Identification and Pathogenic Potential of Bacillus cereus Strains Isolated from a Dairy Processing Plant Producing PDO Taleggio Cheese


 , ,
, ,
Abstract

1. Introduction
2. Materials and Methods
2.1. Experimental Plan
2.2. Sample Analysis
2.3. MALDI-TOF MS Identification
2.4. RAPD-PCR
2.5. Detection of Hemolysins, Hemolysin BL, Diarrheal Toxin, Phosphatidylcholine-Specific Phospholipase C, and Proteases
2.6. Detection of Toxin-Encoding Genes
2.7. Biofilm Formation
2.8. Statistical Analysis
3. Results
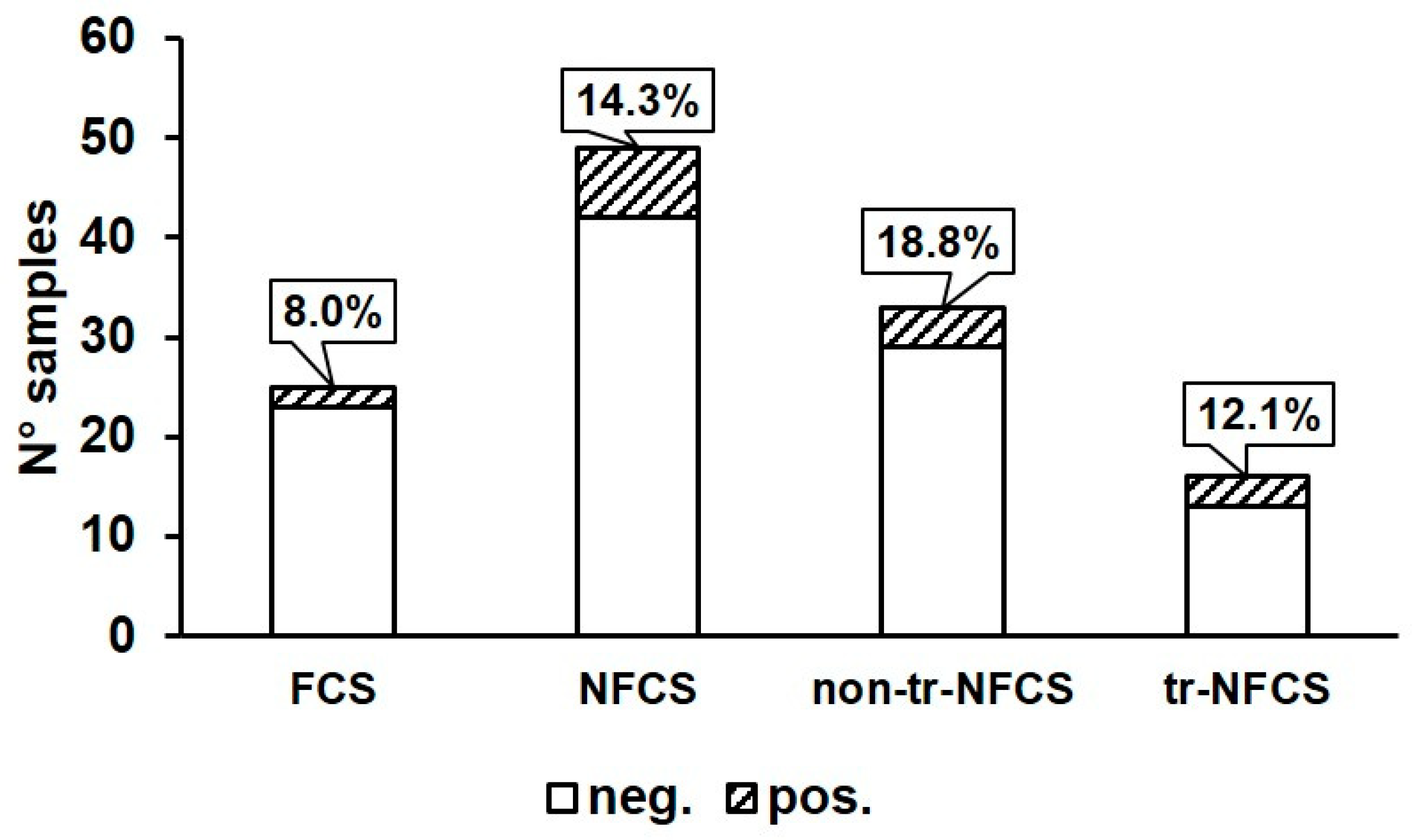
3.1. Prevalence of B. cereus in Environmental Samples, Curd, and Final Product
3.2. B. cereus Identification and Clustering
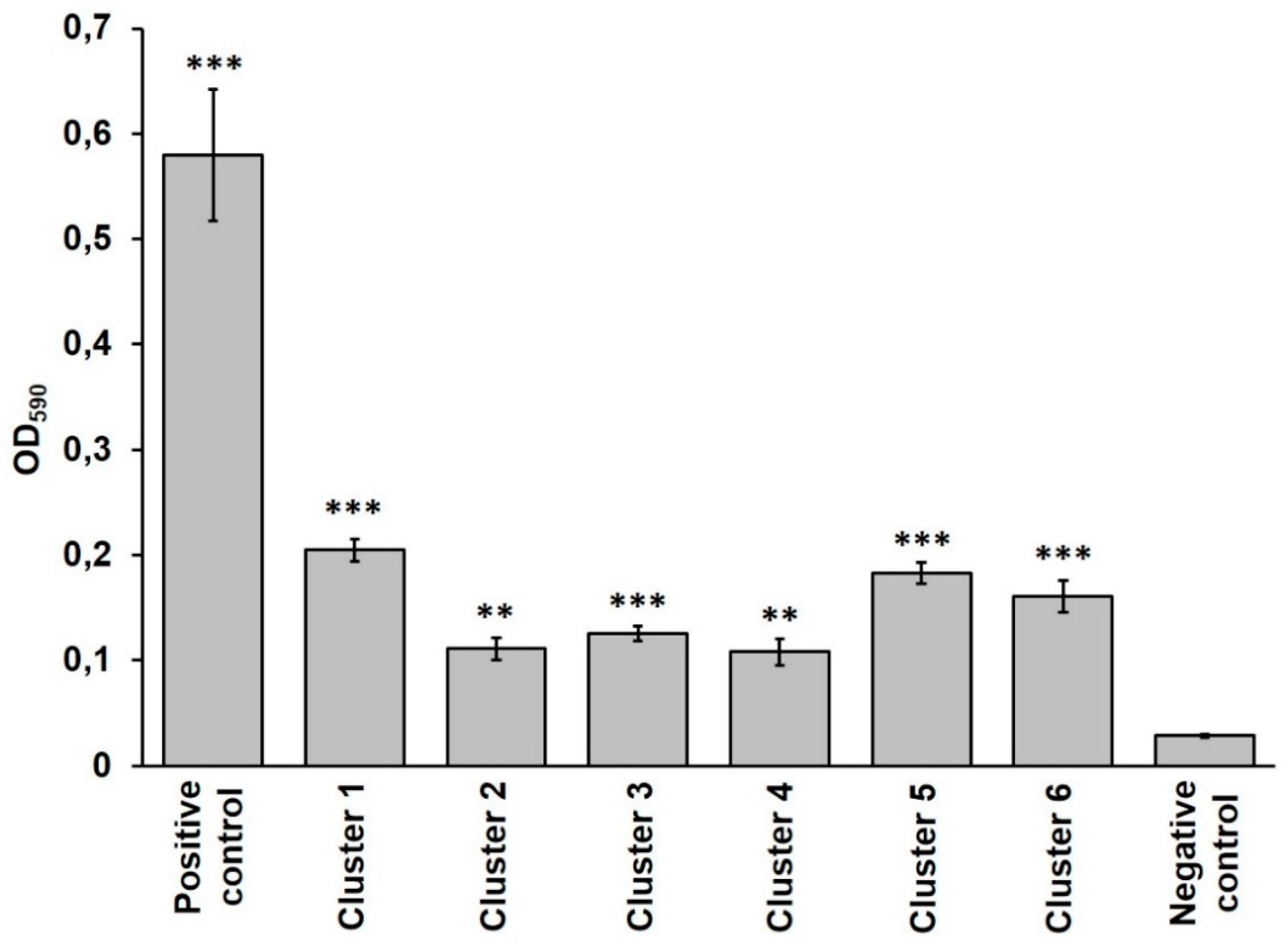
3.3. Virulence Potential of B. cereus Isolates
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Christiansson, A.; Bertilsson, J.; Svensson, B. Bacillus cereus spores in raw milk: Factors affecting the contamination of milk during the grazing period. J. Dairy Sci. 1999, 82, 305–314. [Google Scholar] [CrossRef]
- Svensson, B.; Monthán, A.; Shaheen, R.; Andersson, M.A.; Salkinoja-Salonen, M.; Christiansson, A. Occurrence of emetic toxin producing Bacillus cereus in the dairy production chain. Int. Dairy J. 2006, 16, 740–749. [Google Scholar] [CrossRef]
- Bartoszewicz, M.; Hansen, B.M.; Swiecick, I. The members of the Bacillus cereus group are commonly present contaminants of fresh and heat-treated milk. Food Microbiol. 2008, 25, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Wong, H.C.; Chang, M.H.; Fan, J.Y. Incidence and characterization of Bacillus cereus isolates contamination dairy products. Appl. Environ. Microbiol. 1988, 54, 699–702. [Google Scholar] [CrossRef] [PubMed]
- Spanu, C.; Scarano, C.; Spanu, V.; Pala, C.; Casti, D.; Lamon, S.; Cossu, F.; Ibba, M.; Nieddu, G.; De Santis, E.P.L. Occurrence and behavior of Bacillus cereus in naturally contaminated ricotta salata cheese during refrigerated storage. Food Microbiol. 2016, 58, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Tirloni, E.; Ghelardi, E.; Celandroni, F.; Bernardi, C.; Casati, R.; Rosshaug, P.S.; Stella, S. Bacillus cereus in fresh ricotta: Comparison of growth and Haemolysin BL production after artificial contamination during production or post processing. Food Control. 2017, 79, 272–278. [Google Scholar] [CrossRef]
- Tirloni, E.; Ghelardi, E.; Celandroni, F.; Bernardi, C.; Stella, S. Effect of dairy product environment on the growth of Bacillus Cereus. J. Dairy Sci. 2017, 100, 7026–7034. [Google Scholar] [CrossRef]
- Lücking, G.; Stoeckel, M.; Atamer, Z.; Hinrichs, J.; Ehling-Schulz, M. Characterization of aerobic spore-forming bacteria associated with industrial dairy processing environments and product spoilage. Int. J. Food Microbiol. 2013, 166, 270–279. [Google Scholar] [CrossRef]
- Kumari, S.; Sarkar, P.K. Bacillus cereus hazard and control in industrial dairy processing environment. Food Control. 2016, 69, 20–29. [Google Scholar] [CrossRef]
- Senesi, S.; Ghelardi, E. Production, secretion and biological activity of Bacillus cereus enterotoxins. Toxins 2010, 2, 1690–1703. [Google Scholar] [CrossRef]
- Ehling-Schulz, M.; Lereclus, D.; Koehler, T.M. The Bacillus cereus Group: Bacillus Species with Pathogenic Potential. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [PubMed]
- Celandroni, F.; Vecchione, A.; Cara, A.; Mazzantini, D.; Lupetti, A.; Ghelardi, E. Identification of Bacillus species: Implication on the quality of probiotic formulations. PLoS ONE 2019, 14, e0217021. [Google Scholar] [CrossRef] [PubMed]
- Celandroni, F.; Ghelardi, E.; Pastore, M.; Lupetti, A.; Kolstø, A.-B.; Senesi, S. Characterization of the chemotaxis fliY and cheA genes in Bacillus cereus. Fems Microbiol. Lett. 2000, 190, 247–253. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ghelardi, E.; Celandroni, F.; Salvetti, S.; Barsotti, C.; Baggiani, A.; Senesi, S. Identification and characterization of toxigenic Bacillus cereus isolates responsible for two food-poisoning outbreaks. Fems Microbiol. Lett. 2002, 208, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Celandroni, F.; Salvetti, S.; Gueye, S.A.; Mazzantini, D.; Lupetti, A.; Senesi, S.; Ghelardi, E. Identification and pathogenic potential of clinical Bacillus and Paenibacillus isolates. PLoS ONE 2016, 11, e0152831. [Google Scholar] [CrossRef]
- Shih, A.; Grant, M.; Sankey, C. Bacillus cereus Bacteremia and Matrix-Assisted Laser Desorption/Ionization Time-of-Flight Mass Spectrometry. Infect. Dis. Clin. Pract. 2016, 24, 14–17. [Google Scholar] [CrossRef]
- Ulrich, S.; Gottschalk, C.; Dietrich, R.; Märtlbauer, E.; Gareis, M. Identification of cereulide producing Bacillus cereus by MALDI-TOF MS. Food Microbiol. 2019, 82, 75–81. [Google Scholar] [CrossRef]
- Beecher, D.J.; Wong, A.C.L. Improved purification and characterization of hemolysin BL, a hemolytic dermonecrotic vascular permeability factor from Bacillus cereus. Infect. Immunol. 1994, 62, 980–986. [Google Scholar] [CrossRef]
- Gopal, N.; Hill, C.; Ross, P.R.; Beresford, T.P.; Fenelon, M.A.; Cott, P.D. The Prevalence and Control of Bacillus and Related Spore-Forming Bacteria in the Dairy Industry. Front. Microbiol. 2015, 6, 1418. [Google Scholar] [CrossRef]
- Berthold-Pluta, A.; Pluta, A.; Garbowska, M.; Stefańska, I. Prevalence and toxicity characterization of Bacillus cereus in food products from Poland. Foods 2019, 8, 269. [Google Scholar] [CrossRef] [PubMed]
- Pirttijärvi, T.S.M.; Ahonen, L.M.; Maunuksela, L.M.; Salkinoja-Salonen, M.S. Bacillus cereus in a whey process. Int. J. Food Microbiol. 1998, 44, 31–41. [Google Scholar] [CrossRef]
- Moradi-Khatoonabadi, Z.; Ezzatpanah, H.; Maghsoudlou, Y.; Khomeiri, M.; Aminafshar, M. Tracking Bacillus cereus in UF-Feta cheese processing line. Int. Dairy J. 2014, 39, 47–52. [Google Scholar] [CrossRef]
- Lin, S.; Schraft, A.; Odumeru, J.A.; Griffiths, W.G. Identification of contamination sources of Bacillus cereus in pasteurized milk. Int. J. Food Microbiol. 1998, 43, 159–171. [Google Scholar] [CrossRef]
- Wong, H.C.; Chen, Y. Effects of lactic acid bacteria and organic acids on growth and germination of Bacillus cereus. Appl. Environ. Microbiol. 1988, 54, 2179–2184. [Google Scholar] [CrossRef] [PubMed]
- Rukure, G.; Bester, B.H. Survival and growth of Bacillus cereus during Gouda cheese manufacturing. Food Control. 2001, 12, 31–36. [Google Scholar] [CrossRef]
- Muñoz, A.; Maqueda, M.; Gálvez, A.; Martínez-Bueno, M.; Rodríguez, A.; Valdivia, E. Biocontrol of psychrotrophic enterotoxigenic Bacillus cereus in a nonfat hard cheese by an enterococcal strain–producing Enterocin AS-48. J. Food Protect. 2004, 67, 1517–1521. [Google Scholar] [CrossRef]
- Sadek, Z.I.; Fathi, F.A.; Salem, M.M.E. Incidence, survival and biocontrol of psychrotrophic Bacillus cereus and its potential for toxin production in milk and Tallaga cheese. Pol. J. Food Nutr. Sci. 2006, 15, 419–425. [Google Scholar]
- Galli, A.; Franzetti, L.; Ottogalli, G. Characterization of surface bacterial microflora of Taleggio cheese. Ann. Microbiol. Enzim. 1996, 16, 369–377. [Google Scholar]
- Fontana, C.; Cappa, F.; Rebecchi, A.; Cocconcelli, P.S. Surface microbiota analysis of Taleggio, Gorgonzola, Casera, Scimudin, and Formaggio di Fossa Italian cheeses. Int. J. Food Microbiol. 2010, 138, 205–211. [Google Scholar] [CrossRef]
- Molva, C.; Sudagidan, M.; Okuklu, B. Extracellular enzyme production and enterotoxigenic gene profiles of Bacillus cereus and Bacillus thuringiensis strains isolated from cheese in Turkey. Food Control. 2009, 20, 829–834. [Google Scholar] [CrossRef]
- Moradi-Khatoonabadi, Z.; Ezzatpanah, H.; Maghsoudlou, Y.; Khomeiri, M.; Aminafshar, M. Bacillus cereus contamination of UF-Feta cheese during ripening and shelf life. J. Food Saf. 2015, 35, 41–49. [Google Scholar] [CrossRef]
- Yim, J.H.; Kim, K.Y.; Chon, J.W.; Kim, D.H.; Kim, H.S.; Choi, D.S.; Choi, I.S.; Seo, K.H. Incidence, antibiotic susceptibility, and toxin profiles of Bacillus cereus sensu lato isolated from Korean fermented soybean products. J. Food Sci. 2015, 80, M1266–M1270. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Liu, X.; Dietrich, R.; Märtlbauer, E.; Cao, J.; Ding, S.; Zhu, K. Characterization of Bacillus cereus isolates from local dairy farms in China. Fems Microbiol. Lett. 2016, 363, fnw096. [Google Scholar] [CrossRef] [PubMed]
- Yibar, A.; Çetinkaya, F.; Soyutemiz, E.; Yaman, G. Prevalence, enterotoxin production and antibiotic resistance of Bacillus cereus isolated from milk and cheese. Kafkas Univ. Vet. Fak. Derg. 2017, 23, 635–642. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, H.; He, J.; Yuan, Y.; Yuan, Z. The occurrence of Bacillus cereus, B. thuringiensis and B. mycoides in Chinese pasteurized full fat milk. Int. J. Food Microbiol. 2008, 121, 195–200. [Google Scholar] [CrossRef] [PubMed]
- De Santis, E.P.L.; Foddai, A.; Virdis, S.; Marongiu, P.; Pilo, A.L.; Scarano, C. Toxin gene pattern in Bacillus cereus group strains isolated from sheep ricotta cheese. Vet. Res. Commun. 2008, 32, S323–S326. [Google Scholar] [CrossRef] [PubMed]
- Nduhiu, G.J.; Ombui, J.N.; Nduati, D.W.; Gicheru, M.M. Genetic characterisation of food borne Bacillus cereus strains from milk, cheese and rice by multiplex PCR assay. Int. J. Integr. Biol. 2009, 5, 82–86. [Google Scholar]
- Castiaux, V.; Liu, X.; Delbrassinne, L.; Mahillon, J. Is Cytotoxin K from Bacillus cereus a bona fide enterotoxin? Int. J. Food Microbiol. 2015, 211, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.H.; Beuchat, L.R. Biofilm formation and sporulation by Bacillus cereus on a stainless steel surface and subsequent resistance of vegetative cells and spores to chlorine, chlorine dioxide, and a peroxyacetic acid–based sanitizer. J. Food Prot. 2005, 68, 2614–2622. [Google Scholar] [CrossRef]
- Srey, S.; Jahid, I.K.; Ha, S.D. Biofilm formation in food industries: A food safety concern. Food Control. 2013, 31, 572–585. [Google Scholar] [CrossRef]
- Majed, R.; Faille, C.; Kallassy, M.; Gohar, M. Bacillus cereus Biofilms-Same, Only Different. Front. Microbiol. 2016, 7, 1054. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Area | Typology | Sample | N° Positive/Analyzed Samples | Identified Isolates (Cluster Number *) |
|---|---|---|---|---|
| Production | FCS | Curdling tank | 0/3 | − |
| Table-grind-box-whey duct (internal surface) | 1/7 | A9M (3) | ||
| Operator’s hand | 0/2 | − | ||
| tr-NFCS | Door-handle | 0/1 | − | |
| Trolley wheels | 0/3 | − | ||
| Operator’s boots | 0/2 | − | ||
| non-tr-NFCS | Table-boxes- whey duct (external surface) | 0/5 | − | |
| Water hose | 1/2 | A20M (3) | ||
| Wall (drip) | 0/2 | − | ||
| Floor | 2/4 | A11M (1), A13M (2) | ||
| Drains | 0/6 | − | ||
| Total | 4/37 # | − | ||
| Brining/Ripening | FCS | Brining vat | 1/2 | A21M (5) |
| Brushing table | 0/2 | − | ||
| Brush | 0/2 | − | ||
| Operator’s gloves | 0/2 | − | ||
| Boxes-board-cheese cloth | 0/5 | − | ||
| tr-NFCS | Operator’s boots | 1/2 | A28M (2) | |
| Plastic doors | 0/1 | − | ||
| non-Tr-NFCS | Water hose | 0/1 | − | |
| Floor | 0/6 | − | ||
| Drains | 0/5 | − | ||
| Total | 2/28 # | − | ||
| Packaging | tr-NFCS | Trolley wheels | 1/2 | A39M (2) |
| non-Tr-NFCS | Drain | 1/2 | A30L (4) | |
| Total | 2/4 # | − | ||
| Sampler overshoes | tr-NFCS | 1/5 | A31M (2) | |
| Salt/ Saline | 0/3 | − | ||
| Curd | 2/2 | A29M (2), A32L (6) | ||
| Final product (PDO Taleggio) | 0/4 | − | ||
| Cluster 1 | Cluster 2 | Cluster 3 | Cluster 4 | Cluster 5 | Cluster 6 | |
|---|---|---|---|---|---|---|
| Production of virulence factors evaluated by phenotypic tests | ||||||
| PC-PLC | + | + | + | + | + | + |
| Proteases | + | + | + | + | + | + |
| Hemolysins | + | + | + | + | + | + |
| L2 component of HBL | + | + | − | + | + | + |
| HBL | + | + | − | + | - | + |
| Detection of toxin-encoding genes performed by PCR | ||||||
| nheA | + | + | + | + | + | + |
| nheB | + | + | + | + | − | + |
| nheC | + | + | + | − | − | + |
| sph | + | + | + | + | + | + |
| plcA | + | + | + | + | + | + |
| cytK | + | + | + | + | + | + |
| bcet | − | + | − | + | - | + |
| entFM | + | + | + | + | + | + |
| entS | + | + | − | + | − | + |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tirloni, E.; Stella, S.; Bernardi, C.; Mazzantini, D.; Celandroni, F.; Ghelardi, E. Identification and Pathogenic Potential of Bacillus cereus Strains Isolated from a Dairy Processing Plant Producing PDO Taleggio Cheese. Microorganisms 2020, 8, 949. https://doi.org/10.3390/microorganisms8060949
Tirloni E, Stella S, Bernardi C, Mazzantini D, Celandroni F, Ghelardi E. Identification and Pathogenic Potential of Bacillus cereus Strains Isolated from a Dairy Processing Plant Producing PDO Taleggio Cheese. Microorganisms. 2020; 8(6):949. https://doi.org/10.3390/microorganisms8060949
Chicago/Turabian StyleTirloni, Erica, Simone Stella, Cristian Bernardi, Diletta Mazzantini, Francesco Celandroni, and Emilia Ghelardi. 2020. "Identification and Pathogenic Potential of Bacillus cereus Strains Isolated from a Dairy Processing Plant Producing PDO Taleggio Cheese" Microorganisms 8, no. 6: 949. https://doi.org/10.3390/microorganisms8060949
APA StyleTirloni, E., Stella, S., Bernardi, C., Mazzantini, D., Celandroni, F., & Ghelardi, E. (2020). Identification and Pathogenic Potential of Bacillus cereus Strains Isolated from a Dairy Processing Plant Producing PDO Taleggio Cheese. Microorganisms, 8(6), 949. https://doi.org/10.3390/microorganisms8060949