Knockdown of Dinoflagellate Condensin CcSMC4 Subunit Leads to S-Phase Impediment and Decompaction of Liquid Crystalline Chromosomes


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Dinoflagellate Cell Cultures and Flow Cytometry
2.2. In Silico Analysis
2.3. Immunological Techniques, Preparation of Cell Lysates and Molecular Biological Techniques
2.4. Functional Suppression of cut3-477 Conditional Lethality through CcSMC4p Expression
2.5. Antisense Oligonucleotide-Mediated Gene-Knockdown Experiment
3. Results
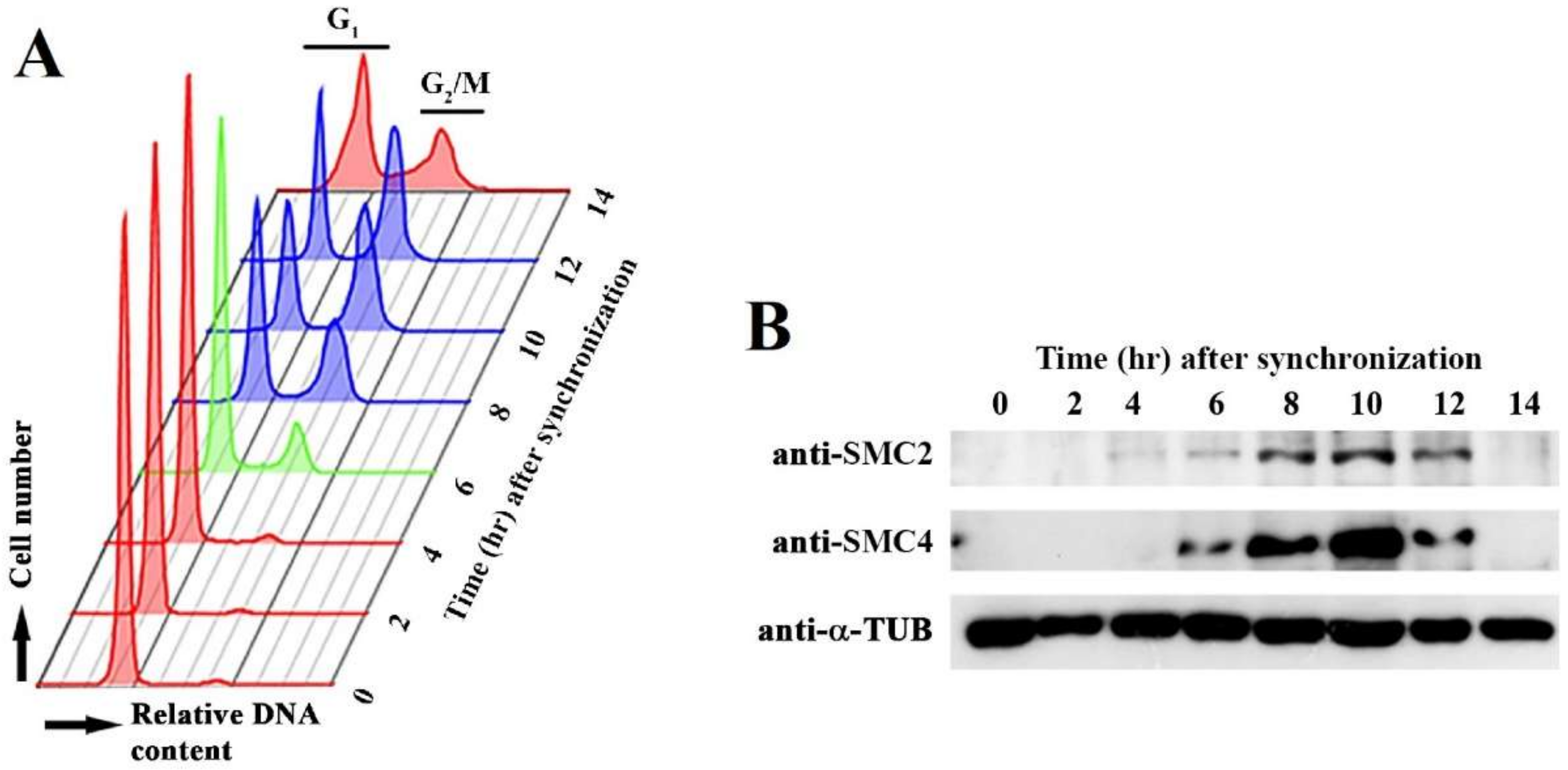
3.1. Cell-Cycle-Phased Expression of CcSMC4p
3.2. Heterologous CcSMC4p Expression Suppressed Conditional Lethality of Fission Yeast cut3-477
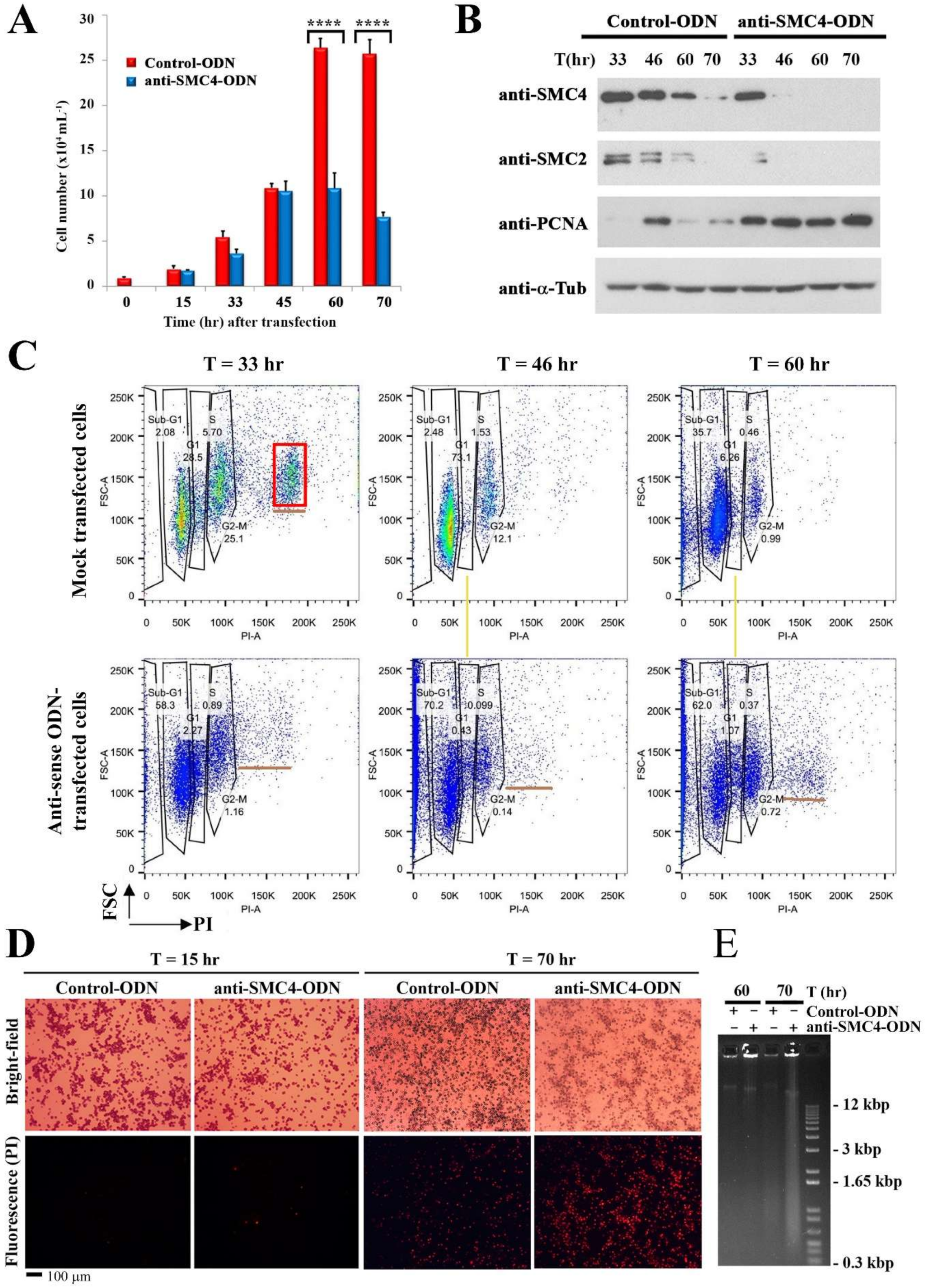
3.3. Prolong CcSMC4p-Knockdown Led to S-Phase Impediment with Nuclear Swelling
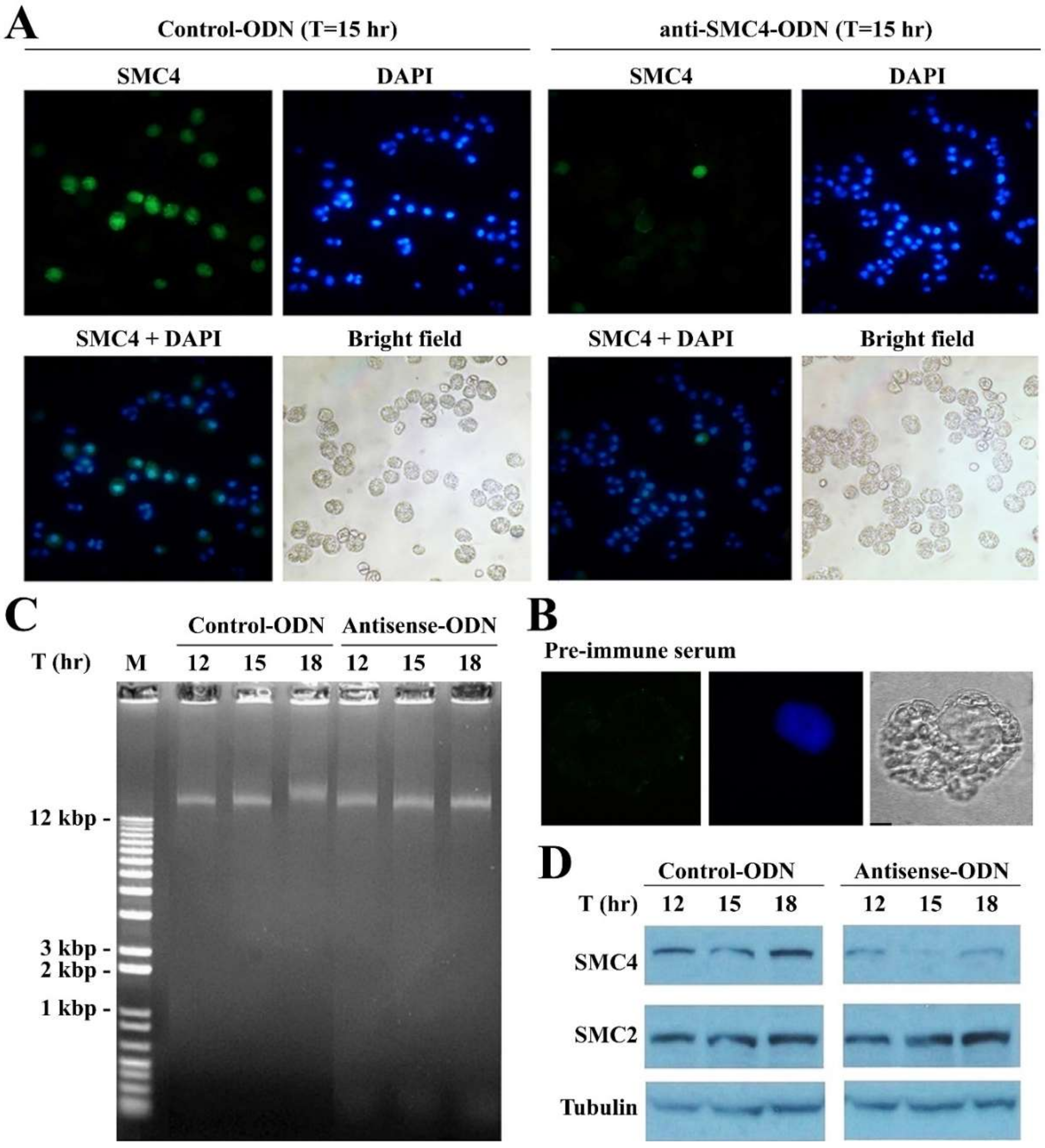
3.4. CcSMC4p-Knockdown Led to S-Phase Impediment from the First Cell Cycle
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gibcus, J.H.; Samejima, K.; Goloborodko, A.; Samejima, I.; Naumova, N.; Nuebler, J.; Kanemaki, M.T.; Xie, L.; Paulson, J.R.; Earnshaw, W.C.; et al. A pathway for mitotic chromosome formation. Science 2018, 359, eaao6135. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, P.J.; Nooden, L.D. Isolation and partial characterization of dinoflagellate chromatin. Biochim. Biophys. Acta 1974, 349, 402–414. [Google Scholar] [CrossRef]
- Shupe, K.; Rizzo, P.J. Nuclease digestion of dhromatin from the eukaryotic algae Olisthodiscus luteus, Peridinium balticum, and Crypthecodinium cohnii. J. Protozool. 1983, 30, 599–606. [Google Scholar] [CrossRef]
- Herzog, M.; Soyer, M.O. Distinctive features of dinoflagellate chromatin. Absence of nucleosomes in a primitive species Prorocentrum micans E. Eur. J. Cell Biol. 1981, 23, 295–302. [Google Scholar]
- Spector, D.L. Dinoflagellate nuclei. In Dinoflagellates; Academic Press: San Diego, CA, USA, 1984; Volume 1, pp. 107–147. [Google Scholar]
- Chow, M.H.; Yan, K.T.H.; Bennett, M.J.; Wong, J.T.Y. Liquid crystalline chromosomes: Birefringence and DNA condensation. Eukaryot. Cell 2010, 10, 1577–1587. [Google Scholar] [CrossRef]
- Wong, J.T.Y. Architectural organization of dinoflagellate liquid crystalline chromosomes. Microorganisms 2019, 7, 27. [Google Scholar] [CrossRef]
- Ziv, R.; Levin-Zaidman, S.; Gutman, S.B.; Arad, T.; Minsky, A. Supercoiling-regulated liquid-crystalline packaging of topologically-constrained, nucleosome-free DNA molecules. Biochemistry 1984, 33, 14177–14184. [Google Scholar] [CrossRef]
- Levi-Setti, R.; Gavrilov, K.L.; Rizzo, P.J. Divalent cation distribution in dinoflagellate chromosomes imaged by high-resolution ion probe mass spectrometry. Eur. J. Cell Biol. 2008, 87, 963–976. [Google Scholar] [CrossRef]
- Strick, R.; Strissel, P.L.; Gavrilov, K.; Levi-Setti, R. Cation–chromatin binding as shown by ion microscopy is essential for the structural integrity of chromosomes. J. Cell Biol. 2001, 155, 899–910. [Google Scholar] [CrossRef]
- Herzog, M.; Soyer, M.O. The native structure of dinoflagellate chromosomes and their stabilization by Ca2+ and Mg2+ cations. Eur. J. Cell Biol. 1983, 30, 33–41. [Google Scholar]
- Sun, S.; Wong, J.T.; Liu, M.; Dong, F. Counterion-mediated decompaction of liquid crystalline chromosomes. DNA Cell Biol. 2012, 31, 1657–1664. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.T.Y. Architectural Organization of Dinoflagellate Liquid Crystalline Chromosomes–the non-nucleosomal eukaryotic chromosome packaging system. Front. Microbiol. 2018. [Google Scholar] [CrossRef]
- Hirano, T. Condensins: Universal organizers of chromosomes with diverse functions. Genes Dev. 2012, 26, 1659–1678. [Google Scholar] [CrossRef] [PubMed]
- Cuylen, S.; Metz, J.; Haering, C.H. Condensin structures chromosomal DNA through topological links. Nat. Struct. Mol. Biol. 2011, 18, 894–901. [Google Scholar] [CrossRef]
- Hirano, T. SMC proteins and chromosome mechanics: From bacteria to humans. Philos. Trans. R. Soc. Lond B Biol. Sci. 2005, 360, 507–514. [Google Scholar] [CrossRef]
- Onn, I.; Aono, N.; Hirano, M.; Hirano, T. Reconstitution and subunit geometry of human condensin complexes. EMBO J. 2007, 26, 1024–1034. [Google Scholar] [CrossRef]
- Kschonsak, M.; Merkel, F.; Bisht, S.; Metz, J.; Rybin, V.; Hassler, M.C.H. Structural basis for a safety-belt mechanism that anchors condensin to chromosomes. Cell 2017, 17, 588–600. [Google Scholar] [CrossRef]
- Rana, V.; Bosco, G. Condensin regulation of genome architecture. J. Cell. Physiol. 2017, 232, 1617–1625. [Google Scholar] [CrossRef]
- Ganji, M.; Shaltiel, I.A.; Bisht, S.; Kim, E.; Kalichava, A.; Haering, C.H.; Dekker, C. Real-time imaging of DNA loop extrusion by condensin. Science 2018, 360, 102–105. [Google Scholar] [CrossRef]
- Case, R.B.; Chang, Y.P.; Smith, S.B.; Gore, J.; Cozzarelli, N.R.; Bustamante, C. The bacterial condensin MukBEF compacts DNA into a repetitive, stable structure. Science 2004, 305, 222–227. [Google Scholar] [CrossRef][Green Version]
- Petrushenko, Z.M.; Lai, C.-H.; Rai, R.; Rybenkov, V.V. DNA reshaping by MukB right-handed knoting and left-handed supercoiling. J. Biol. Chem. 2006, 281, 4606–4615. [Google Scholar] [CrossRef] [PubMed]
- Kwok, A.C.; Wong, J.T.Y. The activity of a wall-bound cellulase is required for and is coupled to cell cycle progression in the dinoflagellate Crypthecodinium Cohnii. Plant. Cell 2010, 22, 1281–1298. [Google Scholar] [CrossRef] [PubMed]
- Wong, J.T.Y.; Whiteley, A. An improved method of cell cycle synchronisation for the heterotrophic dinoflagellate Crypthecodinium Cohnii Biecheler analyzed by flow cytometry. J. Exp. Mar. Biol. Ecol. 1996, 197, 91–99. [Google Scholar] [CrossRef]
- Kwok, A.C.; Mak, C.C.; Wong, F.T.; Wong, J.T. Novel method for preparing spheroplasts from cells with an internal cellulosic cell wall. Eukaryot. Cell 2007, 6, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Harlow, E.; Lane, D.P. Antibodies: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Plainview, NY, USA, 1988. [Google Scholar]
- Chan, W.S.; Kwok, A.C.M.; Wong, J.T.Y. Knockdown of dinoflagellate cellulose synthase CesA1 resulted in malformed intracellular cellulosic thecal plates and severely impeded cyst-to-swarmer transition. Front. Microbiol. 2019, 10, 546. [Google Scholar] [CrossRef] [PubMed]
- Leveson, A.C.; Wong, J.T.Y. PCNA-like proteins in dinoflagellates. J. Phycol. 1999, 35, 798–805. [Google Scholar] [CrossRef]
- Liu, J.; Jiao, N.; Hong, H.; Luo, T.; Cai, H. Proliferating cell nuclear antigen (PCNA) as a marker of cell proliferation in the marine dinoflagellate Prorocentrum donghaiense Lu and the green alga Dunaliella salina Teodoresco. J. Appl. Phycol. 2005, 17, 323–330. [Google Scholar] [CrossRef]
- Hirano, T.; Funahashi, S.; Uemura, T.; Yanagida, M. Isolation and characterization of Schizosaccharomyces pombe cut m.utants that block nuclear division but not cytokinesis. EMBO J. 1986, 5, 2973–2979. [Google Scholar] [CrossRef]
- Saka, Y.; Sutani, T.; Yamashita, Y.; Saitoh, S.; Takeuchi, M.; Nakaseko, Y.; Yanagida, M. Fission yeast cut3 and cut14, members of a ubiquitous protein family, are required for chromosome condensation and segregation in mitosis. EMBO J. 1994, 13, 4938–4952. [Google Scholar] [CrossRef]
- Maundrell, K. nmt1 of fission yeast. A highly transcribed gene completely repressed by thiamine. J. Biol. Chem. 1990, 265, 10857–10864. [Google Scholar]
- Forsburg, S.L.; Rhind, N. Basic methods for fission yeast. Yeast 2006, 23, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, A.; Kimura, K.; Yokoyama, S.; Hanaoka, F. Cell Cycle-dependent Phosphorylation, Nuclear Localization, and Activation of Human Condensin. J. Biol. Chem. 2004, 279, 4551–4559. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.; Patkaniowska, A.; Urlaub, H.; Luhrmann, R.; Tuschl, T. Single-stranded antisense siRNAs guide target RNA cleavage in RNAi. Cell 2002, 110, 563–574. [Google Scholar] [CrossRef]
- Ono, T.; Yamashita, D.; Hirano, T. Condensin II initiates sister chromatid resolution during S phase. J. Cell Biol. 2013, 200, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Mak, C.K.M.; Hung, V.K.L.; Wong, J.T.Y. Type II topoisomerase activities in both the G1 and G2/M phases of the dinoflagellate cell cycle. Chromosoma 2005, 114, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Yeung, P.K.; New, D.C.; Leveson, A.; Yam, C.H.; Poon, R.Y.; Wong, J.T. The spindle checkpoint in the dinoflagellate Crypthecodinium cohnii. Exp. Cell Res. 2000, 254, 120–129. [Google Scholar] [CrossRef]
- Lam, C.M.C.; Yeung, P.K.K.; Lee, H.C.; Wong, J.T. Cyclic ADP-ribose links metabolism to multiple fission in the dinoflagellate Crypthecodinium cohnii. Cell Calcium 2009, 45, 346–357. [Google Scholar] [CrossRef]
- Blow, J.J.; Dutta, A. Preventing re-replication of chromosomal DNA. Nat. Rev. Mol. Cell Biol. 2005, 6, 476–486. [Google Scholar] [CrossRef]
- Heale, J.T.; Ball, A.R.; Schmiesing, J.A.; Kim, J.-S.; Kong, X.; Zhou, S.; Hudson, D.F.; Earnshaw, W.C.; Yokomori, K. Condensin I interacts with the PARP-1-XRCC1 complex and functions in DNA single-strand break repair. Mol. Cell 2006, 21, 837–848. [Google Scholar] [CrossRef]
- Wu, N.; Yu, H. The Smc complexes in DNA damage response. Cell Biosci. 2012, 2, 5. [Google Scholar] [CrossRef]
- Waseem, N.H.; Labib, K.; Nurse, P.; Lane, D.P. Isolation and analysis of the fission yeast gene encoding polymerase delta accessory protein PCNA. EMBO J. 1992, 11, 5111–5120. [Google Scholar] [CrossRef] [PubMed]
- Maeshima, K.; Tamura, S.; Shimamoto, Y. Chromatin as a nuclear spring. Biophys. Phys. 2018, 15, 189–195. [Google Scholar] [CrossRef]
- Marko, J.F. Micromechanical studies of mitotic chromosomes. Chromosome Res. 2008, 16, 469–497. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, S.H.; Hirano, T. HEAT repeats-versatile arrays of amphiphilic helices working in crowded environments? J. Cell Sci. 2016, 129, 3963–3970. [Google Scholar] [CrossRef] [PubMed]
- Houlard, M.; Godwin, J.; Metson, J.; Lee, J.; Hirano, T.; Nasmyth, K. Condensin confers the longitudinal rigidity of chromosomes. Nat. Cell Biol. 2015, 17, 771. [Google Scholar] [CrossRef]
- Gan, H.; Serra-Cardona, A.; Hua, X.; Zhou, H.; Labib, K.; Yu, C.; Zhang, Z. The Mcm2-Ctf4-Polα axis facilitates parental histone H3-H4 transfer to lagging strands. Mol. Cell 2018, 72, 140–151. [Google Scholar] [CrossRef]
- Foltman, M.; Evrin, C.; De Piccoli, G.; Jones, R.C.; Edmondson, R.D.; Katou, Y.; Nakato, R.; Shirahige, K.; Labib, K. Eukaryotic replisome components cooperate to process histones during chromosome replication. Cell Rep. 2013, 3, 892–904. [Google Scholar] [CrossRef]
- Johzuka, K.; Terasawa, M.; Ogawa, H.; Ogawa, T.; Horiuchi, T. Condensin loaded onto the Replication Fork Barrier Site in the rRNA Gene Repeats during S phase in a FOB1-dependent fashion to prevent contraction of a long repetitive array in Saccharomyces cerevisiae. Mol. Cell. Biol. 2006, 26, 2226–2236. [Google Scholar] [CrossRef]
- Bhaud, Y.; Guillebault, D.; Lennon, J.; Defacque, H.; Soyer-Gobillard, M.O.; Moreau, H. Morphology and behaviour of dinoflagellate chromosomes during the cell cycle and mitosis. J. Cell Sci. 2000, 113, 1231–1239. [Google Scholar]
- Woodward, A.M.; Göhler, T.; Luciani, M.G.; Oehlmann, M.; Ge, X.; Gartner, A.; Jackson, D.A.; Blow, J.J. Excess Mcm2–7 license dormant origins of replication that can be used under conditions of replicative stress. J. Cell Biol. 2006, 173, 673–683. [Google Scholar] [CrossRef]
- Vas, A.; Mok, W.; Leatherwood, J. Control of DNA rereplication via Cdc2 phosphorylation sites in the origin recognition complex. Mol. Cell. Biol. 2001, 21, 5767–5777. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.W.; Jallepalli, P.V.; Huneycutt, B.J.; Kelly, T.J. Interaction of the S phase regulator cdc18 with cyclin-dependent kinase in fission yeast. Proc. Natl. Acad. Sci. USA 1997, 94, 6142–6147. [Google Scholar] [CrossRef] [PubMed]
- Dewar, J.M.; Walter, J.C. Mechanisms of DNA replication termination. Nat. Rev. Mol. Cell Biol. 2017, 18, 507. [Google Scholar] [CrossRef] [PubMed]
- Galarreta, A.; Lecona, E.; Valledor, P.; Ubieto, P.; Lafarga, V.; Specks, J.; Fernandez-Capetillo, O. USP7 couples DNA replication termination to mitotic entry. bioRxiv 2018, 1, 305318. [Google Scholar] [CrossRef]
- Sutani, T.; Yuasa, T.; Tomonaga, T.; Dohmae, N.; Takio, K.; Yanagida, M. Fission yeast condensin complex: Essential roles of non-SMC subunits for condensation and Cdc2 phosphorylation of Cut3/SMC4. Genes Develop. 1999, 13, 2271–2283. [Google Scholar] [CrossRef]
- Samejima, I.; Matsumoto, T.; Nakaseko, Y.; Beach, D.; Yanagida, M. Identification of seven new cut genes involved in Schizosaccharomyces pombe mitosis. J. Cell Sci. 1993, 105, 135–143. [Google Scholar]
- Liu, C.-m.; McElver, J.; Tzafrir, I.; Joosen, R.; Wittich, P.; Patton, D.; Van Lammeren, A.A.M.; Meinke, D. Condensin and cohesin knockouts in Arabidopsis exhibit a titan seed phenotype. Plant. J. 2002, 29, 405–415. [Google Scholar] [CrossRef]
- Lioy, V.S.; Cournac, A.; Marbouty, M.; Duigou, S.; Mozziconacci, J.; Espéli, O.; Boccard, F.; Koszul, R. Multiscale structuring of the E. coli chromosome by nucleoid-associated and condensin proteins. Cell 2018, 172, 771–783. [Google Scholar] [CrossRef]
- Trojanowski, D.; Hołówka, J.; Zakrzewska-Czerwińska, J. Where and When Bacterial Chromosome Replication Starts: A Single Cell Perspective. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, T.H.K.; Wu, Z.; Kwok, A.C.M.; Wong, J.T.Y. Knockdown of Dinoflagellate Condensin CcSMC4 Subunit Leads to S-Phase Impediment and Decompaction of Liquid Crystalline Chromosomes. Microorganisms 2020, 8, 565. https://doi.org/10.3390/microorganisms8040565
Yan THK, Wu Z, Kwok ACM, Wong JTY. Knockdown of Dinoflagellate Condensin CcSMC4 Subunit Leads to S-Phase Impediment and Decompaction of Liquid Crystalline Chromosomes. Microorganisms. 2020; 8(4):565. https://doi.org/10.3390/microorganisms8040565
Chicago/Turabian StyleYan, Ting Hin Kosmo, Zhihao Wu, Alvin Chun Man Kwok, and Joseph Tin Yum Wong. 2020. "Knockdown of Dinoflagellate Condensin CcSMC4 Subunit Leads to S-Phase Impediment and Decompaction of Liquid Crystalline Chromosomes" Microorganisms 8, no. 4: 565. https://doi.org/10.3390/microorganisms8040565
APA StyleYan, T. H. K., Wu, Z., Kwok, A. C. M., & Wong, J. T. Y. (2020). Knockdown of Dinoflagellate Condensin CcSMC4 Subunit Leads to S-Phase Impediment and Decompaction of Liquid Crystalline Chromosomes. Microorganisms, 8(4), 565. https://doi.org/10.3390/microorganisms8040565