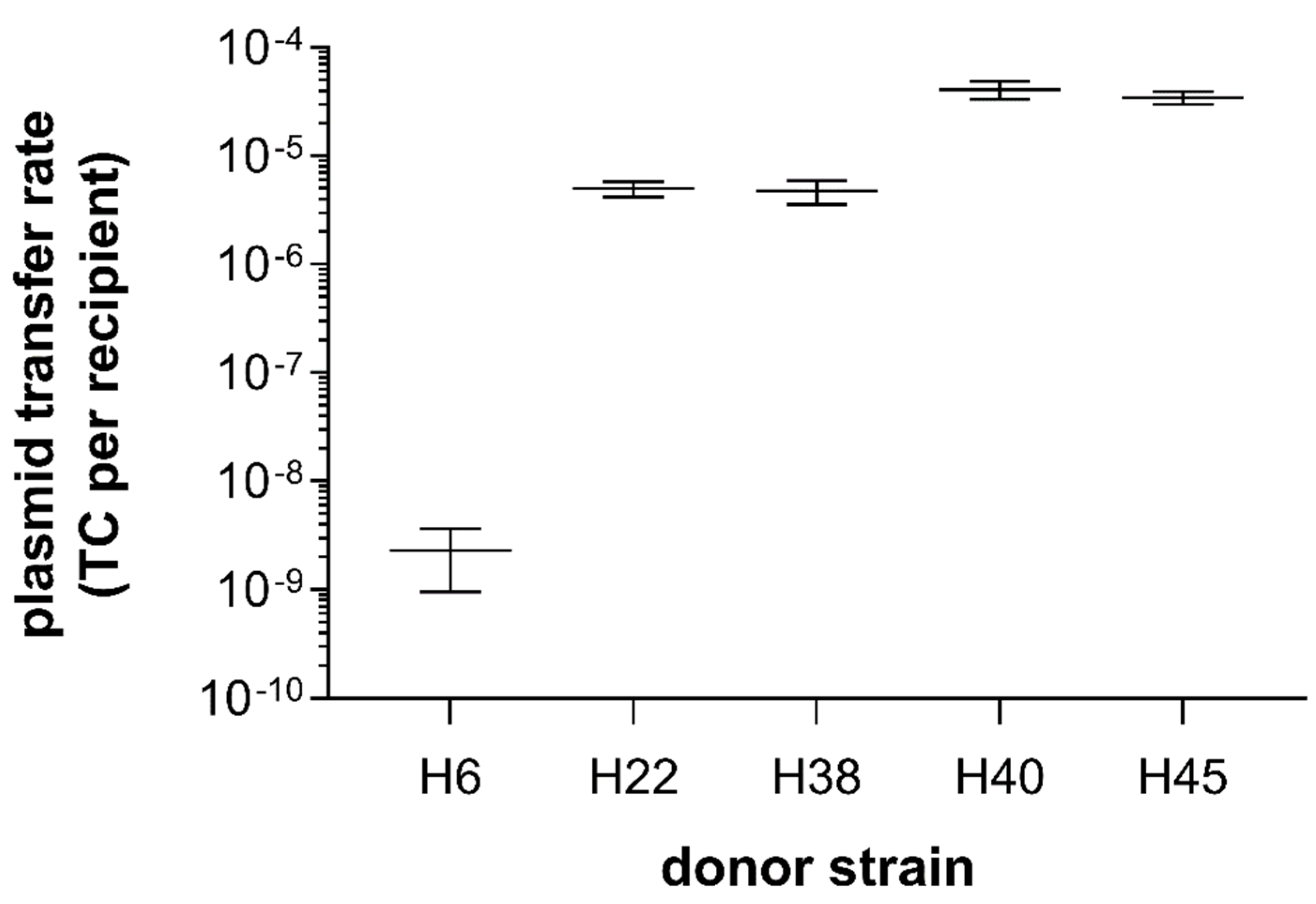
3.3. Filter and Broth Conjugation
3.4. Donor and Transconjugant Strain Comparison
3.4.1. Phenotypic Antibiotic Resistance
3.4.2. Molecular Detection of ARGs and Resistance Point Mutations
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Cycling Step | PCR Conditions | ||
|---|---|---|---|
| Enzyme activation | 95 °C, 5 min |  | |
| Denaturation | 94 °C, 3 s | ||
| 92 °C, 30 s | 30 cycles | ||
| Annealing | 40 °C, 1 min | ||
| Extension | 65 °C, 8 min | ||
| Final extension | 65 °C, 16 min | ||
References
- Kupferschmidt, K. Resistance fighters. Science 2016, 352, 758–761. [Google Scholar] [CrossRef]
- Morar, M.; Wright, G.D. The genomic enzymology of antibiotic resistance. Annu. Rev. Genet. 2010, 44, 25–51. [Google Scholar] [CrossRef] [PubMed]
- Svara, F.; Rankin, D.J. The evolution of plasmid-carried antibiotic resistance. BMC Evol. Biol. 2011, 11, 130. [Google Scholar] [CrossRef] [PubMed]
- Thomas, C.M.; Nielsen, K.M. Mechanisms of, and barriers to, horizontal gene transfer between bacteria. Nat. Rev. Microbiol. 2005, 3, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (CDC): Antibiotic/Antimicrobial Resistance—Biggest Threats; c2019. Available online: https://www.cdc.gov/drugresistance/biggest_threats.html (accessed on 29 June 2020).
- Ambler, R.P. The structure of beta-lactamases. Philos. Trans. R. Soc. Lond. 1980, 289, 321–331. [Google Scholar]
- Rawat, D.; Nair, D. Extended-spectrum β-lactamases in Gram Negative Bacteria. J. Glob. Infect. Dis. 2010, 2, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Dahmen, S.; Métayer, V.; Gay, E.; Madec, J.-Y.; Haenni, M. Characterization of extended-spectrum beta-lactamase (ESBL)-carrying plasmids and clones of Enterobacteriaceae causing cattle mastitis in France. Vet. Microbiol. 2013, 162, 793–799. [Google Scholar] [CrossRef]
- Ewers, C.; Grobbel, M.; Bethe, A.; Wieler, L.H.; Guenther, S. Extended-spectrum beta-lactamases-producing Gram-negative bacteria in companion animals: Action is clearly warranted. Berl. Munch. Tierarztl. Wochenschr. 2011, 124, 4–101. [Google Scholar]
- Nüesch-Inderbinen, M.; Zurfluh, K.; Peterhans, S.; Hächler, H.; Stephan, R. Assessment of the prevalence of extended-spectrum beta-lactamase-producing Enterobacteriaceae in ready-to-eat salads, fresh-cut fruit, and sprouts from the Swiss market. J. Food Prot. 2015, 78, 1178–1181. [Google Scholar] [CrossRef]
- Szmolka, A.; Nagy, B. Multidrug resistant commensal Escherichia coli in animals and its impact for public health. Front. Microbiol. 2013, 4, 258. [Google Scholar] [CrossRef] [PubMed]
- Geser, N.; Stephan, R.; Hächler, H. Occurrence and characteristics of extended-spectrum β-lactamase (ESBL) producing Enterobacteriaceae in food producing animals, minced meat and raw milk. BMC Vet. Res. 2012, 8, 21. [Google Scholar] [CrossRef] [PubMed]
- Reuland, E.A.; al Naiemi, N.; Raadsen, S.A.; Savelkoul, P.H.M.; Kluytmans, J.A.J.W.; Vandenbroucke-Grauls, C.M.J.E. Prevalence of ESBL-producing Enterobacteriaceae in raw vegetables. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1843–1846. [Google Scholar] [CrossRef] [PubMed]
- Gekenidis, M.T.; Qi, W.; Hummerjohann, J.; Zbinden, R.; Walsh, F.; Drissner, D. Antibiotic-resistant indicator bacteria in irrigation water: High prevalence of extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli. PLoS ONE 2018, 13, e0207857. [Google Scholar] [CrossRef]
- Zurfluh, K.; Hächler, H.; Nüesch-Inderbinen, M.; Stephan, R. Characteristics of extended-spectrum beta-lactamase- and carbapenemase-producing Enterobacteriaceae isolates from rivers and lakes in Switzerland. Appl. Environ. Microbiol. 2013, 79, 3021–3026. [Google Scholar] [CrossRef]
- Graham, D.W.; Knapp, C.W.; Christensen, B.T.; McCluskey, S.; Dolfing, J. Appearance of β-lactam Resistance Genes in Agricultural Soils and Clinical Isolates over the 20th Century. Sci. Rep. 2016, 6, 21550. [Google Scholar] [CrossRef]
- Gatica, J.; Yang, K.; Pagaling, E.; Jurkevitch, E.; Yan, T.; Cytryn, E. Resistance of Undisturbed Soil Microbiomes to Ceftriaxone Indicates Extended Spectrum β-Lactamase Activity. Front. Microbiol. 2015, 6, 1233. [Google Scholar] [CrossRef]
- Guenther, S.; Ewers, C.; Wieler, L.H. Extended-spectrum beta-lactamases producing E. coli in wildlife, yet another form of environmental pollution? Front. Microbiol. 2011, 2, 246. [Google Scholar] [CrossRef]
- Cantón, R.; González-Alba, J.M.; Galán, J.C. CTX-M Enzymes: Origin and Diffusion. Front. Microbiol. 2012, 3, 110. [Google Scholar] [CrossRef]
- Reid, C.J.; Blau, K.; Jechalke, S.; Smalla, K.; Djordjevic, S.P. Whole Genome Sequencing of Escherichia coli From Store-Bought Produce. Front. Microbiol. 2020, 10, 3050. [Google Scholar] [CrossRef]
- Pesavento, G.; Calonico, C.; Ducci, B.; Magnanini, A.; Lo Nostro, A. Prevalence and antibiotic resistance of Enterococcus spp. isolated from retail cheese, ready-to-eat salads, ham, and raw meat. Food Microbiol. 2014, 41, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gekenidis, M.T.; Schöner, U.; von Ah, U.; Schmelcher, M.; Walsh, F.; Drissner, D. Tracing back multidrug-resistant bacteria in fresh herb production: From chive to source through the irrigation water chain. FEMS Microbiol. Ecol. 2018, 94, fiy149. [Google Scholar] [CrossRef] [PubMed]
- Vital, P.G.; Caballes, M.B.D.; Rivera, W.L. Antimicrobial resistance in Escherichia coli and Salmonella spp. isolates from fresh produce and the impact to food safety. J. Environ. Sci. Health B 2017, 52, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Gekenidis, M.T.; Gossin, D.; Schmelcher, M.; Schöner, U.; Remus-Emsermann, M.N.P.; Drissner, D. Dynamics of culturable mesophilic bacterial communities of three fresh herbs and their production environment. J. Appl. Microbiol. 2017, 123, 916–932. [Google Scholar] [CrossRef]
- Steele, M.; Odumeru, J. Irrigation water as source of foodborne pathogens on fruit and vegetables. J. Food Prot. 2004, 67, 2839–2849. [Google Scholar] [CrossRef]
- Czekalski, N.; Díez, E.G.; Bürgmann, H. Wastewater as a point source of antibiotic-resistance genes in the sediment of a freshwater lake. ISME J. 2014, 8, 1381–1390. [Google Scholar] [CrossRef]
- Liu, X.; Liang, C.; Liu, X.; Zhao, F.; Han, C. Occurrence and human health risk assessment of pharmaceuticals and personal care products in real agricultural systems with long-term reclaimed wastewater irrigation in Beijing, China. Ecotoxicol. Environ. Saf. 2020, 190, 110022. [Google Scholar] [CrossRef]
- Chen, Q.-L.; Cui, H.-L.; Su, J.-Q.; Penuelas, J.; Zhu, Y.-G. Antibiotic resistomes in plant microbiomes. Trends Plant. Sci. 2019, 24, 530–541. [Google Scholar] [CrossRef]
- SwissGAP: Risikoanalyse Bewässerungswasser (2017); c2018. Available online: https://agrosolution.ch/swissgap/ (accessed on 29 June 2020).
- Heuer, H.; Krögerrecklenfort, E.; Wellington, E.M.; Egan, S.; van Elsas, J.D.; van Overbeek, L.; Collard, J.M.; Guillaume, G.; Karagouni, A.D.; Nikolakopoulou, T.L.; et al. Gentamicin resistance genes in environmental bacteria: Prevalence and transfer. FEMS Microbiol. Ecol. 2002, 42, 289–302. [Google Scholar] [CrossRef]
- Gekenidis, M.-T.; Studer, P.; Wüthrich, S.; Brunisholz, R.; Drissner, D. Beyond the matrix-assisted laser desorption ionization (MALDI) biotyping workflow: In search of microorganism-specific tryptic peptides enabling discrimination of subspecies. Appl. Environ. Microbiol. 2014, 80, 4234–4241. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Pontén, T.S.; Ussery, D.W.; Aarestrup, F.M. Multilocus sequence typing of total genome sequenced bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef] [PubMed]
- Kaas, R.S.; Leekitcharoenphon, P.; Aarestrup, F.M.; Lund, O. Solving the problem of comparing whole bacterial genomes across different sequencing platforms. PLoS ONE 2014, 9, e104984. [Google Scholar] [CrossRef]
- Rambaut, A. FigTree v1.4.4, a Graphical Viewer of Phylogenetic Trees, 2018. 2014 Java. Available online: https://github.com/rambaut/figtree (accessed on 29 June 2020).
- Zankari, E.; Hasman, H.; Cosentino, S.; Vestergaard, M.; Rasmussen, S.; Lund, O.; Aarestrup, F.M.; Larsen, M.V. Identification of acquired antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 2640–2644. [Google Scholar] [CrossRef]
- Carattoli, A.; Zankari, E.; Garcìa-Fernandez, A.; Larsen, M.V.; Lund, O.; Villa, L.; Aarestrup, F.M.; Hasman, H. PlasmidFinder and pMLST: In silico detection and typing of plasmids. Antimicrob. Agents Chemother. 2014, 58, 3895–3903. [Google Scholar] [CrossRef]
- Blau, K.; Bettermann, A.; Jechalke, S.; Fornefeld, E.; Vanrobaeys, Y.; Stalder, T.; Top, E.M.; Smalla, K. The Transferable Resistome of Produce. mBio 2018, 9, e01300–e01318. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, B.R.; Mazumder, A. Comparative efficacy of five different rep-PCR methods to discriminate Escherichia coli populations in aquatic environments. Water Sci. Technol. 2008, 58, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V. Genomic Epidemiology—ResFinder 4 database 2020. Available online: https://bitbucket.org/genomicepidemiology/resfinder_db/src/master/phenotypes.txt (accessed on 29 June 2020).
- Kaase, M.; Szabados, F.; Anders, A.; Gatermann, S.G. Fosfomycin susceptibility in carbapenem-resistant Enterobacteriaceae from Germany. J. Clin. Microbiol. 2014, 52, 1893–1897. [Google Scholar] [CrossRef]
- Jolley, K.A.; Bray, J.E.; Maiden, M.C.J. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. Wellcome Open Res. 2018, 3, 124. [Google Scholar] [CrossRef]
- Izdebski, R.; Baraniak, A.; Herda, M.; Fiett, J.; Bonten, M.J.M.; Carmeli, Y.; Goossens, H.; Hryniewicz, W.; Brun-Buisson, C.; Gniadkowski, M. MLST reveals potentially high-risk international clones of Enterobacter cloacae. J. Antimicrob. Chemoth. 2014, 70, 48–56. [Google Scholar] [CrossRef]
- Girlich, D.; Poirel, L.; Nordmann, P. Clonal distribution of multidrug-resistant Enterobacter cloacae. Diagnost. Microbiol. Infect. Dis. 2015, 81, 264–268. [Google Scholar] [CrossRef]
- Jin, L.; Wang, R.; Wang, X.; Wang, Q.; Zhang, Y.; Yin, Y.; Wang, H. Emergence of mcr-1 and carbapenemase genes in hospital sewage water in Beijing, China. J. Antimicrob. Chemother. 2017, 73, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Falgenhauer, L.; Schwengers, O.; Schmiedel, J.; Baars, C.; Lambrecht, O.; Heß, S.; Berendonk, T.U.; Falgenhauer, J.; Chakraborty, T.; Imirzalioglu, C. Multidrug-Resistant and Clinically Relevant Gram-Negative Bacteria Are Present in German Surface Waters. Front. Microbiol 2019, 10, 2779. [Google Scholar] [CrossRef] [PubMed]
- Verdet, C.; Gautier, V.; Chachaty, E.; Ronco, E.; Hidri, N.; Decré, D.; Arlet, G. Genetic Context of Plasmid-Carried blaCMY-2-Like Genes in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 4002–4006. [Google Scholar] [CrossRef] [PubMed]
- Ku, Y.-H.; Lee, M.-F.; Chuang, Y.-C.; Yu, W.-L. Detection of Plasmid-Mediated β-Lactamase Genes and Emergence of a Novel AmpC (CMH-1) in Enterobacter cloacae at a Medical Center in Southern Taiwan. J. Clin. Med. 2018, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Piotrowska, M.; Kowalska, S.; Popowska, M. Diversity of β-lactam resistance genes in Gram-negative rods isolated from a municipal wastewater treatment plant. Ann. Microbiol. 2019, 69, 591–601. [Google Scholar] [CrossRef]



| Strain ID | Antibiotic Resistance Phenotypes (R and I) | Acquired Antibiotic Resistance Genes | Plasmid Replicons (% ID; HSP/Query) |
|---|---|---|---|
| Ent.H9 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, ETP, SMZ, SXT | blaACT-9, fosA†, mcr-9 | none detected |
| Ent.H16 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, ETP, F100 | blaACT-12, fosA2† | none detected |
| Ent.H24 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, ETP, F100 | blaACT-12, fosA2† | none detected |
| Ent.H34 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, ETP, F100 | blaACT-7 | none detected |
| Ent.H36 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, ETP, MI, NA, F100 | fosA†, qnrE1 | none detected |
| Citr.H12 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, FEP, NA, F100 | blaCMY-83 | none detected |
| Citr.H41 | AM, TEMO, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, FEP, ETP | blaCMY-101 | none detected |
| Citr.H42 | AM, AMC, TPZ, KF, FOX, CXM, CPD, CRO, CTX, CAZ, TE, MI, NA, TMP, SMZ, SXT | blaTEM-1B, blaCMY-48, tet(B), qnrB, dfrA17, sul1, sul2, aadA5 ˟, catA1 ˟ | IncQ1 (100; 529/796) * |
| Antibiotic Resistant Phenotypes | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strain ID | AMK | AMC | SAM | AM | ATM | FEP | CTX | CAZ | CXM | CIP | LVX | MI | PIP | TE | TOB | SXT |
| E. coli CV601 | I | S | S | S | S | S | S | S | S | S | S | S | S | S | I | S |
| H6 | S | S | R | R | + | R | + | + | R | R | R | S | R | R | S | R |
| H6-TC | S | S | R | R | + | R | + | + | R | S | S | S | R | R | S | R |
| H22 | S | R | R | R | + | R | + | + | R | S | S | I | R | R | S | S |
| H22-TC | I | R | R | R | + | R | + | + | R | S | S | I | R | R | I | S |
| H38 | S | S | R | R | + | R | + | + | R | R | R | S | R | S | S | S |
| H38-TC | I | S | R | R | + | R | + | + | R | S | S | S | R | S | S | S |
| H40 | S | R | R | R | + | R | + | + | R | S | S | I | R | R | S | R |
| H40-TC | I | R | R | R | + | R | + | + | R | S | S | S | R | R | I | S |
| H45 | S | R | R | R | + | R | + | + | R | S | S | S | R | S | S | S |
| H45-TC | R | R | R | R | + | R | + | + | R | S | S | S | R | S | R | S |
| ESBL-Producing Donors | Transconjugants | ||||||
|---|---|---|---|---|---|---|---|
| ID | Acquired ARGs (Grouped by Contig) | Plasmid Replicons (% ID) | Point Mutations | ID | Acquired ARGs (Grouped by Contig) | Plasmid Replicons (% ID) | Point Mutations |
| H6 | I. strA, strB, aadA5, blaCTX-M-27, mph(A), sul1, sul2, tet(A), dfrA17 | I. IncFIA (99.74), IncFIB (96.63) | parE p.L416F parC p.S80I gyrA p.S83L gyrA p.D87N | H6-TC | I. strA, strB, aadA5, blaCTX-M-27, mph(A), sul1, sul2, tet(A), dfrA17 II. aph(3′)-III | none detected | none detected |
| H22 | I. blaCTX-M-1, sul2, tet(A) | I. IncI1 (98.59) | none detected | H22-TC | I. blaCTX-M-1, sul2, tet(A) II. aph(3′)-III | I. IncI1 (99.3) | none detected |
| H38 | I. blaCTX-M-15, mph(A) | I. IncFIB (98.39) | parE p.I529L parC p.S80I parC p.E84V gyrA p.S83L gyrA p.D87N | H38-TC | I. blaCTX-M-15, mph(A) II. aph(3′)-III | I. IncFIB (98.39) | none detected |
| H40 | I. blaCTX-M-1, sul2, tet(A) II. strA, strB, aadA1, sul1, sul2, tet(A), dfrA1 | I. IncI1 (98.59) II. IncFIB (97.07), IncFIC (95.59) | none detected | H40-TC | I. blaCTX-M-1, sul2, tet(A) II. aph(3′)-III | I. IncI1 (98.59) | none detected |
| H45 | I. blaCTX-M-1 | I. IncI1 (98.59) | none detected | H45-TC | I. blaCTX-M-1 II. aph(3′)-III | I. IncI1 (99.3) | none detected |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gekenidis, M.-T.; Kläui, A.; Smalla, K.; Drissner, D. Transferable Extended-Spectrum β-Lactamase (ESBL) Plasmids in Enterobacteriaceae from Irrigation Water. Microorganisms 2020, 8, 978. https://doi.org/10.3390/microorganisms8070978
Gekenidis M-T, Kläui A, Smalla K, Drissner D. Transferable Extended-Spectrum β-Lactamase (ESBL) Plasmids in Enterobacteriaceae from Irrigation Water. Microorganisms. 2020; 8(7):978. https://doi.org/10.3390/microorganisms8070978
Chicago/Turabian StyleGekenidis, Maria-Theresia, Anita Kläui, Kornelia Smalla, and David Drissner. 2020. "Transferable Extended-Spectrum β-Lactamase (ESBL) Plasmids in Enterobacteriaceae from Irrigation Water" Microorganisms 8, no. 7: 978. https://doi.org/10.3390/microorganisms8070978
APA StyleGekenidis, M.-T., Kläui, A., Smalla, K., & Drissner, D. (2020). Transferable Extended-Spectrum β-Lactamase (ESBL) Plasmids in Enterobacteriaceae from Irrigation Water. Microorganisms, 8(7), 978. https://doi.org/10.3390/microorganisms8070978