An Intertwined Network of Regulation Controls Membrane Permeability Including Drug Influx and Efflux in Enterobacteriaceae


Abstract
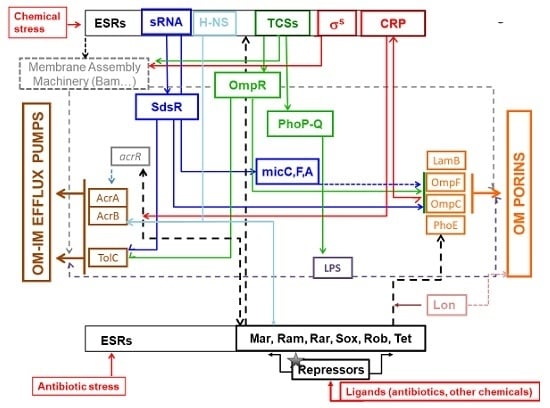
{kind=link}
{kind=link}
{kind=link}
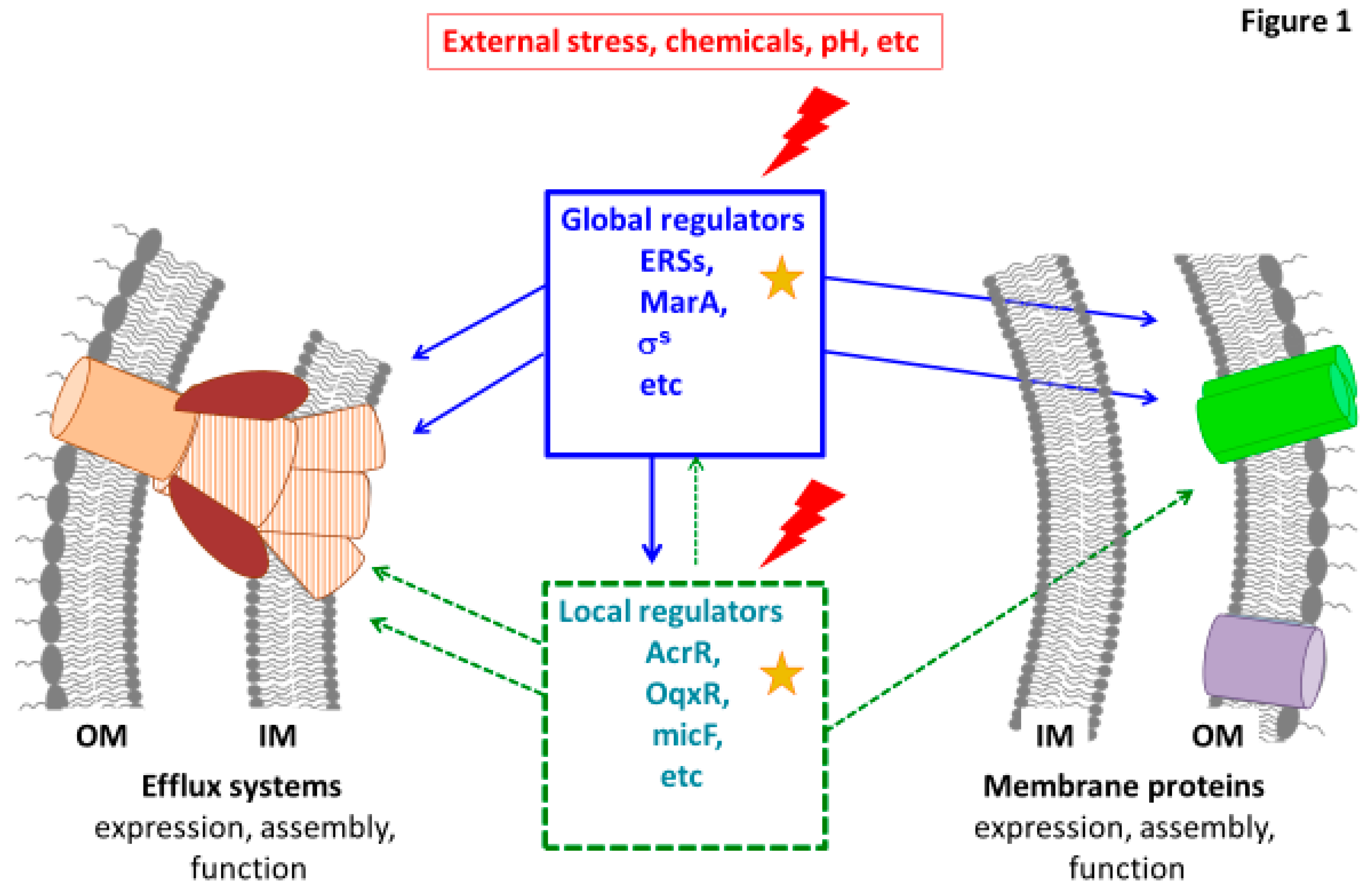
1. Introduction
2. Regulators Type 1—Helix-Turn-Helix (HTH) Family Regulators, Including the AraC/XylS Family
2.1. Positive Regulation by Global or Local Transcriptional Activators
2.1.1. The AraC-XylS Family
The Mar Regulon
The Oxidative Stress Regulon SoxRS
The Rob Regulon
The Regulator RamA
The Regulator RarA
TetD and NimR (YeaM)
2.1.2. Other Regulators
2.2. Repressors of Pump Genes Expression
2.2.1. TetR/AcrR Repressor Family
2.2.2. GntR Repressor Type: OqxR
3. Regulators Type 2—TCS and Other Regulators
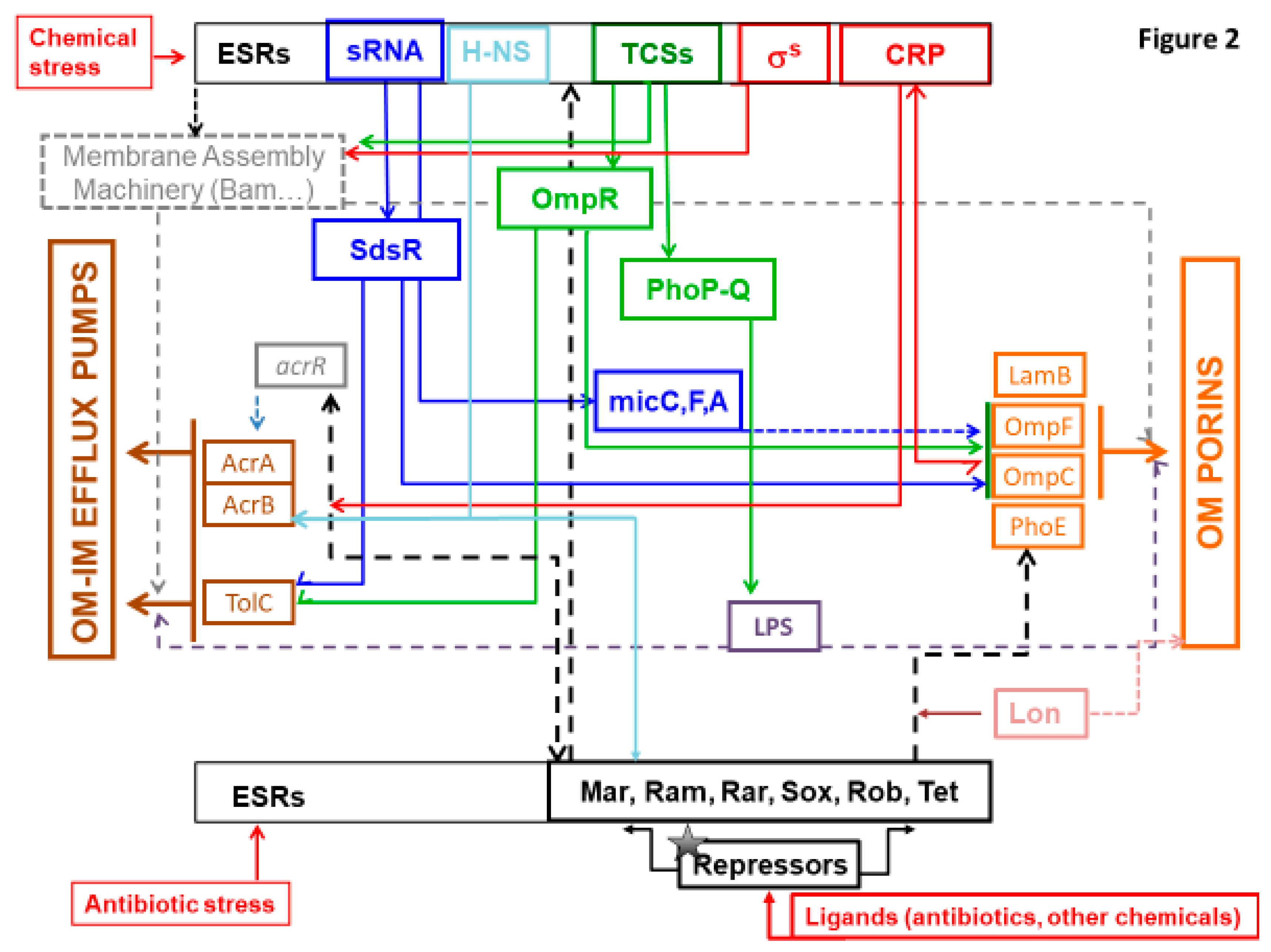
3.1. Envelope Stress Response (ESR) and TCS
3.2. ESR, TCS, and Antibiotic Resistance
3.2.1. INFLUX Modulation
3.2.2. EFFLUX Modulation
3.3. sRNAs
3.4. Other Mechanisms/Partners
3.5. Key External Factors Other Than Antibiotics
4. Concluding Remarks
Funding
Acknowledgments
Conflicts of Interest
References
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Matsoso, P.; Pant, S.; Brower, C.; Røttingen, J.-A.; Klugman, K.; Davies, S. Access to effective antimicrobials: A worldwide challenge. Lancet 2016, 387, 168–175. [Google Scholar] [CrossRef]
- Watkins, R.R.; Bonomo, R.A. Overview: Global and local impact of antibiotic resistance. Infect. Dis. Clin. N. Am. 2016, 30, 313–322. [Google Scholar] [CrossRef]
- Silhavy, T.J.; Kahne, D.; Walker, S. The bacterial cell envelope. Cold Spring Harb. Perspect. Biol. 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Winterhalter, M.; Pagès, J.-M. Outer Membrane Porins. In Bacterial Cell Walls and Membranes; Kuhn, A., Ed.; Subcellular Biochemistry; Springer: Cham, Switzerland, 2019; Volume 92, pp. 79–123. ISBN 978-3-030-18768-2. [Google Scholar]
- Braun, V.; Hantke, K. Lipoproteins: Structure, Function, Biosynthesis. In Bacterial Cell Walls and Membranes; Kuhn, A., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 39–77. ISBN 978-3-030-18768-2. [Google Scholar]
- Kudva, R.; Denks, K.; Kuhn, P.; Vogt, A.; Müller, M.; Koch, H.-G. Protein translocation across the inner membrane of Gram-negative bacteria: The Sec and Tat dependent protein transport pathways. Res. Microbiol. 2013, 164, 505–534. [Google Scholar] [CrossRef]
- Du, D.; Wang-Kan, X.; Neuberger, A.; van Veen, H.W.; Pos, K.M.; Piddock, L.J.V.; Luisi, B.F. Multidrug efflux pumps: Structure, function and regulation. Nat. Rev. Microbiol. 2018, 16, 523–539. [Google Scholar] [CrossRef]
- Vergalli, J.; Bodrenko, I.V.; Masi, M.; Moynié, L.; Acosta-Gutiérrez, S.; Naismith, J.H.; Davin-Regli, A.; Ceccarelli, M.; van den Berg, B.; Winterhalter, M.; et al. Porins and small-molecule translocation across the outer membrane of Gram-negative bacteria. Nat. Rev. Microbiol. 2020, 18, 164–176. [Google Scholar] [CrossRef]
- Zgurskaya, H.I.; Löpez, C.A.; Gnanakaran, S. Permeability barrier of Gram-negative cell envelopes and approaches to bypass it. ACS Infect. Dis. 2015, 1, 512–522. [Google Scholar] [CrossRef]
- Boos, W.; Shuman, H. Maltose/maltodextrin system of Escherichia coli: Transport, metabolism, and regulation. Microbiol. Mol. Biol. Rev. 1998, 62, 204–229. [Google Scholar] [CrossRef]
- Celia, H.; Noinaj, N.; Buchanan, S.K. Structure and Stoichiometry of the Ton Molecular Motor. Int. J. Mol. Sci. 2020, 21, 375. [Google Scholar] [CrossRef] [PubMed]
- Venter, H.; Chitsaz, M.; Brown, M.H. The role played by drug efflux pumps in bacterial multidrug resistance. Essays Biochem. 2017, 61, 127–139. [Google Scholar] [CrossRef] [PubMed]
- McNeil, H.E.; Alav, I.; Torres, R.C.; Rossiter, A.E.; Laycock, E.; Legood, S.; Kaur, I.; Davies, M.; Wand, M.; Webber, M.A.; et al. Identification of binding residues between periplasmic adapter protein (PAP) and RND efflux pumps explains PAP-pump promiscuity and roles in antimicrobial resistance. PLoS Pathog. 2019, 15, e1008101. [Google Scholar] [CrossRef] [PubMed]
- Weston, N.; Sharma, P.; Ricci, V.; Piddock, L.J.V. Regulation of the AcrAB-TolC efflux pump in Enterobacteriaceae. Res. Microbiol. 2018, 169, 425–431. [Google Scholar] [CrossRef]
- Nikaido, H.; Pagès, J.-M. Broad specificity efflux pumps and their role in multidrug resistance of Gram negative bacteria. FEMS Microbiol. Rev. 2012, 36, 340–363. [Google Scholar] [CrossRef]
- Grimsey, E.M.; Weston, N.; Ricci, V.; Stone, J.W.; Piddock, L.J.V. Over-expression of RamA, which regulates production of the multidrug resistance efflux pump AcrAB-TolC, increases mutation rate and influences drug-resistance phenotype. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Zhang, C.; Chen, S.; Bai, X.; Dedkova, L.M.; Hecht, S.M. Alteration of transcriptional regulator Rob in vivo: Enhancement of promoter DNA binding and antibiotic resistance in the presence of nucleobase amino acids. Biochemistry 2020, 59, 1217–1220. [Google Scholar] [CrossRef]
- Zaman, T.U.; Albladi, M.; Siddique, M.I.; Aljohani, S.M.; Balkhy, H.H. Insertion element mediated mgrB disruption and presence of ISKpn28 in colistin-resistant Klebsiella pneumoniae isolates from Saudi Arabia. Infect. Drug Resist. 2018, 11, 1183–1187. [Google Scholar] [CrossRef]
- Thota, S.S.; Chubiz, L.M. Multidrug resistance regulators MarA, SoxS, Rob and RamA repress flagella gene expression and motility in Salmonella enterica serovar Typhimurium. J. Bacteriol. 2019, 201. [Google Scholar] [CrossRef]
- Egan, S.M. Growing repertoire of AraC/XylS activators. J. Bacteriol. 2002, 184, 5529–5532. [Google Scholar] [CrossRef]
- Santiago, A.E.; Yan, M.B.; Tran, M.; Wright, N.; Luzader, D.H.; Kendall, M.M.; Ruiz-Perez, F.; Nataro, J.P. A large family of anti-activators accompanying XylS/AraC family regulatory proteins. Mol. Microbiol. 2016, 101, 314–332. [Google Scholar] [CrossRef] [PubMed]
- Majewski, P.; Gutowska, A.; Sacha, P.; Schneiders, T.; Talalaj, M.; Majewska, P.; Zebrowska, A.; Ojdana, D.; Wieczorek, P.; Hauschild, T.; et al. Expression of AraC/XylS stress response regulators in two distinct carbapenem-resistant Enterobacter cloacae ST89 biotypes. J. Antimicrob. Chemother. 2020. [Google Scholar] [CrossRef] [PubMed]
- Chollet, R.; Bollet, C.; Chevalier, J.; Malléa, M.; Pagès, J.-M.; Davin-Regli, A. mar operon involved in multidrug resistance of Enterobacter. aerogenes. Antimicrob. Agents Chemother. 2002, 46, 1093–1097. [Google Scholar] [CrossRef]
- Udani, R.A.; Levy, S.B. MarA-like regulator of multidrug resistance in Yersinia pestis. Antimicrob. Agents Chemother. 2006, 50, 2971–2975. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lee, J.O.; Cho, K.-S.; Kim, O.B. Overproduction of AcrR increases organic solvent tolerance mediated by modulation of SoxS regulon in Escherichia coli. Appl. Microbiol. Biotechnol. 2014, 98, 8763–8773. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, C.; Levy, S.B. Regulation of acrAB expression by cellular metabolites in Escherichia coli. J. Antimicrob. Chemother. 2014, 69, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Alekshun, M.N.; Levy, S.B.; Mealy, T.R.; Seaton, B.A.; Head, J.F. The crystal structure of MarR, a regulator of multiple antibiotic resistance, at 2.3 A resolution. Nat. Struct. Biol. 2001, 8, 710–714. [Google Scholar] [CrossRef]
- Linde, H.-J.; Notka, F.; Metz, M.; Kochanowski, B.; Heisig, P.; Lehn, N. In vivo increase in resistance to ciprofloxacin in Escherichia coli associated with deletion of the C-terminal part of MarR. Antimicrob. Agents Chemother. 2000, 44, 1865–1868. [Google Scholar] [CrossRef]
- Fang, L.; Chen, Q.; Shi, K.; Li, X.; Shi, Q.; He, F.; Zhou, J.; Yu, Y.; Hua, X. Step-wise increase in tigecycline resistance in Klebsiella pneumoniae associated with mutations in ramR, lon and rpsJ. PLoS ONE 2016, 11, e0165019. [Google Scholar] [CrossRef]
- Schneiders, T.; Barbosa, T.M.; McMurry, L.M.; Levy, S.B. The Escherichia coli transcriptional regulator MarA directly represses transcription of purA and hdeA. J. Biol. Chem. 2004, 279, 9037–9042. [Google Scholar] [CrossRef]
- Sharma, P.; Haycocks, J.R.J.; Middlemiss, A.D.; Kettles, R.A.; Sellars, L.E.; Ricci, V.; Piddock, L.J.V.; Grainger, D.C. The multiple antibiotic resistance operon of enteric bacteria controls DNA repair and outer membrane integrity. Nat. Commun. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Chetri, S.; Das, B.J.; Bhowmik, D.; Chanda, D.D.; Chakravarty, A.; Bhattacharjee, A. Transcriptional response of mar, sox and rob regulon against concentration gradient carbapenem stress within Escherichia coli isolated from hospital acquired infection. BMC Res. Notes 2020, 13. [Google Scholar] [CrossRef] [PubMed]
- Gaudu, P.; Moon, N.; Weiss, B. Regulation of the soxRS oxidative stress regulon: Reversible oxidation of the Fe-S centers of SoxR in vivo. J. Biol. Chem. 1997, 272, 5082–5086. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.G.; Gillette, W.K.; Rhee, S.; Rosner, J.L. Structural requirements for marbox function in transcriptional activation of mar/sox/rob regulon promoters in Escherichia coli: Sequence, orientation and spatial relationship to the core promoter. Mol. Microbiol. 1999, 34, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-J.; Gottesman, S. sRNA roles in regulating transcriptional regulators: Lrp and SoxS regulation by sRNAs. Nucleic Acids Res. 2016, 44, 6907–6923. [Google Scholar] [CrossRef]
- Martin, R.G.; Gillette, W.K.; Rosner, J.L. Promoter discrimination by the related transcriptional activators MarA and SoxS: Differential regulation by differential binding. Mol. Microbiol. 2000, 35, 623–634. [Google Scholar] [CrossRef] [PubMed]
- Koutsolioutsou, A.; Peña-Llopis, S.; Demple, B. Constitutive soxR mutations contribute to multiple-antibiotic resistance in clinical Escherichia coli isolates. Antimicrob. Agents Chemother. 2005, 49, 2746–2752. [Google Scholar] [CrossRef] [PubMed]
- Händel, N.; Hoeksema, M.; Freijo Mata, M.; Brul, S.; ter Kuile, B.H. Effects of stress, reactive oxygen species, and the SOS response on de novo acquisition of antibiotic resistance in Escherichia coli. Antimicrob. Agents Chemother. 2016, 60, 1319–1327. [Google Scholar] [CrossRef]
- Ariza, R.R.; Li, Z.; Ringstad, N.; Demple, B. Activation of multiple antibiotic resistance and binding of stress-inducible promoters by Escherichia coli Rob protein. J. Bacteriol. 1995, 177, 1655–1661. [Google Scholar] [CrossRef] [PubMed]
- Jair, K.W.; Yu, X.; Skarstad, K.; Thöny, B.; Fujita, N.; Ishihama, A.; Wolf, R.E. Transcriptional activation of promoters of the superoxide and multiple antibiotic resistance regulons by Rob, a binding protein of the Escherichia coli origin of chromosomal replication. J. Bacteriol. 1996, 178, 2507–2513. [Google Scholar] [CrossRef]
- Pletzer, D.; Schweizer, G.; Weingart, H. AraC/XylS family stress response regulators Rob, SoxS, PliA, and OpiA in the fire blight pathogen Erwinia amylovora. J. Bacteriol. 2014, 196, 3098–3110. [Google Scholar] [CrossRef] [PubMed]
- Bennik, M.H.J.; Pomposiello, P.J.; Thorne, D.F.; Demple, B. Defining a rob regulon in Escherichia coli by using transposon mutagenesis. J. Bacteriol. 2000, 182, 3794–3801. [Google Scholar] [CrossRef] [PubMed]
- Schneiders, T.; Levy, S.B. MarA-mediated transcriptional repression of the rob promoter. J. Biol. Chem. 2006, 281, 10049–10055. [Google Scholar] [CrossRef]
- Davin-Regli, A.; Pagès, J.M. Regulation of efflux pumps in Enterobacteriaceae: Genetic and chemical effectors. In Multiple Drug Resistance Bacteria, 2nd ed.; Carlos, F., Ed.; Horizon Bioscience: Wymondham, UK, 2006. [Google Scholar]
- Davin-Regli, A.; Bolla, J.-M.; James, C.E.; Lavigne, J.-P.; Chevalier, J.; Garnotel, E.; Molitor, A.; Pagès, J.-M. Membrane permeability and regulation of drug “influx and efflux” in enterobacterial pathogens. Curr. Drug Targets 2008, 9, 750–759. [Google Scholar] [CrossRef] [PubMed]
- Chollet, R.; Chevalier, J.; Bollet, C.; Pages, J.-M.; Davin-Regli, A. RamA is an alternate activator of the multidrug resistance cascade in Enterobacter aerogenes. Antimicrob. Agents Chemother. 2004, 48, 2518–2523. [Google Scholar] [CrossRef] [PubMed]
- Schneiders, T.; Amyes, S.G.B.; Levy, S.B. Role of AcrR and RamA in fluoroquinolone resistance in clinical Klebsiella pneumoniae isolates from Singapore. Antimicrob. Agents Chemother. 2003, 47, 2831–2837. [Google Scholar] [CrossRef]
- Ricci, V.; Busby, S.J.W.; Piddock, L.J.V. Regulation of RamA by RamR in Salmonella enterica serovar Typhimurium: Isolation of a RamR superrepressor. Antimicrob. Agents Chemother. 2012, 56, 6037–6040. [Google Scholar] [CrossRef]
- Jiménez-Castellanos, J.-C.; Wan Ahmad Kamil, W.N.I.; Cheung, C.H.P.; Tobin, M.S.; Brown, J.; Isaac, S.G.; Heesom, K.J.; Schneiders, T.; Avison, M.B. Comparative effects of overproducing the AraC-type transcriptional regulators MarA, SoxS, RarA and RamA on antimicrobial drug susceptibility in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2016, 71, 1820–1825. [Google Scholar] [CrossRef]
- Molitor, A.; James, C.E.; Fanning, S.; Pagès, J.-M.; Davin-Regli, A. Ram locus is a key regulator to trigger multidrug resistance in Enterobacter aerogenes. J. Med. Microbiol. 2018, 67, 148–159. [Google Scholar] [CrossRef]
- Fàbrega, A.; Ballesté-Delpierre, C.; Vila, J. Differential impact of ramRA mutations on both ramA transcription and decreased antimicrobial susceptibility in Salmonella Typhimurium. J. Antimicrob. Chemother. 2016, 71, 617–624. [Google Scholar] [CrossRef]
- Belmar Campos, C.; Aepfelbacher, M.; Hentschke, M. Molecular analysis of the ramRA locus in clinical Klebsiella pneumoniae isolates with reduced susceptibility to tigecycline. New Microbiol. 2017, 40, 135–138. [Google Scholar]
- Ricci, V.; Blair, J.M.A.; Piddock, L.J.V. RamA, which controls expression of the MDR efflux pump AcrAB-TolC, is regulated by the Lon protease. J. Antimicrob. Chemother. 2014, 69, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L.J.V. The 2019 Garrod Lecture: MDR efflux in Gram-negative bacteria—How understanding resistance led to a new tool for drug discovery. J. Antimicrob. Chemother. 2019, 74, 3128–3134. [Google Scholar] [CrossRef] [PubMed]
- Ricci, V.; Attah, V.; Overton, T.; Grainger, D.C.; Piddock, L.J.V. CsrA maximizes expression of the AcrAB multidrug resistance transporter. Nucleic Acids Res. 2017, 45, 12798–12807. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-Z.; Chen, P.-X.; Yang, L.; Li, W.; Chang, M.-X.; Jiang, H.-X. Coordinated expression of acrAB-tolC and eight other functional efflux pumps through activating ramA and marA in Salmonella enterica serovar Typhimurium. Microb. Drug Resist. 2017, 24, 120–125. [Google Scholar] [CrossRef]
- Van der Straaten, T.; Zulianello, L.; van Diepen, A.; Granger, D.L.; Janssen, R.; van Dissel, J.T. Salmonella enterica serovar Typhimurium RamA, intracellular oxidative stress response, and bacterial virulence. Infect. Immun. 2004, 72, 996–1003. [Google Scholar] [CrossRef]
- Jiménez-Castellanos, J.-C.; Wan Nur Ismah, W.A.K.; Takebayashi, Y.; Findlay, J.; Schneiders, T.; Heesom, K.J.; Avison, M.B. Envelope proteome changes driven by RamA overproduction in Klebsiella pneumoniae that enhance acquired β-lactam resistance. J. Antimicrob. Chemother. 2018, 73, 88–94. [Google Scholar] [CrossRef]
- Veleba, M.; Higgins, P.G.; Gonzalez, G.; Seifert, H.; Schneiders, T. Characterization of RarA, a novel AraC family multidrug resistance regulator in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2012, 56, 4450–4458. [Google Scholar] [CrossRef]
- Li, J.; Zhang, H.; Ning, J.; Sajid, A.; Cheng, G.; Yuan, Z.; Hao, H. The nature and epidemiology of OqxAB, a multidrug efflux pump. Antimicrob. Resist. Infect. Control. 2019, 8. [Google Scholar] [CrossRef]
- Bialek-Davenet, S.; Lavigne, J.-P.; Guyot, K.; Mayer, N.; Tournebize, R.; Brisse, S.; Leflon-Guibout, V.; Nicolas-Chanoine, M.-H. Differential contribution of AcrAB and OqxAB efflux pumps to multidrug resistance and virulence in Klebsiella pneumoniae. J. Antimicrob. Chemother. 2015, 70, 81–88. [Google Scholar] [CrossRef]
- Lin, Y.-T.; Huang, Y.-W.; Huang, H.-H.; Yang, T.-C.; Wang, F.-D.; Fung, C.-P. In vivo evolution of tigecycline-non-susceptible Klebsiella pneumoniae strains in patients: Relationship between virulence and resistance. Int. J. Antimicrob. Agents 2016, 48, 485–491. [Google Scholar] [CrossRef] [PubMed]
- De Majumdar, S.; Veleba, M.; Finn, S.; Fanning, S.; Schneiders, T. Elucidating the regulon of multidrug resistance regulator RarA in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2013, 57, 1603–1609. [Google Scholar] [CrossRef] [PubMed]
- Griffith, K.L.; Becker, S.M.; Wolf, R.E. Characterization of TetD as a transcriptional activator of a subset of genes of the Escherichia coli SoxS/MarA/Rob regulon. Mol. Microbiol. 2005, 56, 1103–1117. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, H.; Ohe, S.; Ishihama, A. Role of transcription factor NimR (YeaM) in sensitivity control of Escherichia coli to 2-nitroimidazole. FEMS Microbiol. Lett. 2015, 362, 1–8. [Google Scholar] [CrossRef]
- Pomposiello, P.J.; Bennik, M.H.J.; Demple, B. Genome-wide transcriptional profiling of the Escherichia coli responses to superoxide stress and sodium salicylate. J. Bacteriol. 2001, 183, 3890–3902. [Google Scholar] [CrossRef]
- Nishino, K.; Hayashi-Nishino, M.; Yamaguchi, A. H-NS modulates multidrug resistance of Salmonella enterica serovar Typhimurium by repressing multidrug efflux genes acrEF. Antimicrob. Agents Chemother. 2009, 53, 3541–3543. [Google Scholar] [CrossRef]
- Dupont, M.; James, C.E.; Chevalier, J.; Pagès, J.-M. An Early Response to Environmental Stress Involves Regulation of OmpX and OmpF, Two Enterobacterial Outer Membrane Pore-Forming Proteins. Antimicrob. Agents Chemother. 2007, 51, 3190–3198. [Google Scholar] [CrossRef]
- Grkovic, S.; Brown, M.H.; Skurray, R.A. Regulation of bacterial drug export systems. Microbiol. Mol. Biol. Rev. 2002, 66, 671–701. [Google Scholar] [CrossRef]
- Martin, R.G.; Rosner, J.L. Fis, an accessorial factor for transcriptional activation of the mar (multiple antibiotic resistance) promoter of Escherichia coli in the presence of the activator MarA, SoxS, or Rob. J. Bacteriol. 1997, 179, 7410–7419. [Google Scholar] [CrossRef]
- Perez-Rueda, E.; Hernandez-Guerrero, R.; Martinez-Nuñez, M.A.; Armenta-Medina, D.; Sanchez, I.; Ibarra, J.A. Abundance, diversity and domain architecture variability in prokaryotic DNA-binding transcription factors. PLoS ONE 2018, 13. [Google Scholar] [CrossRef]
- Kumar, A.; Schweizer, H.P. Bacterial resistance to antibiotics: Active efflux and reduced uptake. Adv. Drug Deliv. Rev. 2005, 57, 1486–1513. [Google Scholar] [CrossRef] [PubMed]
- Olliver, A.; Vallé, M.; Chaslus-Dancla, E.; Cloeckaert, A. Role of an acrR mutation in multidrug resistance of in vitro-selected fluoroquinolone-resistant mutants of Salmonella enterica serovar Typhimurium. FEMS Microbiol. Lett. 2004, 238, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Pradel, E.; Pagès, J.-M. The AcrAB-TolC efflux pump contributes to multidrug resistance in the nosocomial pathogen Enterobacter aerogenes. Antimicrob. Agents Chemother. 2002, 46, 2640–2643. [Google Scholar] [CrossRef] [PubMed]
- Webber, M.A.; Talukder, A.; Piddock, L.J.V. Contribution of mutation at amino acid 45 of AcrR to acrB expression and ciprofloxacin resistance in clinical and veterinary Escherichia coli isolates. Antimicrob. Agents Chemother. 2005, 49, 4390–4392. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Chen, C.-C. Computational analysis of the molecular mechanism of RamR mutations contributing to antimicrobial resistance in Salmonella enterica. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Yamasaki, S.; Nakashima, R.; Sakurai, K.; Baucheron, S.; Giraud, E.; Doublet, B.; Cloeckaert, A.; Nishino, K. Crystal structure of the multidrug resistance regulator RamR complexed with bile acids. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Chiu, S.-K.; Huang, L.-Y.; Chen, H.; Tsai, Y.-K.; Liou, C.-H.; Lin, J.-C.; Siu, L.K.; Chang, F.-Y.; Yeh, K.-M. Roles of ramR and tet(A) mutations in conferring tigecycline resistance in carbapenem-resistant Klebsiella pneumoniae clinical isolates. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef]
- Liu, H.; Jia, X.; Zou, H.; Sun, S.; Li, S.; Wang, Y.; Xia, Y. Detection and characterization of tigecycline heteroresistance in E. cloacae: Clinical and microbiological findings. Emerg. Microbes Infect. 2019, 8, 564–574. [Google Scholar] [CrossRef]
- Wang, X.; Chen, H.; Zhang, Y.; Wang, Q.; Zhao, C.; Li, H.; He, W.; Zhang, F.; Wang, Z.; Li, S.; et al. Genetic characterisation of clinical Klebsiella pneumoniae isolates with reduced susceptibility to tigecycline: Role of the global regulator RamA and its local repressor RamR. Int. J. Antimicrob. Agents 2015, 45, 635–640. [Google Scholar] [CrossRef]
- Matange, N. Highly contingent phenotypes of Lon protease deficiency in Escherichia coli upon antibiotic challenge. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef]
- Nové, M.; Kincses, A.; Molnar, J.; Amaral, L.; Spengler, G. The role of efflux pumps and environmental pH in bacterial multidrug resistance. In Vivo 2020, 34, 65–71. [Google Scholar] [CrossRef]
- Dam, S.; Pagès, J.-M.; Masi, M. Stress responses, outer membrane permeability control and antimicrobial resistance in Enterobacteriaceae. Microbiology 2018, 164, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Cardona, S.T.; Choy, M.; Hogan, A.M. Essential two-component systems regulating cell envelope functions: Opportunities for novel antibiotic therapies. J. Membr. Biol. 2018, 251, 75–89. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, R.; Ferenci, T. The fitness costs and benefits of antibiotic resistance in drug-free microenvironments encountered in the human body. Environ. Microbiol. Rep. 2017, 9, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Felden, B.; Cattoir, V. Bacterial adaptation to antibiotics through regulatory RNAs. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed]
- Filippova, E.V.; Zemaitaitis, B.; Aung, T.; Wolfe, A.J.; Anderson, W.F. Structural basis for DNA recognition by the two-component response regulator RcsB. mBio 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Dersch, P.; Khan, M.A.; Mühlen, S.; Görke, B. Roles of regulatory RNAs for antibiotic resistance in bacteria and their potential value as novel drug targets. Front. Microbiol. 2017, 8, 803. [Google Scholar] [CrossRef]
- Raivio, T.L.; Leblanc, S.K.D.; Price, N.L. The Escherichia coli Cpx envelope stress response regulates genes of diverse function that impact antibiotic resistance and membrane integrity. J. Bacteriol. 2013, 195, 2755–2767. [Google Scholar] [CrossRef]
- Laubacher, M.E.; Ades, S.E. The Rcs phosphorelay is a cell envelope stress response activated by peptidoglycan stress and contributes to intrinsic antibiotic resistance. J. Bacteriol. 2008, 190, 2065–2074. [Google Scholar] [CrossRef]
- Ruiz, N.; Silhavy, T.J. Sensing external stress: Watchdogs of the Escherichia coli cell envelope. Curr. Opin. Microbiol. 2005, 8, 122–126. [Google Scholar] [CrossRef]
- Masi, M.; Pinet, E.; Pagès, J.-M. Complex response of the CpxAR two-component system to β-lactams on antibiotic resistance and envelop homeostasis in Enterobacteriaceae. Antimicrob. Agents Chemother. 2020. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Costa, R.D.; Das, M.; Nandi, D. Interplay of cold shock protein E with an uncharacterized protein, YciF, lowers porin expression and enhances bile resistance in Salmonella Typhimurium. J. Biol. Chem. 2019, 294, 9084–9099. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Jamal, S.B.; Hassan, S.S.; Carvalho, P.V.S.D.; Almeida, S.; Barh, D.; Ghosh, P.; Silva, A.; Castro, T.L.P.; Azevedo, V. Two-component signal transduction systems of pathogenic bacteria as targets for antimicrobial therapy: An overview. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Tierney, A.R.; Rather, P.N. Roles of two-component regulatory systems in antibiotic resistance. Future Microbiol. 2019, 14, 533–552. [Google Scholar] [CrossRef] [PubMed]
- Singer, A.C.; Shaw, H.; Rhodes, V.; Hart, A. Review of antimicrobial resistance in the environment and its relevance to environmental regulators. Front. Microbiol 2016, 7. [Google Scholar] [CrossRef]
- Klein, G.; Raina, S. Regulated assembly of LPS, its structural alterations and cellular response to LPS defects. Int. J. Mol. Sci. 2019, 20, 356. [Google Scholar] [CrossRef]
- Delhaye, A.; Collet, J.-F.; Laloux, G. Fine-tuning of the Cpx envelope stress response is required for cell wall homeostasis in Escherichia coli. mBio 2016, 7. [Google Scholar] [CrossRef]
- Delhaye, A.; Collet, J.-F.; Laloux, G. A Fly on the wall: How stress response systems can sense and respond to damage to peptidoglycan. Front. Cell Infect. Microbiol. 2019, 9. [Google Scholar] [CrossRef]
- Hews, C.L.; Cho, T.; Rowley, G.; Raivio, T.L. Maintaining integrity under stress: Envelope stress response regulation of pathogenesis in Gram-negative bacteria. Front. Cell Infect. Microbiol. 2019, 9. [Google Scholar] [CrossRef]
- Cho, H.; Uehara, T.; Bernhardt, T.G. Beta-lactam antibiotics induce a lethal malfunctioning of the bacterial cell wall synthesis machinery. Cell 2014, 159, 1300–1311. [Google Scholar] [CrossRef]
- Aghapour, Z.; Gholizadeh, P.; Ganbarov, K.; Bialvaei, A.Z.; Mahmood, S.S.; Tanomand, A.; Yousefi, M.; Asgharzadeh, M.; Yousefi, B.; Kafil, H.S. Molecular mechanisms related to colistin resistance in Enterobacteriaceae. Infect. Drug Resist. 2019, 12, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.S.; Chen, H.-W.; Zhang, R.-Y.; Huang, C.-Y.; Shen, C.-F. The expression levels of outer membrane proteins STM1530 and OmpD, which are influenced by the CpxAR and BaeSR two-component systems, play important roles in the ceftriaxone resistance of Salmonella enterica serovar Typhimurium. Antimicrob. Agents Chemother. 2011, 55, 3829–3837. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Sun, Y.; Yuan, L.; Pan, Y.; Gao, Y.; Ma, C.; Hu, G. Regulation of the two-component regulator CpxR on aminoglycosides and β-lactams resistance in Salmonella enterica serovar Typhimurium. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Klobucar, K.; French, S.; Côté, J.-P.; Howes, J.R.; Brown, E.D. Genetic and chemical-genetic interactions map biogenesis and permeability determinants of the outer membrane of Escherichia coli. mBio 2020, 11. [Google Scholar] [CrossRef]
- Knopp, M.; Andersson, D.I. Amelioration of the fitness costs of antibiotic resistance due to reduced outer membrane permeability by upregulation of alternative porins. Mol. Biol. Evol. 2015, 32, 3252–3263. [Google Scholar] [CrossRef]
- Anandan, A.; Vrielink, A. Structure and function of lipid A–modifying enzymes. Ann. New York Acad. Sci. 2020, 1459, 19–37. [Google Scholar] [CrossRef]
- Pagnout, C.; Sohm, B.; Razafitianamaharavo, A.; Caillet, C.; Offroy, M.; Leduc, M.; Gendre, H.; Jomini, S.; Beaussart, A.; Bauda, P.; et al. Pleiotropic effects of rfa-gene mutations on Escherichia coli envelope properties. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef]
- Rome, K.; Borde, C.; Taher, R.; Cayron, J.; Lesterlin, C.; Gueguen, E.; De Rosny, E.; Rodrigue, A. The two-component system ZraPSR is a novel ESR that contributes to intrinsic antibiotic tolerance in Escherichia coli. J. Mol. Biol. 2018, 430, 4971–4985. [Google Scholar] [CrossRef]
- Dean, C.R.; Barkan, D.T.; Bermingham, A.; Blais, J.; Casey, F.; Casarez, A.; Colvin, R.; Fuller, J.; Jones, A.K.; Li, C.; et al. Mode of action of the monobactam LYS228 and mechanisms decreasing in vitro susceptibility in Escherichia coli and Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef]
- Philippe, N.; Maigre, L.; Santini, S.; Pinet, E.; Claverie, J.-M.; Davin-Régli, A.-V.; Pagès, J.-M.; Masi, M. In vivo evolution of bacterial resistance in two cases of Enterobacter aerogenes infections during treatment with imipenem. PLoS ONE 2015, 10, e0138828. [Google Scholar] [CrossRef]
- Wang, H.; Huang, M.; Zeng, X.; Peng, B.; Xu, X.; Zhou, G. Resistance profiles of Salmonella isolates exposed to stresses and the expression of small non-coding RNAs. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Diallo, I.; Provost, P. RNA-sequencing analyses of small bacterial RNAs and their emergence as virulence factors in host-pathogen interactions. Int. J. Mol. Sci. 2020, 21, 1627. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, K.S.; Gottesman, S. Small regulatory RNAs in the enterobacterial response to envelope damage and oxidative stress. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Van Puyvelde, S.; Vanderleyden, J.; De Keersmaecker, S.C.J. Experimental approaches to identify small RNAs and their diverse roles in bacteria – what we have learnt in one decade of MicA research. Microbiologyopen 2015, 4, 699–711. [Google Scholar] [CrossRef] [PubMed]
- Kröger, C.; Colgan, A.; Srikumar, S.; Händler, K.; Sivasankaran, S.K.; Hammarlöf, D.L.; Canals, R.; Grissom, J.E.; Conway, T.; Hokamp, K.; et al. An infection-relevant transcriptomic compendium for Salmonella enterica serovar Typhimurium. Cell Host Microbe 2013, 14, 683–695. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Takada, H.; Yamamoto, K.; Ishihama, A. Expanded roles of two-component response regulator OmpR in Escherichia coli: Genomic SELEX search for novel regulation targets. Genes Cells 2015, 20, 915–931. [Google Scholar] [CrossRef]
- Alvarado, A.; Behrens, W.; Josenhans, C. Protein activity sensing in bacteria in regulating metabolism and motility. Front. Microbiol. 2020, 10. [Google Scholar] [CrossRef]
- Henly, E.L.; Dowling, J.A.R.; Maingay, J.B.; Lacey, M.M.; Smith, T.J.; Forbes, S. Biocide exposure induces changes in susceptibility, pathogenicity, and biofilm formation in uropathogenic Escherichia coli. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- Lu, J.; Jin, M.; Nguyen, S.H.; Mao, L.; Li, J.; Coin, L.J.M.; Yuan, Z.; Guo, J. Non-antibiotic antimicrobial triclosan induces multiple antibiotic resistance through genetic mutation. Environ. Int. 2018, 118, 257–265. [Google Scholar] [CrossRef]
- Moore, J.P.; Li, H.; Engmann, M.L.; Bischof, K.M.; Kunka, K.S.; Harris, M.E.; Tancredi, A.C.; Ditmars, F.S.; Basting, P.J.; George, N.S.; et al. Inverted regulation of multidrug efflux pumps, acid resistance, and porins in benzoate-evolved Escherichia coli K-12. Appl. Environ. Microbiol. 2019, 85. [Google Scholar] [CrossRef]
- Jin, M.; Lu, J.; Chen, Z.; Nguyen, S.H.; Mao, L.; Li, J.; Yuan, Z.; Guo, J. Antidepressant fluoxetine induces multiple antibiotics resistance in Escherichia coli via ROS-mediated mutagenesis. Environ. Int. 2018, 120, 421–430. [Google Scholar] [CrossRef] [PubMed]
- Balagué, C.; Véscovi, E.G. Activation of multiple antibiotic resistance in uropathogenic Escherichia coli strains by aryloxoalcanoic acid compounds. Antimicrob. Agents Chemother. 2001, 45, 1815–1822. [Google Scholar] [CrossRef] [PubMed]
- Masi, M.; Réfrégiers, M.; Pos, K.M.; Pagès, J.-M. Mechanisms of envelope permeability and antibiotic influx and efflux in Gram-negative bacteria. Nat. Microbiol. 2017, 2, 17001. [Google Scholar] [CrossRef] [PubMed]
- Burby, P.E.; Simmons, L.A. Regulation of cell division in bacteria by monitoring genome integrity and DNA replication status. J. Bacteriol. 2020, 202. [Google Scholar] [CrossRef] [PubMed]
- Harms, A.; Maisonneuve, E.; Gerdes, K. Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 2016, 354. [Google Scholar] [CrossRef]
- Khatoon, A.; Malik, H.M.T.; Aurongzeb, M.; Raza, S.A.; Karim, A. Draft genome of a macrolide resistant XDR Salmonella enterica serovar Paratyphi A strain using a shotgun sequencing approach. J. Glob. Antimicrob. Resist. 2019, 19, 129–131. [Google Scholar] [CrossRef]
- Naha, S.; Sands, K.; Mukherjee, S.; Roy, C.; Rameez, M.J.; Saha, B.; Dutta, S.; Walsh, T.R.; Basu, S. KPC-2-producing Klebsiella pneumoniae ST147 in a neonatal unit: Clonal isolates with differences in colistin susceptibility attributed to AcrAB-TolC pump. Int. J. Antimicrob. Agents 2020, 55, 105903. [Google Scholar] [CrossRef]
- Gandra, S.; Choi, J.; McElvania, E.; Green, S.J.; Harazin, M.; Thomson, R.B.; Dantas, G.; Singh, K.S.; Das, S. Faropenem resistance causes in vitro cross-resistance to carbapenems in ESBL-producing Escherichia coli. Int. J. Antimicrob. Agents 2020, 55, 105902. [Google Scholar] [CrossRef]
- Blázquez, J.; Rodríguez-Beltrán, J.; Matic, I. Antibiotic-induced genetic variation: How it arises and how it can be prevented. Annu. Rev. Microbiol. 2018, 72, 209–230. [Google Scholar] [CrossRef]
- Mishra, M.; Panda, S.; Barik, S.; Sarkar, A.; Singh, D.V.; Mohapatra, H. Antibiotic resistance profile, outer membrane proteins, virulence factors and genome sequence analysis reveal clinical isolates of Enterobacter are potential pathogens compared to environmental isolates. Front. Cell Infect. Microbiol. 2020, 10. [Google Scholar] [CrossRef]
- Vashishtha, K.; Mahadevan, S. Catabolism of aromatic β-glucosides by bacteria can lead to antibiotics resistance. Arch. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bhagirath, A.Y.; Li, Y.; Patidar, R.; Yerex, K.; Ma, X.; Kumar, A.; Duan, K. Two Component regulatory systems and antibiotic resistance in Gram-negative pathogens. Int. J. Mol. Sci. 2019, 20, 1781. [Google Scholar] [CrossRef] [PubMed]
- El-Mowafi, S.A.; Alumasa, J.N.; Ades, S.E.; Keiler, K.C. Cell-based assay to identify inhibitors of the Hfq-sRNA regulatory pathway. Antimicrob. Agents Chemother. 2014, 58, 5500–5509. [Google Scholar] [CrossRef]
- Bellio, P.; Mancini, A.; Di Pietro, L.; Cracchiolo, S.; Franceschini, N.; Reale, S.; de Angelis, F.; Perilli, M.; Amicosante, G.; Spyrakis, F.; et al. Inhibition of the transcriptional repressor LexA: Withstanding drug resistance by inhibiting the bacterial mechanisms of adaptation to antimicrobials. Life Sci. 2020, 241, 117116. [Google Scholar] [CrossRef] [PubMed]
- Barrett, T.C.; Mok, W.W.K.; Murawski, A.M.; Brynildsen, M.P. Enhanced antibiotic resistance development from fluoroquinolone persisters after a single exposure to antibiotic. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Recacha, E.; Machuca, J.; Díaz-Díaz, S.; García-Duque, A.; Ramos-Guelfo, M.; Docobo-Pérez, F.; Blázquez, J.; Pascual, A.; Rodríguez-Martínez, J.M. Suppression of the SOS response modifies spatiotemporal evolution, post-antibiotic effect, bacterial fitness and biofilm formation in quinolone-resistant Escherichia coli. J. Antimicrob. Chemother. 2019, 74, 66–73. [Google Scholar] [CrossRef]


© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferrand, A.; Vergalli, J.; Pagès, J.-M.; Davin-Regli, A. An Intertwined Network of Regulation Controls Membrane Permeability Including Drug Influx and Efflux in Enterobacteriaceae. Microorganisms 2020, 8, 833. https://doi.org/10.3390/microorganisms8060833
Ferrand A, Vergalli J, Pagès J-M, Davin-Regli A. An Intertwined Network of Regulation Controls Membrane Permeability Including Drug Influx and Efflux in Enterobacteriaceae. Microorganisms. 2020; 8(6):833. https://doi.org/10.3390/microorganisms8060833
Chicago/Turabian StyleFerrand, Aurélie, Julia Vergalli, Jean-Marie Pagès, and Anne Davin-Regli. 2020. "An Intertwined Network of Regulation Controls Membrane Permeability Including Drug Influx and Efflux in Enterobacteriaceae" Microorganisms 8, no. 6: 833. https://doi.org/10.3390/microorganisms8060833
APA StyleFerrand, A., Vergalli, J., Pagès, J.-M., & Davin-Regli, A. (2020). An Intertwined Network of Regulation Controls Membrane Permeability Including Drug Influx and Efflux in Enterobacteriaceae. Microorganisms, 8(6), 833. https://doi.org/10.3390/microorganisms8060833