Dual-Purpose Inoculants and Their Effects on Corn Silage

 and
and
Abstract
1. Introduction
2. Methods and Materials
2.1. Corn Silage Collection as Isolate Source
2.2. Lactic Acid Bacteria Isolation
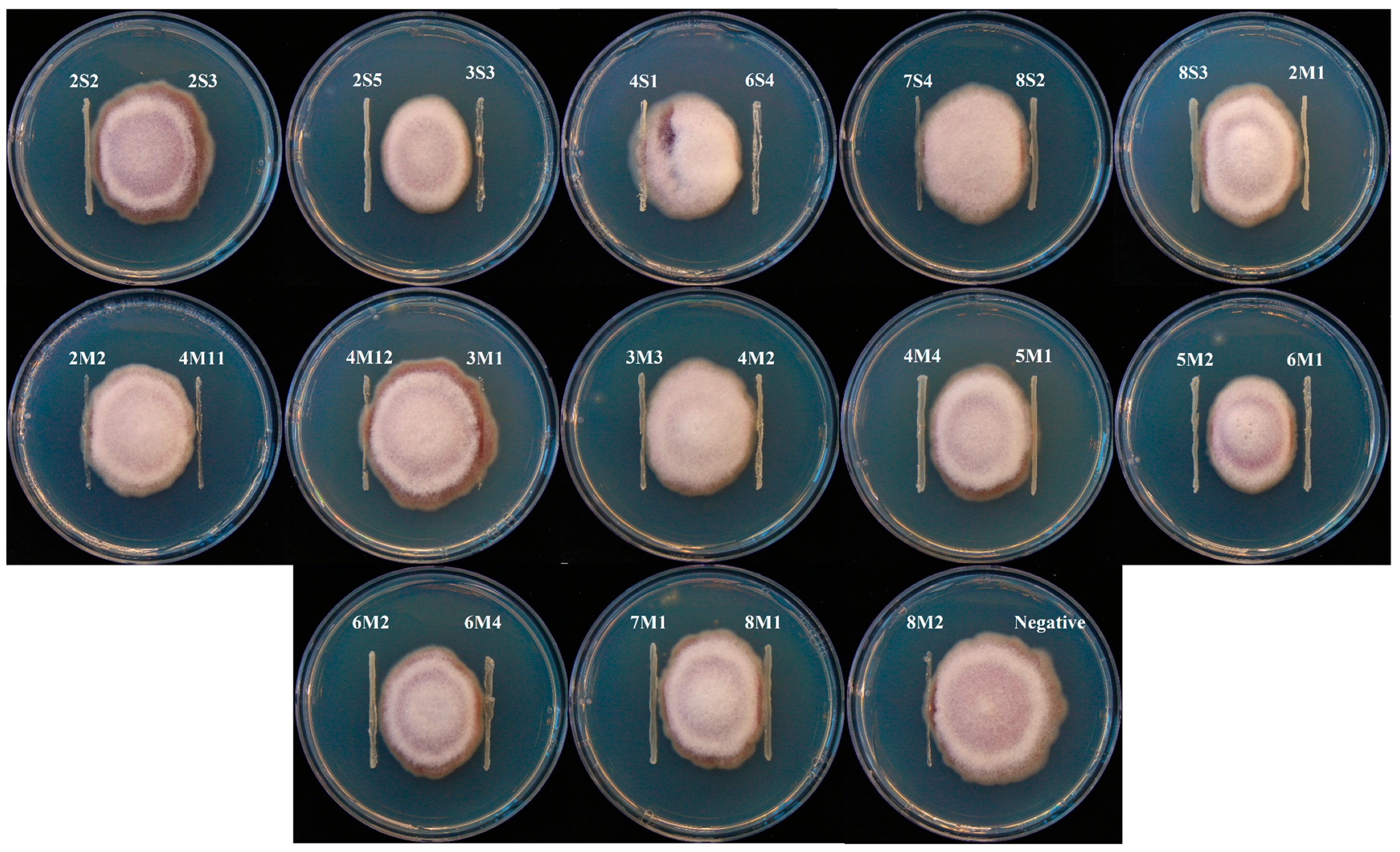
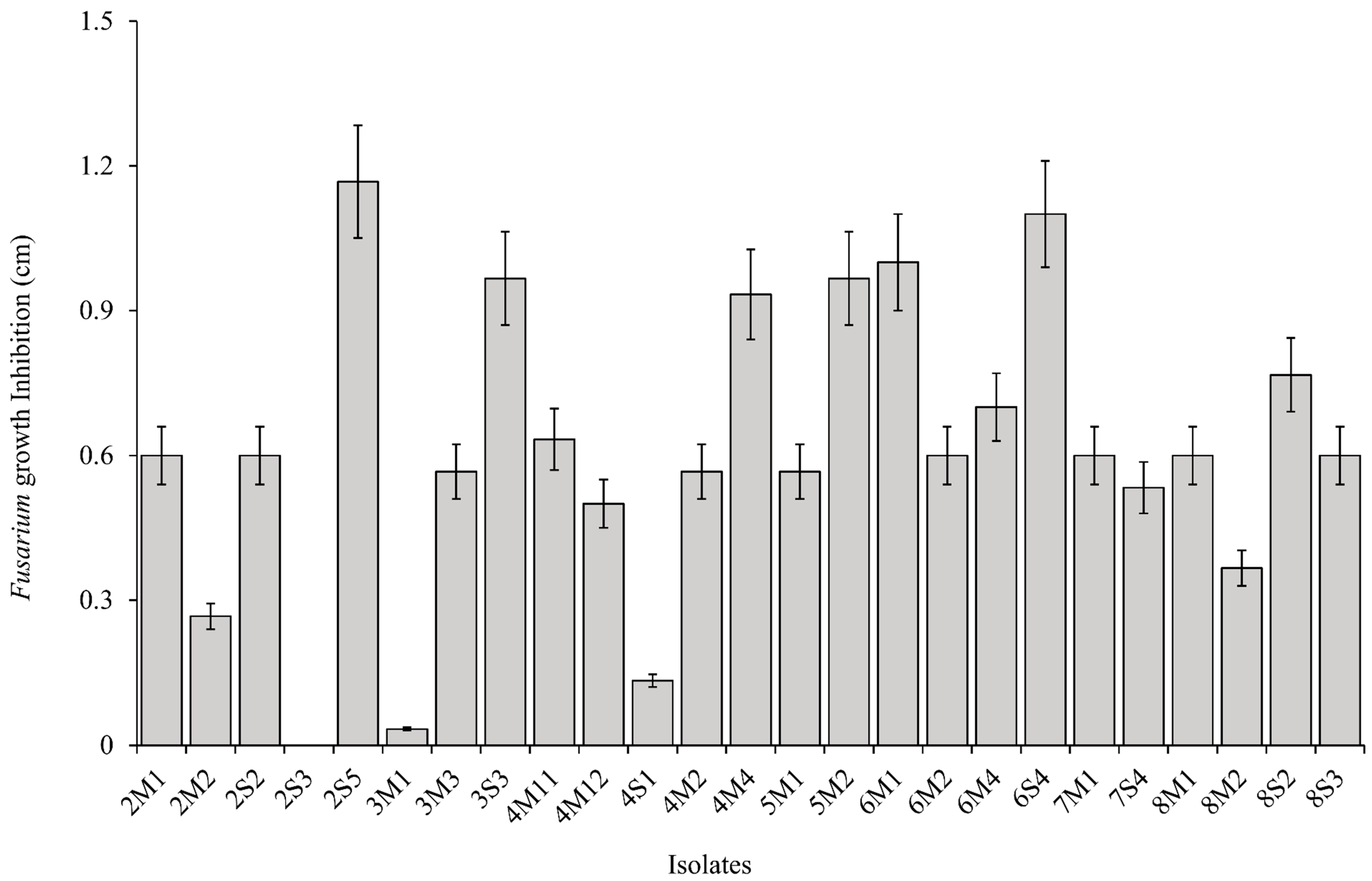
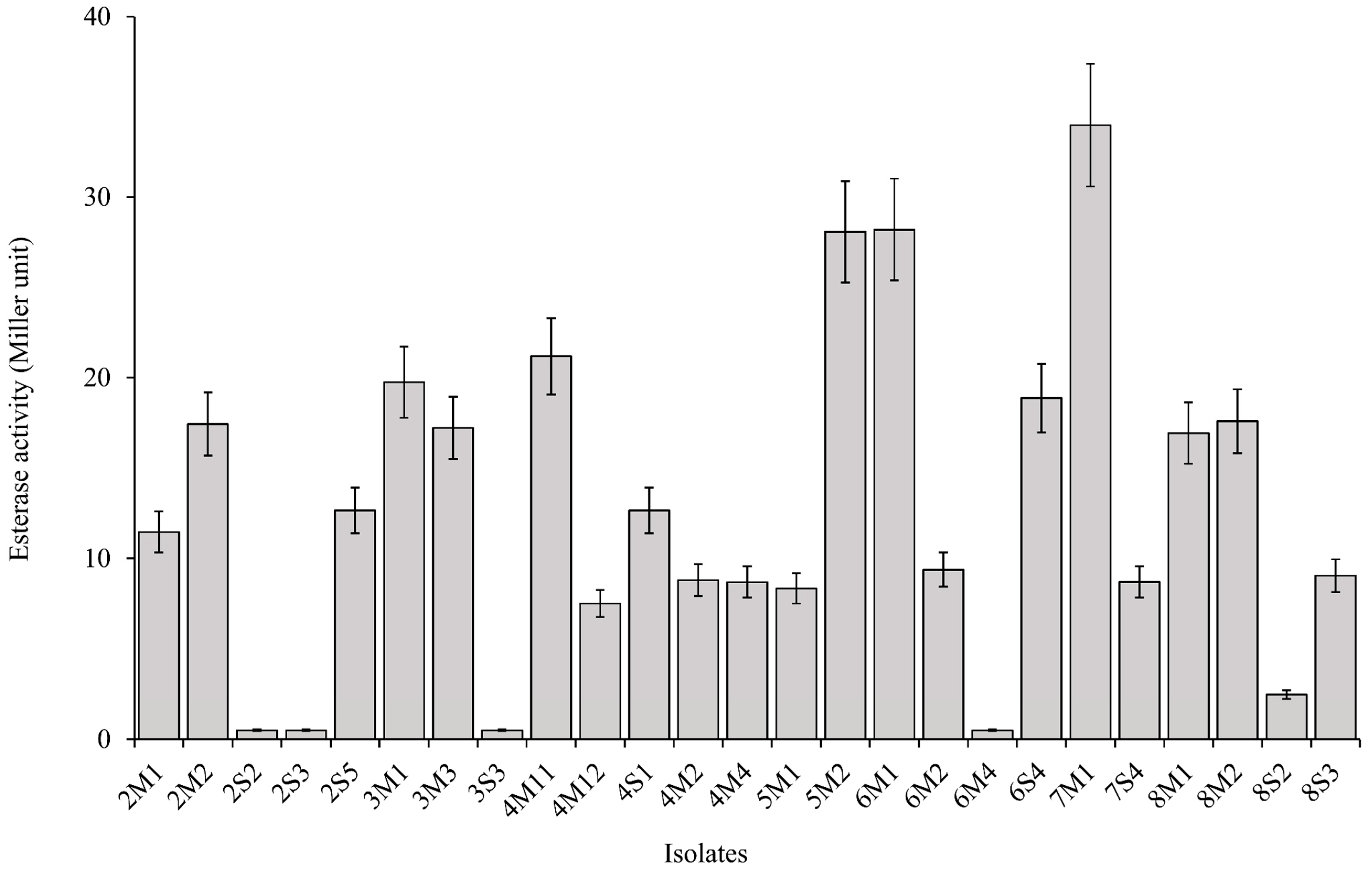
2.3. Antifungal and Carboxylesterase Activities
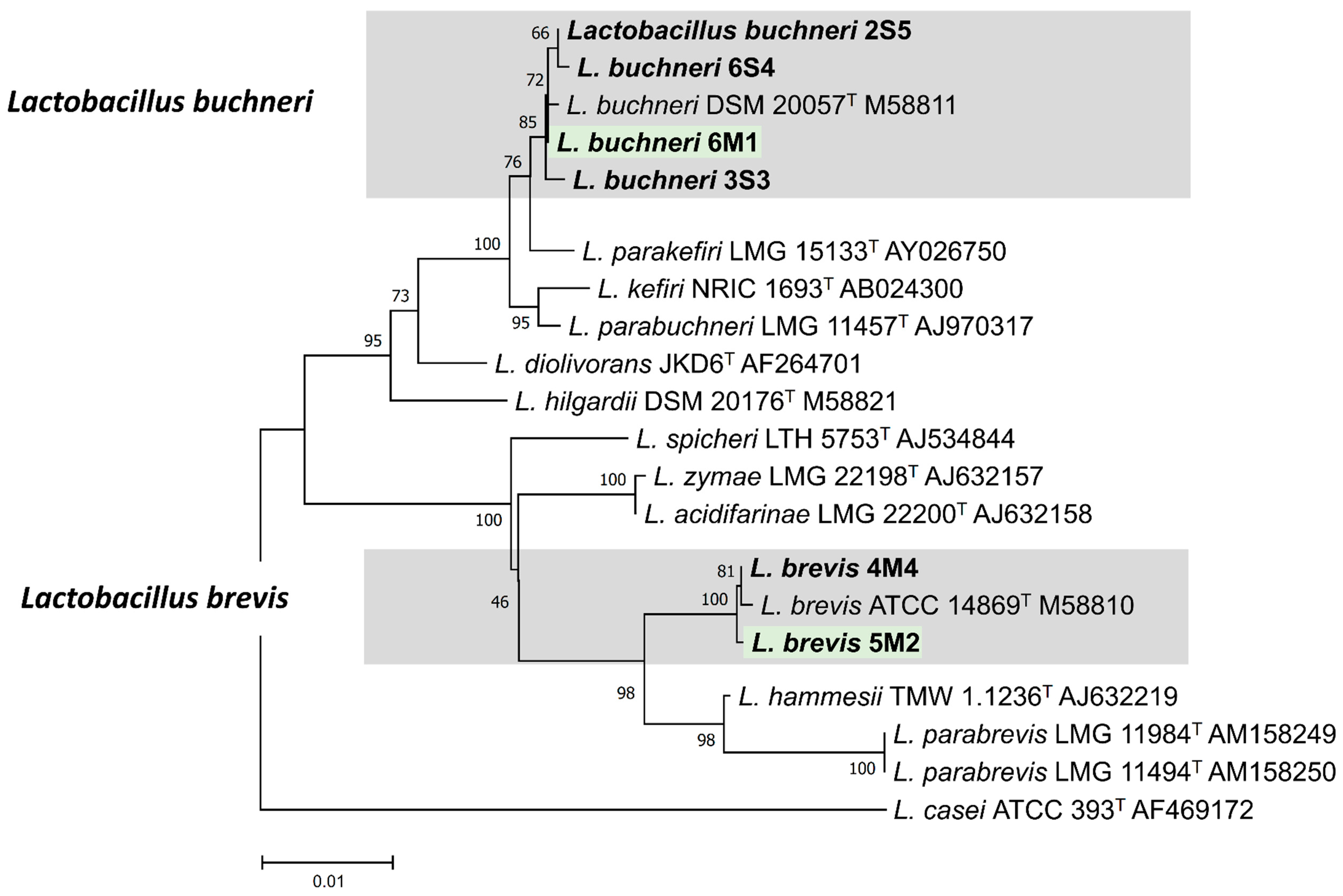
2.4. Identification Using the 16S rRNA Gene Sequencing
2.5. Inoculant Preparation
2.6. Silage Production
2.7. Chemical Compositions and In Vitro Rumen Digestibility
2.8. Fermentation Characteristics and Microbial Counts
2.9. Aerobic Stability
2.10. Statistical Analysis
3. Results
3.1. General Information of Collected Corn Silages as Isolate Source
3.2. Antifungal and Carboxylesterase Activities of Isolates
3.3. Identification of Isolates Using the 16S rRNA Gene Sequencing
3.4. Chemical Compositions and In Vitro Digestibility of Forage and Silage
3.5. Fermentation Characteristics and Microbial Counts of Silage
3.6. Aerobic Stability of Silage
4. Discussion
4.1. Screening Isolate
4.2. Applying Screened Isolates on Corn Silage
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Alonso, V.A.; Pereyra, C.M.; Keller, L.A.M.; Calcero, A.M.; Rosa, C.A.R.; Chiacchiera, S.M.; Cavaglieri, L.R. Fungi and mycotoxins in silage: an overview. J. Appl. Microbiol. 2013, 115, 637–643. [Google Scholar] [CrossRef]
- Eckard, S.; Wettstein, F.E.; Forrer, H.-R.; Vogelgsang, S. Incidence of Fusarium species and mycotoxins in silage maize. Toxins 2011, 3, 949–967. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, J.M.; Davies, D.R. The aerobic stability of silage: Key findings and recent developments. Grass Forage Sci. 2012, 68, 1–19. [Google Scholar] [CrossRef]
- McDonald, P.; Henderson, A.R.; Heron, S.J.E. The Biochemistry of Silage, 2nd ed.; Chalcombe Publ: Marlow, Bucks, UK, 1991; pp. 48–291. [Google Scholar]
- Kleinschmit, D.H.; Schmidt, R.J.; Kung, L., Jr. The effects of various antifungal additives on the fermentation and aerobic stability of corn silage. J. Dairy Sci. 2005, 88, 2130–2139. [Google Scholar] [CrossRef]
- Danner, H.; Holzer, M.; Mayrhuber, E.; Braun, R. Acetic acid increases stability of silage under aerobic conditions. Appl. Environ. Microbiol. 2003, 69, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Schnürer, J.; Magnusson, J. Antifungal lactic acid bacteria as biopreservatives. Trends Food Sci. Technol. 2005, 16, 70–78. [Google Scholar] [CrossRef]
- Hatfield, R.D.; Jung, H.J.G.; Broderick, G.; Jenkins, T.C. Nutritional chemistry of forages. In Forages: The Science of Grassland Agriculture; Barnes, R.F., Nelson, C.J., Moore, K.J., Collins, M., Eds.; Blackwell Publishing: Ames, IA, USA, 2007; pp. 467–485. [Google Scholar]
- Adesogan, A.T.; Ma, Z.X.; Romero, J.J.; Arriola, K.G. Ruminant Nutrition Symposium: Improving cell wall digestion and animal performance with fibrolytic enzymes. J. Anim. Sci. 2014, 92, 1317–1330. [Google Scholar] [CrossRef]
- Romero, J.J.; Zarate, M.A.; Adesogan, A.T. Effect of the dose of exogenous fibrolytic enzyme preparations on preingestive fiber hydrolysis, ruminal fermentation, and in vitro digestibility of bermudagrass haylage. J. Dairy Sci. 2015, 98, 406–417. [Google Scholar] [CrossRef]
- Kang, T.W.; Adesogan, A.T.; Kim, S.C.; Lee, S.S. Effects of an esterase-producing inoculant on fermentation, aerobic stability, and neutral detergent fiber digestibility of corn silage. J. Dairy Sci. 2009, 92, 732–738. [Google Scholar] [CrossRef]
- Lynch, J.P.; Lin, L.; Lara, E.C.; Baah, J.; Beauchemin, K.A. The effect of exogenous fibrinolytic enzymes and a ferulic acid esterase-producing inoculant on the fiber degradability, chemical composition and conservation characteristics of alfalfa silage. Anim. Feed Sci. Technol. 2014, 193, 21–31. [Google Scholar] [CrossRef]
- Paradhipta, D.H.V.; Joo, Y.H.; Lee, H.J.; Lee, S.S.; Kwak, Y.S.; Han, O.K.; Kim, D.H.; Kim, S.C. Effects of wild or mutated inoculant on rye silage and its rumen fermentation indices. Asian-Australas. J. Anim. Sci. 2020, 33, 949–956. [Google Scholar] [CrossRef]
- Gardner, N.J.; Savard, T.; Obermeier, P.; Caldwll, G.; Champagne, C.P. Selection and characterization of mixed starter cultures for lactic acid fermentation of carrot, cabbage, beet, and onion vegetable mixtures. Int. J. Food Microbiol. 2001, 64, 261–275. [Google Scholar] [CrossRef]
- Giraffa, G.; Chanishvili, N.; Widyastuti, Y. Importance of lactobacilli in food and feed biotechnology. Res. Microbiol. 2010, 161, 480–487. [Google Scholar] [CrossRef] [PubMed]
- Muck, R.E.; Nadeau, E.M.G.; McAllister, T.A.; Contreras-Govea, F.E.; Santos, M.C.; Kung, L., Jr. Silage review: Recent advances and future uses of silage additives. J. Dairy Sci. 2018, 101, 3980–4000. [Google Scholar] [CrossRef] [PubMed]
- Arriola, K.G.; Kim, S.C.; Huisden, C.M.; Adesogan, A.T. Stay-green ranking and maturity of corn hybrids:1. Effects on dry matter yield, nutritional value, fermentation characteristics, and aerobic stability of silage hybrids in Florida. J. Dairy Sci. 2012, 95, 964–974. [Google Scholar] [CrossRef] [PubMed]
- Flieg, O. A key for evaluation of silage samples. Futterb Garfutterber 1983, 1, 112–128. [Google Scholar]
- Pérez-Martín, F.; Seseña, S.; Izquierdo, P.M.; Palop, M.L. Esterase activity of lactic acid bacteria isolated from malolactic fermentation of red wines. Int. J. Food Microbiol. 2013, 163, 2–3. [Google Scholar] [CrossRef]
- Dawson, R.M.C.; Elliott, D.C.; Elliot, W.H.; Jones, K.M. Data for Biochemical Research, 3rd ed.; Clarendon Press: Oxford, UK, 1986; pp. 356–357. [Google Scholar]
- Miller, J.H. Experiments in Molecular Genetics; Cold Spring Harbor: Suffolk, NY, USA, 1972; pp. 352–355. [Google Scholar]
- Basic Local Alignment Search Tool: US National Library of Medicine. Available online: http://www.ncbi.nlm.nih.gov/blast/ (accessed on 31 July 2018).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Bio. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis, 18th ed.; AOAC International: Washington DC, USA, 2005. [Google Scholar]
- Tilley, J.M.A.; Terry, R.A. A two-stage technique for the in vitro digestion of forage crops. J. Br. Grassl. Soc. 1963, 18, 104–111. [Google Scholar] [CrossRef]
- Chaney, A.L.; Marbach, E.P. Modified reagents for determination of urea and ammonia. Clin. Chem. 1962, 8, 130–132. [Google Scholar] [CrossRef]
- Muck, R.E.; Dickerson, J.T. Storage temperature effects on proteolysis in alfalfa silage. Trans. ASASE 1988, 31, 1005–1009. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT User’s Guide, Version 9; SAS Institute Inc.: Cary, NC, USA, 2002. [Google Scholar]
- Kung, L., Jr.; Shaver, R.D.; Grant, R.J.; Schmidt, R.J. Silage review: Interpretation of chemical, microbial, and organoleptic components of silage. J. Dairy Sci. 2018, 101, 4020–4033. [Google Scholar] [CrossRef] [PubMed]
- Scudamore, K.A.; Livesey, C.T. Occurrence and significance of mycotoxins in forage crops and silage: A review. J. Sci. Food Agric. 1998, 77, 1–17. [Google Scholar] [CrossRef]
- Lee, J.; Kim, H.; Jeon, J.J.; Kim, H.S.; Zeller, K.A.; Carter, L.L.A.; Leslie, J.F.; Lee, Y.W. Population structure of and mycotoxin production by Fusarium graminearum from maize in South Korea. J. Appl. Environ. Microbiol. 2012, 78, 2161–2167. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, G.O.; Gruninger, R.J.; Badhan, A.; McAllister, T.A. Mining the rumen for fibrolytic feed enzymes. Anim. Front. 2016, 6, 20–26. [Google Scholar] [CrossRef]
- Lee, S.S.; Lee, H.J.; Paradhipta, D.H.V.; Joo, Y.H.; Kim, S.B.; Kim, D.H.; Kim, S.C. Temperature and microbial changes of corn silage during aerobic exposure. Asian-Australas. J. Anim. Sci. 2019, 32, 988–995. [Google Scholar] [CrossRef]
- Stokes, M.R.; Chen, J. Effects of an enzyme-inoculant mixture on the course of fermentation of corn silage. J. Dairy Sci. 1994, 77, 3401–3409. [Google Scholar] [CrossRef]
- Oba, M.; Allen, M.S. Evaluation of the importance of the digestibility of neutral detergent fiber from the forage: Effects on dry matter intake and milk yield of dairy cows. J. Dairy Sci. 1999, 82, 589–596. [Google Scholar] [CrossRef]
- Zheng, J.; Wuttouck, S.; Salvetti, E.; Franz, C.M.A.P.; Haris, H.M.B.; Mattarelli, P.; O’toole, P.W.; Pot, B.; Vandamme, P.; Walter, J.; et al. A taxonomic note on the genus Lactobacillus: Description of 23 of novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. Int. J. Syst. Evol. Microbiol. 2020, 70, 2782–2858. [Google Scholar] [CrossRef]
- Paradhipta, D.H.V.; Joo, Y.H.; Lee, H.J.; Lee, S.S.; Kim, D.H.; Kim, J.D.; Kim, S.C. Effects of inoculant application on fermentation quality and rumen digestibility of high moisture sorghum-sudangrass silage. J. Appl. Anim. Res. 2019, 47, 486–491. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
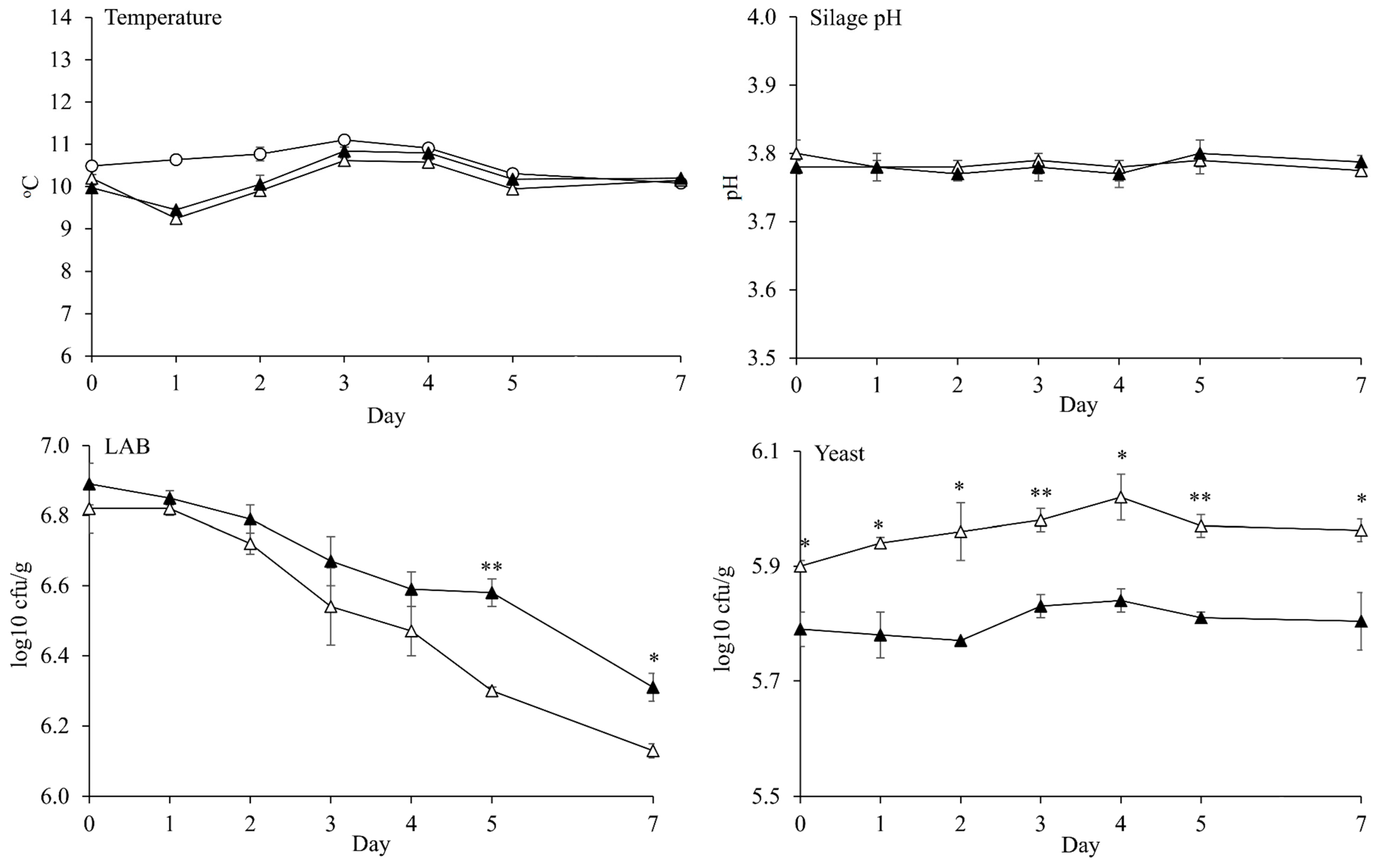{kind=link}
| Item | Source | |||||||
|---|---|---|---|---|---|---|---|---|
| Farm1 | Farm2 | Farm3 | Farm4 | Farm5 | Farm6 | Farm7 | Farm8 | |
| Dry matter, g/kg | 230 | 328 | 370 | 264 | 363 | 281 | 244 | 256 |
| Fermentation characteristics | ||||||||
| pH | 3.67 | 4.09 | 3.94 | 3.65 | 3.89 | 3.68 | 3.74 | 4.55 |
| Ammonia-N, g/kg DM | 0.92 | 0.64 | 0.80 | 0.45 | 0.70 | 0.53 | 0.56 | 1.29 |
| Lactate, g/kg DM | 96.1 | 29.6 | 55.8 | 48.0 | 51.7 | 62.5 | 50.6 | 17.5 |
| Acetate, g/kg DM | 16.7 | 8.08 | 4.57 | 10.8 | 5.51 | 4.82 | 8.98 | 12.0 |
| Propionate, g/kg DM | ND2 | ND | ND | ND | ND | ND | ND | 1.21 |
| Butyrate, g/kg DM | ND | ND | ND | ND | ND | ND | ND | ND |
| Flieg’s Score a | 100 | 94 | 100 | 98 | 100 | 100 | 100 | 78 |
| Microbial count, log10 cfu/g | ||||||||
| Lactic acid bacteria | 6.59 | 7.22 | 6.15 | 6.30 | 7.15 | 6.00 | 7.45 | 6.40 |
| Yeast | 6.00 | 7.13 | 5.70 | 5.65 | 6.39 | 5.30 | 6.57 | 6.18 |
| Mold | ND | 5.00 | ND | ND | 5.00 | ND | ND | 5.00 |
| Isolates | Nucleotide (bp) | Species | Identity (%) |
|---|---|---|---|
| 2S5 | 1466 | L. buchneri | 99.86 |
| 3S3 | 1438 | L. buchneri | 99.93 |
| 4M4 | 1449 | L. brevis | 100.0 |
| 5M2 | 1454 | L. brevis | 100.0 |
| 6M1 | 1446 | L. buchneri | 99.86 |
| 6S4 | 1446 | L. buchneri | 99.65 |
| Item 2 | Treatment 1 | SEM | P-Value | |
|---|---|---|---|---|
| CON | INO | |||
| Dry matter | 283 | 285 | 4.37 | 0.689 |
| Crude protein | 86.4 | 87.0 | 6.04 | 0.897 |
| Ether extract | 33.8 | 34.8 | 0.98 | 0.240 |
| Crude ash | 47.4 | 45.9 | 2.00 | 0.401 |
| Neutral detergent fiber | 470 | 469 | 6.52 | 0.860 |
| Acid detergent fiber | 212 | 213 | 7.47 | 0.798 |
| Hemicellulose | 259 | 256 | 4.62 | 0.471 |
| IVDMD | 668 | 662 | 29.7 | 0.758 |
| IVNDFD | 449 | 452 | 38.3 | 0.936 |
| Item 2 | Treatment 1 | SEM | P-Value | |
|---|---|---|---|---|
| CON | INO | |||
| Dry matter | 266 | 268 | 2.31 | 0.259 |
| Crude protein | 89.8 | 89.6 | 1.60 | 0.833 |
| Ether extract | 42.7 | 42.3 | 3.49 | 0.907 |
| Crude ash | 46.9 | 45.8 | 1.25 | 0.193 |
| Neutral detergent fiber | 493 a | 456 b | 14.8 | 0.013 |
| Acid detergent fiber | 275 a | 250 b | 8.59 | 0.006 |
| Hemicellulose | 218 | 207 | 8.36 | 0.100 |
| IVDMD | 625 b | 652 a | 11.7 | 0.029 |
| IVNDFD | 444 b | 484 a | 4.08 | <0.001 |
| Item | Treatment 1 | SEM | P-value | |
|---|---|---|---|---|
| CON | INO | |||
| Fermentation characteristics | ||||
| pH | 3.80 | 3.78 | 0.03 | 0.363 |
| Ammonia-N, g/kg DM | 0.60 | 0.50 | 0.05 | 0.537 |
| Lactate, g/kg DM | 130.0 | 129.0 | 3.01 | 0.864 |
| Acetate, g/kg DM | 20.9 b | 24.9 a | 1.28 | 0.019 |
| Propionate, g/kg DM | ND | ND | - | |
| Butyrate, g/kg DM | ND | ND | - | |
| Lactate: acetate | 6.22 a | 5.22 b | 0.314 | 0.002 |
| Microbe count, log10 cfu/g | ||||
| Lactic acid bacteria | 6.82 | 6.89 | 0.123 | 0.474 |
| Yeast | 5.89 a | 5.79 b | 0.041 | 0.029 |
| Mold | ND | ND | - | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paradhipta, D.H.V.; Lee, S.S.; Kang, B.; Joo, Y.H.; Lee, H.J.; Lee, Y.; Kim, J.; Kim, S.C. Dual-Purpose Inoculants and Their Effects on Corn Silage. Microorganisms 2020, 8, 765. https://doi.org/10.3390/microorganisms8050765
Paradhipta DHV, Lee SS, Kang B, Joo YH, Lee HJ, Lee Y, Kim J, Kim SC. Dual-Purpose Inoculants and Their Effects on Corn Silage. Microorganisms. 2020; 8(5):765. https://doi.org/10.3390/microorganisms8050765
Chicago/Turabian StyleParadhipta, Dimas Hand Vidya, Seong Shin Lee, Byeongsam Kang, Young Ho Joo, Hyuk Jun Lee, Yeyeong Lee, Jinwoo Kim, and Sam Churl Kim. 2020. "Dual-Purpose Inoculants and Their Effects on Corn Silage" Microorganisms 8, no. 5: 765. https://doi.org/10.3390/microorganisms8050765
APA StyleParadhipta, D. H. V., Lee, S. S., Kang, B., Joo, Y. H., Lee, H. J., Lee, Y., Kim, J., & Kim, S. C. (2020). Dual-Purpose Inoculants and Their Effects on Corn Silage. Microorganisms, 8(5), 765. https://doi.org/10.3390/microorganisms8050765