A Genome-Scale Metabolic Model of 2,3-Butanediol Production by Thermophilic Bacteria Geobacillus icigianus


Abstract
1. Introduction
2. Materials and Methods
2.1. Model Reconstruction
2.2. Applied Constraints and Model Curation
2.3. Model Modification for 2,3-Butanediol Production
2.4. Flux Balance Analysis
2.5. Model Analysis for 2,3-Butanediol Production Optimization
3. Results
3.1. Model Reconstruction
3.2. iMK1321 Model Optimization for 2,3-Butanediol Production
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ji, X.-J.; Huang, H.; Ouyang, P.-K. Microbial 2,3-butanediol production: A state-of-the-art review. Biotechnol. Adv. 2011, 29, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Celińska, E.; Grajek, W. Biotechnological production of 2,3-butanediol—Current state and prospects. Biotechnol. Adv. 2009, 27, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Z.; Wang, X.; Huang, Y.; Huo, F.; Zhu, X.; Xi, L.; Lu, J.R. Thermophilic fermentation of acetoin and 2,3-butanediol by a novel Geobacillus strain. Biotechnol. Biofuels 2012, 5, 88. [Google Scholar] [CrossRef] [PubMed]
- Hussein, A.H.; Lisowska, B.K.; Leak, D.J. The Genus Geobacillus and Their Biotechnological Potential. In Advances in Applied Microbiology; Academic Press: Cambridge, MA, USA, 2015; Volume 92, pp. 1–48. [Google Scholar]
- Suzuki, H. Peculiarities and biotechnological potential of environmental adaptation by Geobacillus species. Appl. Microbiol. Biotechnol. 2018, 102, 10425–10437. [Google Scholar] [CrossRef]
- Bryanskaya, A.V.; Rozanov, A.S.; Slynko, N.M.; Shekhovtsov, S.V.; Peltek, S.E. Geobacillus icigianus sp. nov., a thermophilic bacterium isolated from a hot spring. Int. J. Syst. Evol. Microbiol. 2015, 65, 864–869. [Google Scholar] [CrossRef]
- Bryanskaya, A.V.; Rozanov, A.S.; Logacheva, M.D.; Kotenko, A.V.; Peltek, S.E. Draft Genome Sequence of Geobacillus icigianus Strain G1w1T Isolated from Hot Springs in the Valley of Geysers, Kamchatka (Russian Federation). Genome Announc. 2014, 2, e01098-14. [Google Scholar] [CrossRef]
- Orth, J.D.; Thiele, I.; Palsson, B.Ø. What is flux balance analysis? Nat. Biotechnol. 2010, 28, 245–248. [Google Scholar] [CrossRef]
- Simeonidis, V.; Price, N.D. Genome-scale modeling for metabolic engineering. J. Ind. Microbiol. Biotechnol. 2015, 42, 327–338. [Google Scholar] [CrossRef]
- Gu, C.; Kim, G.B.; Kim, W.J.; Kim, H.U.; Lee, S.Y. Current status and applications of genome-scale metabolic models. Genome Biol. 2019, 20, 121. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Arkin, A.P.; Cottingham, R.W.; Henry, C.S.; Harris, N.L.; Stevens, R.L.; Maslov, S.; Dehal, P.; Ware, D.; Perez, F.; Canon, S.; et al. KBase: The United States Department of Energy Systems Biology Knowledgebase. Nat. Biotechnol. 2018, 36, 566–569. [Google Scholar] [CrossRef] [PubMed]
- Latendresse, M. Efficiently gap-filling reaction networks. BMC Bioinform. 2014, 15, 225. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.-K.; Palsson, B.; Park, S.M.; Schilling, C.H.; Mahadevan, R. Genome-scale Reconstruction of Metabolic Network inBacillus subtilisBased on High-throughput Phenotyping and Gene Essentiality Data. J. Biol. Chem. 2007, 282, 28791–28799. [Google Scholar] [CrossRef]
- Devoid, S.; Overbeek, R.; DeJongh, M.; Vonstein, V.; Best, A.A.; Henry, C. Automated genome annotation and metabolic model reconstruction in the SEED and model SEED. In Systems Metabolic Engineering; Humana Press: Totowa, NJ, USA, 2013; pp. 17–45. [Google Scholar]
- Norsigian, C.J.; Pusarla, N.; McConn, J.L.; Yurkovich, J.T.; Dräger, A.; Palsson, B.; King, Z. BiGG Models 2020: Multi-strain genome-scale models and expansion across the phylogenetic tree. Nucleic Acids Res. 2020, 48, D402–D406. [Google Scholar] [CrossRef] [PubMed]
- Ebrahim, A.; Lerman, J.; Palsson, B.; Hyduke, D.R. COBRApy: COnstraints-Based Reconstruction and Analysis for Python. BMC Syst. Biol. 2013, 7, 74. [Google Scholar] [CrossRef]
- Lieven, C.; Beber, M.E.; Olivier, B.G.; Bergmann, F.T.; Ataman, M.; Babaei, P.; Bartell, J.A.; Blank, L.M.; Chauhan, S.; Correia, K.; et al. MEMOTE for standardized genome-scale metabolic model testing. Nat. Biotechnol. 2020, 38, 272–276. [Google Scholar] [CrossRef]
- Lisowska, B. Genomic analysis and metabolic modelling of Geobacillus Thermoglucosidasius NCIBM 11955. Ph.D. Thesis, University of Bath, Bath, UK, 2016. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Karp, P.D.; Billington, R.; Caspi, R.; Fulcher, C.A.; Latendresse, M.; Kothari, A.; Keseler, I.M.; Krummenacker, M.; Midford, P.E.; Ong, Q.; et al. The BioCyc collection of microbial genomes and metabolic pathways. Briefings Bioinform. 2019, 20, 1085–1093. [Google Scholar] [CrossRef]
- Kanehisa, M. The KEGG database. In Novartis Foundation Symposium; John Wiley: Chichester, NY, USA, 2002. [Google Scholar]
- Jia, X.; Liu, Y.; Han, Y. A thermophilic cell-free cascade enzymatic reaction for acetoin synthesis from pyruvate. Sci. Rep. 2017, 7, 4333. [Google Scholar] [CrossRef]
- Atsumi, S.; Li, Z.; Liao, J.C. Acetolactate Synthase from Bacillus subtilis Serves as a 2-Ketoisovalerate Decarboxylase for Isobutanol Biosynthesis in Escherichia coli. Appl. Environ. Microbiol. 2009, 75, 6306–6311. [Google Scholar] [CrossRef]
- Lewis, N.E.; Hixson, K.K.; Conrad, T.M.; Lerman, J.A.; Charusanti, P.; Polpitiya, A.D.; Adkins, J.N.; Schramm, G.; O Purvine, S.; Lopez-Ferrer, D.; et al. Omic data from evolved E. coli are consistent with computed optimal growth from genome-scale models. Mol. Syst. Biol. 2010, 6, 390. [Google Scholar] [CrossRef] [PubMed]
- Rocha, I.; Maia, P.; Evangelista, P.T.; Vilaça, P.; Soares, S.; Pinto, J.P.B.G.P.; Nielsen, J.; Patil, K.R.; Ferreira, E.C.; Rocha, M. OptFlux: An open-source software platform for in silico metabolic engineering. BMC Syst. Biol. 2010, 4, 45. [Google Scholar] [CrossRef] [PubMed]
- King, Z.A.; Dräger, A.; Ebrahim, A.; Sonnenschein, N.; E Lewis, N.; Palsson, B. Escher: A Web Application for Building, Sharing, and Embedding Data-Rich Visualizations of Biological Pathways. PLoS Comput. Biol. 2015, 11, e1004321. [Google Scholar] [CrossRef] [PubMed]
- Kolpakov, F.; Akberdin, I.; Kashapov, T.; Kiselev, L.; Kolmykov, S.K.; Kondrakhin, Y.; Kutumova, E.; Mandrik, N.; Pintus, S.; Ryabova, A.; et al. BioUML: An integrated environment for systems biology and collaborative analysis of biomedical data. Nucleic Acids Res. 2019, 47, W225–W233. [Google Scholar] [CrossRef]
- Brochado, A.R.; Andrejev, S.; Maranas, C.; Patil, K.R. Impact of Stoichiometry Representation on Simulation of Genotype-Phenotype Relationships in Metabolic Networks. PLoS Comput. Biol. 2012, 8, e1002758. [Google Scholar] [CrossRef]
- Becker, S.A.; Feist, A.M.; Mo, M.L.; Hannum, G.; Palsson, B.; Herrgard, M.J. Quantitative prediction of cellular metabolism with constraint-based models: The COBRA Toolbox. Nat. Protoc. 2007, 2, 727–738. [Google Scholar] [CrossRef]
- Aliyu, H.; Lebre, P.; Blom, J.; Cowan, D.; De Maayer, P. Phylogenomic re-assessment of the thermophilic genus Geobacillus. Syst. Appl. Microbiol. 2016, 39, 527–533. [Google Scholar] [CrossRef]
- López-Agudelo, V.A.; Mendum, T.A.; Laing, E.; Wu, H.; Baena, A.; Barrera, L.F.; Beste, D.J.V.; Rios-Estepa, R. A systematic evaluation of Mycobacterium tuberculosis Genome-Scale Metabolic Networks. PLoS Comput. Biol. 2020, 16, e1007533. [Google Scholar] [CrossRef]
- Robinson, J.L.; Kocabaş, P.; Wang, H.; Cholley, P.-E.; Cook, D.J.; Nilsson, A.; Anton, M.; Ferreira, R.; Domenzain, I.; Billa, V.; et al. An atlas of human metabolism. Sci. Signal. 2020, 13, eaaz1482. [Google Scholar] [CrossRef]
- Nogales, J.; Mueller, J.; Gudmundsson, S.; Canalejo, F.J.; Duque, E.; Monk, J.; Feist, A.M.; Ramos, J.L.; Niu, W.; Palsson, B.O. High-quality genome-scale metabolic modelling of Pseudomonas putida highlights its broad metabolic capabilities. Environ. Microbiol. 2019, 22, 255–269. [Google Scholar] [CrossRef]
- Ahmad, A.; Hartman, H.B.; Krishnakumar, S.; Fell, D.; Poolman, M.; Srivastava, S. A Genome Scale Model of Geobacillus thermoglucosidasius (C56-YS93) reveals its biotechnological potential on rice straw hydrolysate. J. Biotechnol. 2017, 251, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Cordova, L.T.; Long, C.; Venkataramanan, K.P.; Antoniewicz, M.R. Complete genome sequence, metabolic model construction and phenotypic characterization of Geobacillus LC300, an extremely thermophilic, fast growing, xylose-utilizing bacterium. Metab. Eng. 2015, 32, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Maia, P.; Rocha, I.; Rocha, M. An integrated framework for strain optimization. In Proceedings of the 2013 IEEE Congress on Evolutionary Computation, Cancun, Mexico, 20–23 June 2013; pp. 198–205. [Google Scholar] [CrossRef]
- Vilaça, P.; Maia, P.; Giesteira, H.; Rocha, I.; Rocha, M. Analyzing and designing cell factories with OptFlux. In Methods in Molecular Biology; Humana Press Inc.: New York, NY, USA, 2018; Volume 1716, pp. 37–76. ISBN 978-1-4939-7528-0. [Google Scholar]
- Lu, L.; Mao, Y.; Kou, M.; Cui, Z.; Jin, B.; Chang, Z.; Wang, Z.; Ma, H.; Chen, T. Engineering central pathways for industrial-level (3R)-acetoin biosynthesis in Corynebacterium glutamicum. Microb. Cell Factories 2020, 19, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Song, C.W.; Park, J.M.; Chung, S.C.; Lee, S.Y.; Song, H. Microbial production of 2,3-butanediol for industrial applications. J. Ind. Microbiol. Biotechnol. 2019, 46, 1583–1601. [Google Scholar] [CrossRef] [PubMed]
- Yim, H.; Haselbeck, R.; Niu, W.; Pujol-Baxley, C.; Burgard, A.; Boldt, J.; Khandurina, J.; Trawick, J.D.; E Osterhout, R.; Stephen, R.; et al. Metabolic engineering of Escherichia coli for direct production of 1,4-butanediol. Nat. Methods 2011, 7, 445–452. [Google Scholar] [CrossRef]
- Biswas, R.; Yamaoka, M.; Nakayama, H.; Kondo, T.; Yoshida, K.-I.; Bisaria, V.S.; Kondo, A. Enhanced production of 2,3-butanediol by engineered Bacillus subtilis. Appl. Microbiol. Biotechnol. 2012, 94, 651–658. [Google Scholar] [CrossRef]
- Li, L.; Zhang, L.; Li, K.; Wang, Y.; Gao, C.; Han, B.; Ma, C.; Xu, P. A newly isolated Bacillus licheniformis strain thermophilically produces 2,3-butanediol, a platform and fuel bio-chemical. Biotechnol. Biofuels 2013, 6, 123. [Google Scholar] [CrossRef]
- Xiao, Z.; Lu, J.R. Strategies for enhancing fermentative production of acetoin: A review. Biotechnol. Adv. 2014, 32, 492–503. [Google Scholar] [CrossRef]
- Nguyen, A.D.; Hwang, I.Y.; Lee, O.K.; Kim, D.; Kalyuzhnaya, M.G.; Mariyana, R.; Hadiyati, S.; Kim, M.S.; Lee, E.Y. Systematic metabolic engineering of Methylomicrobium alcaliphilum 20Z for 2,3-butanediol production from methane. Metab. Eng. 2018, 47, 323–333. [Google Scholar] [CrossRef]



{kind=link}
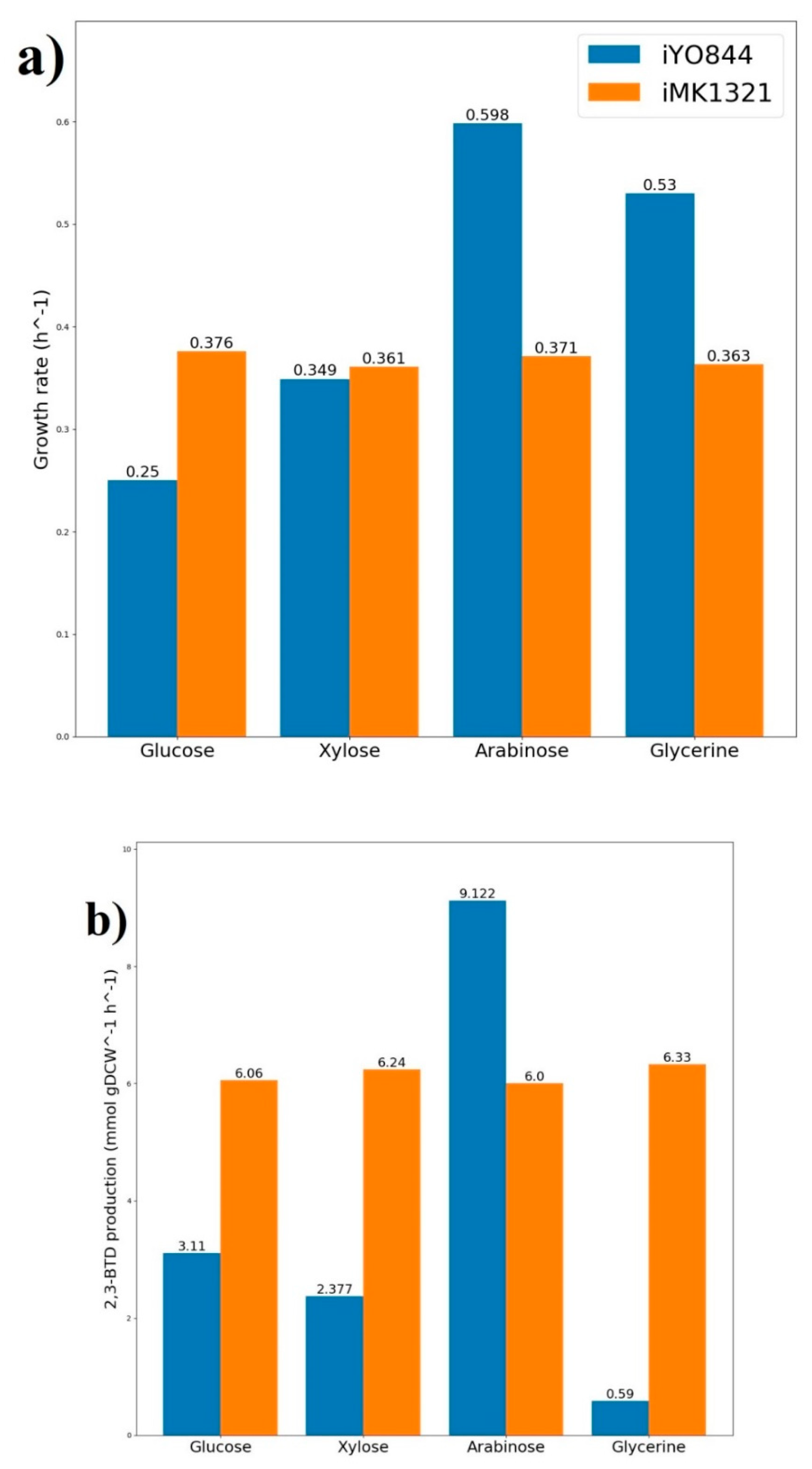{kind=link}
| Species | # of Genes in the Model | # of Reactions in the Model | # of Metabolites in the Model | Genome Size and RefSeq ID | # of Protein-Coding Genes | Reference |
|---|---|---|---|---|---|---|
| Geobacillus thermoglucosidasius (C56-YS93) | 736 | 1159 | 1163 | NZ_CP012712.1 Size = 3.87 Mb | 3659 | [35] |
| Geobacillus thermoglucosidasius (NCIMB 11955) | 859 | 1011 | 1050 | NZ_CP012712.1 Size = 3.87 Mb | 3615 | [19] |
| Geobacillus icigianus (G1W1) | 1321 | 1676 | 1589 | NZ_JPYA00000000.1 Size = 3.46 Mb | 3183 | this study |
| Organism | Growth Rate on Glucose (h−1) | Growth Rate on Xylose (h−1) | Growth Rate on Arabinose (h−1) | Growth Rate on Glycerine (h−1) |
|---|---|---|---|---|
| B.subtilis * | 0.118 | 0.113 | 0.128 | 0.140 |
| G.icigianus | 0.502 | 0.497 | 0.497 | 0.502 |
| B.subtilis ** | 0.624 | 0.624 | 0.624 | 0.624 |
| Substrate | Production of 2,3-Butanediol (mmol gDCW−1 h−1) | Growth Rate (h−1) | Reaction Modifications * |
|---|---|---|---|
| Glucose | 6.06 | 0.376 | CS = 0.03125 |
| GLYO1 = 0.03125 | |||
| Glucose | 6.06 | 0.376 | CS = 0.03125 |
| R00014 = 0.03125 | |||
| Glucose | 4.14 | 0.4136 | O2t = 0.03125 |
| Arabinose | 6.00 | 0.371 | CS = 0.03125 |
| Arabinose | 4.64 | 0.4 | CS = 0.25 |
| Xylose | 6.24 | 0.3612 | CS = 0.03125 |
| ALKP = 0.03125 | |||
| Xylose | 6.24 | 0.3612 | CS = 0.03125 |
| R01440 = 0.125 | |||
| Xylose | 6.14 | 0.3613 | CS = 0.03125 |
| SUCCt2r = 0.03125 | |||
| Glycerine | 6.33 | 0.363 | EX_o2_e = 0.5 |
| GF6PTA = 32.0 | |||
| Glycerine | 6.33 | 0.363 | EX_o2_e = 0.5 |
| Substrate | Production of 2,3-Butanediol (mmol gDCW−1 h−1) | Growth Rate (h−1) | Genetic Modifications ** | Reaction Modifications * |
|---|---|---|---|---|
| Glucose | 5.69 | 0.32 | EP10_15190 = 0.03125 | ACO1 = 0.03125 ACONTb = 0.03125 ACONTa = 0.03125 |
| EP10_02485 = 0.03125 | FEROpp = 0.03125 R00092 = 0.03125 | |||
| Glucose | 5.17 | 0.293 | EP10_09595 = 0.03125 | R0467 = 0.515625 R00014 = 0.7578125 FEROpp = 0.03125 R03050 = 0.515625 |
| EP10_15190 = 0.03125 | ACO1 = 0.03125 ACONTa = 0.03125 ACONTb = 0.03125 | |||
| Arabinose | 6.32 | 0.35 | KFX31147.1 = 16.0 | URIDK3 = 8.5 URA6_1 = 4.75 |
| KFX31511,1 = 0.03125 | ACO1 = 0.03125 ACONTa = 0.03125 ACONTb = 0.03125 | |||
| Arabinose | 6.30 | 0.35 | EP10_02065 = 0.03125 | GDH2 = 0.03125 GLUDy = 0.03125 |
| KFX31511.1 = 0.03125 | ACO1 = 0.03125 ACONTa = 0.03125 ACONTb = 0.03125 | |||
| Xylose | 5.95 | 0.34 | EP10_12945 = 4.0 | PGM = 2.5 |
| EP10_15190 = 0.03125 | ACO1 = 0.03125 ACONTa = 0.03125 ACONTb = 0.03125 | |||
| Xylose | 5.95 | 0.34 | EP10_12945 = 4.0 | PGM = 2.5 |
| KFX31511.1 = 0.03125 | ACO1 = 0.03125 ACONTa = 0.03125 ACONTb = 0.03125 | |||
| Glycerine | 6.55 | 0.357 | EP10_13815 = 0.03125 | FBA = 0.03125 FBA2 = 0.03125 FBA3 = 0.03125 |
| EP10_15190 = 0.03125 | ACO1 = 0.03125 ACONTa = 0.03125 ACONTb = 0.03125 | |||
| Glycerine | 6.55 | 0.357 | EP10_13815 = 0.0625 | FBA = 0.0625 FBA2 = 0.0625 FBA3 = 0.0625 |
| EP10_15190 = 0.03125 | ACO1 = 0.03125 ACONTa = 0.03125 ACONTb = 0.03125 | |||
| Glycerine | 6.46 | 0.359 | EP10_10240 = 0.03125 | R01056 = 0.03125 |
| EP10_15190 = 0.03125 | ACO1 = 0.03125 ACONTa = 0.03125 ACONTb = 0.03125 |
| Substrate | Production of 2,3-Butanediol (mmol gDCW−1 h−1) | Growth Rate (h−1) | Reaction Modifications * |
|---|---|---|---|
| Glucose | 4.16 | 0.42 | EX_o2_e = 0.03125 |
| Arabinose | 4.11 | 0.41 | O2t = 0.03125 |
| Xylose | 4.12 | 0.41 | O2t = 0.03125 |
| FE3abc = 0.03125 | |||
| Xylose | 4.11 | 0.41 | O2t = 0.03125 |
| Glycerine | 12.23 | 0.25 | O2t = 0.0625 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulyashov, M.; Peltek, S.E.; Akberdin, I.R. A Genome-Scale Metabolic Model of 2,3-Butanediol Production by Thermophilic Bacteria Geobacillus icigianus. Microorganisms 2020, 8, 1002. https://doi.org/10.3390/microorganisms8071002
Kulyashov M, Peltek SE, Akberdin IR. A Genome-Scale Metabolic Model of 2,3-Butanediol Production by Thermophilic Bacteria Geobacillus icigianus. Microorganisms. 2020; 8(7):1002. https://doi.org/10.3390/microorganisms8071002
Chicago/Turabian StyleKulyashov, Mikhail, Sergey E. Peltek, and Ilya R. Akberdin. 2020. "A Genome-Scale Metabolic Model of 2,3-Butanediol Production by Thermophilic Bacteria Geobacillus icigianus" Microorganisms 8, no. 7: 1002. https://doi.org/10.3390/microorganisms8071002
APA StyleKulyashov, M., Peltek, S. E., & Akberdin, I. R. (2020). A Genome-Scale Metabolic Model of 2,3-Butanediol Production by Thermophilic Bacteria Geobacillus icigianus. Microorganisms, 8(7), 1002. https://doi.org/10.3390/microorganisms8071002