Preliminary In Vitro Studies on Corynebacterium urealyticum Pathogenetic Mechanisms, a Possible Candidate for Chronic Idiopathic Prostatitis?


 ,
,  , , and
, , and _Di_Marco.png)
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Bacterial Strains
2.3. Cell Cultures
2.4. Influence of AHA on In Vitro Growth Curves by Cell Counting
2.5. In Vitro Bacterial Grown Curve Analysis
2.6. MTT Assay
2.7. In Vitro Biofilm Formation Assay
2.8. Effect of AHA on C. urealyticum Adhesion to Human LNCaP and HeLa Cell Lines
2.9. Statistical Analysis
3. Results
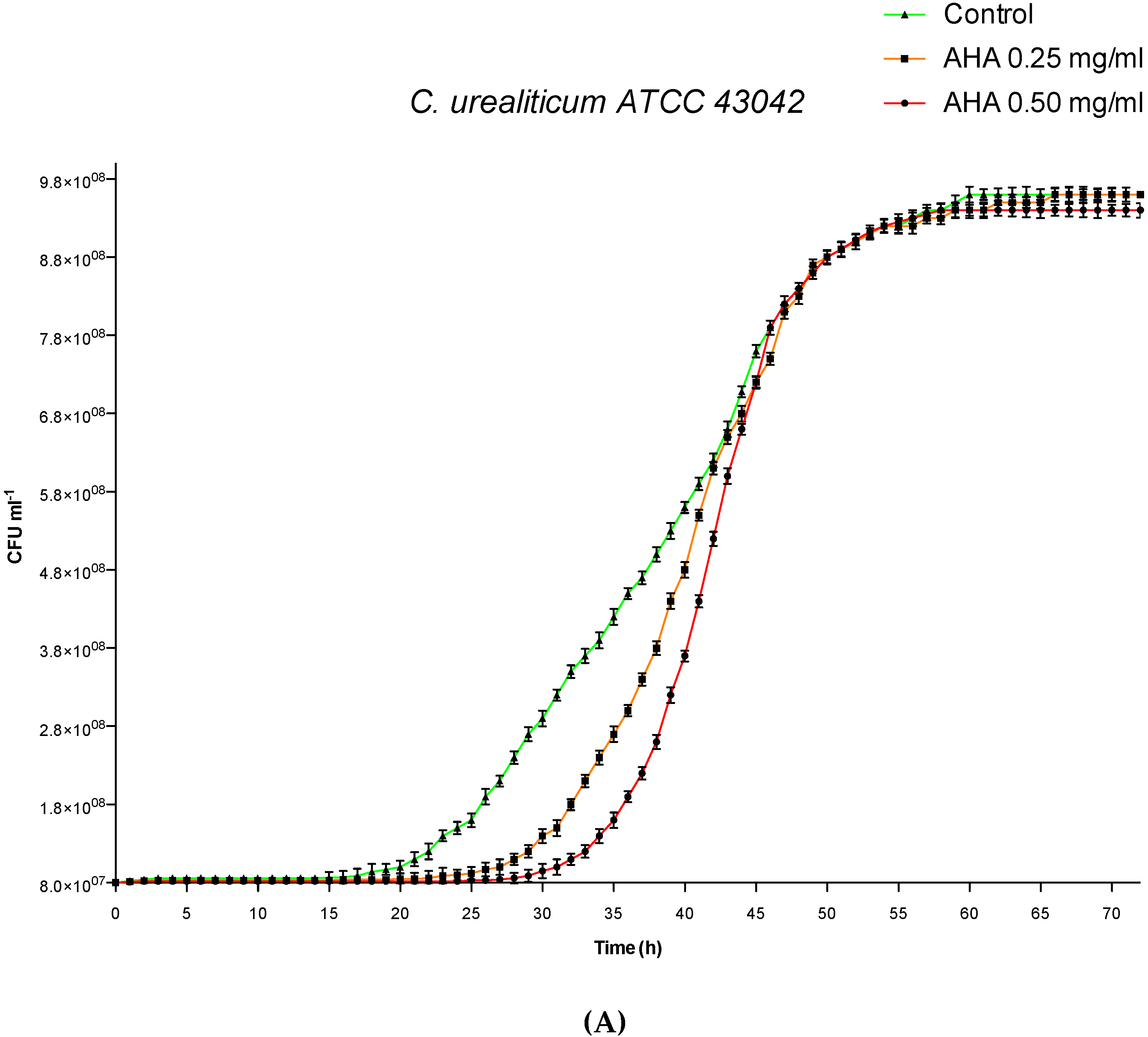
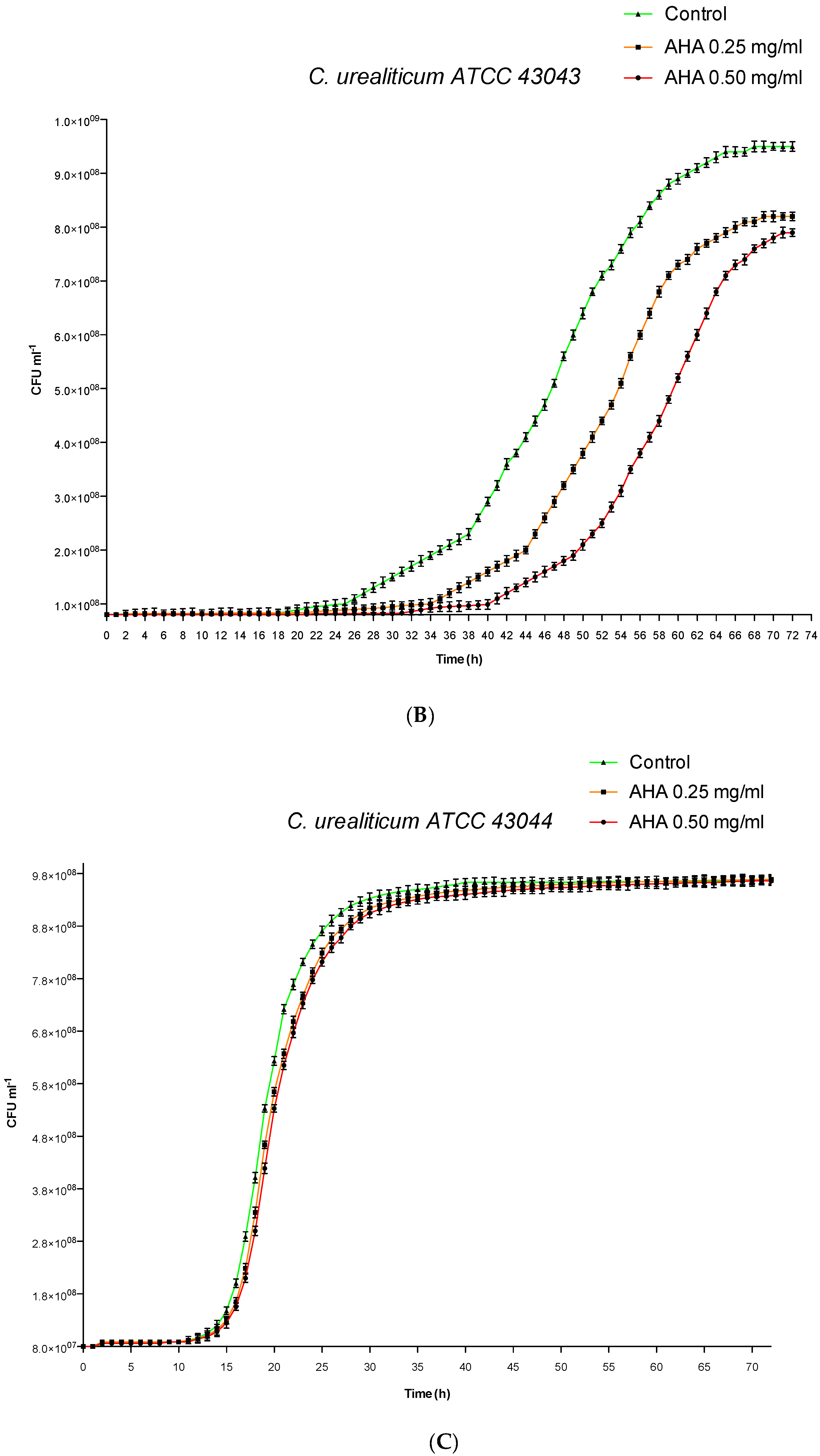
3.1. Influence of AHA on in vitro Growth Curves by Cell Counting and Analysis
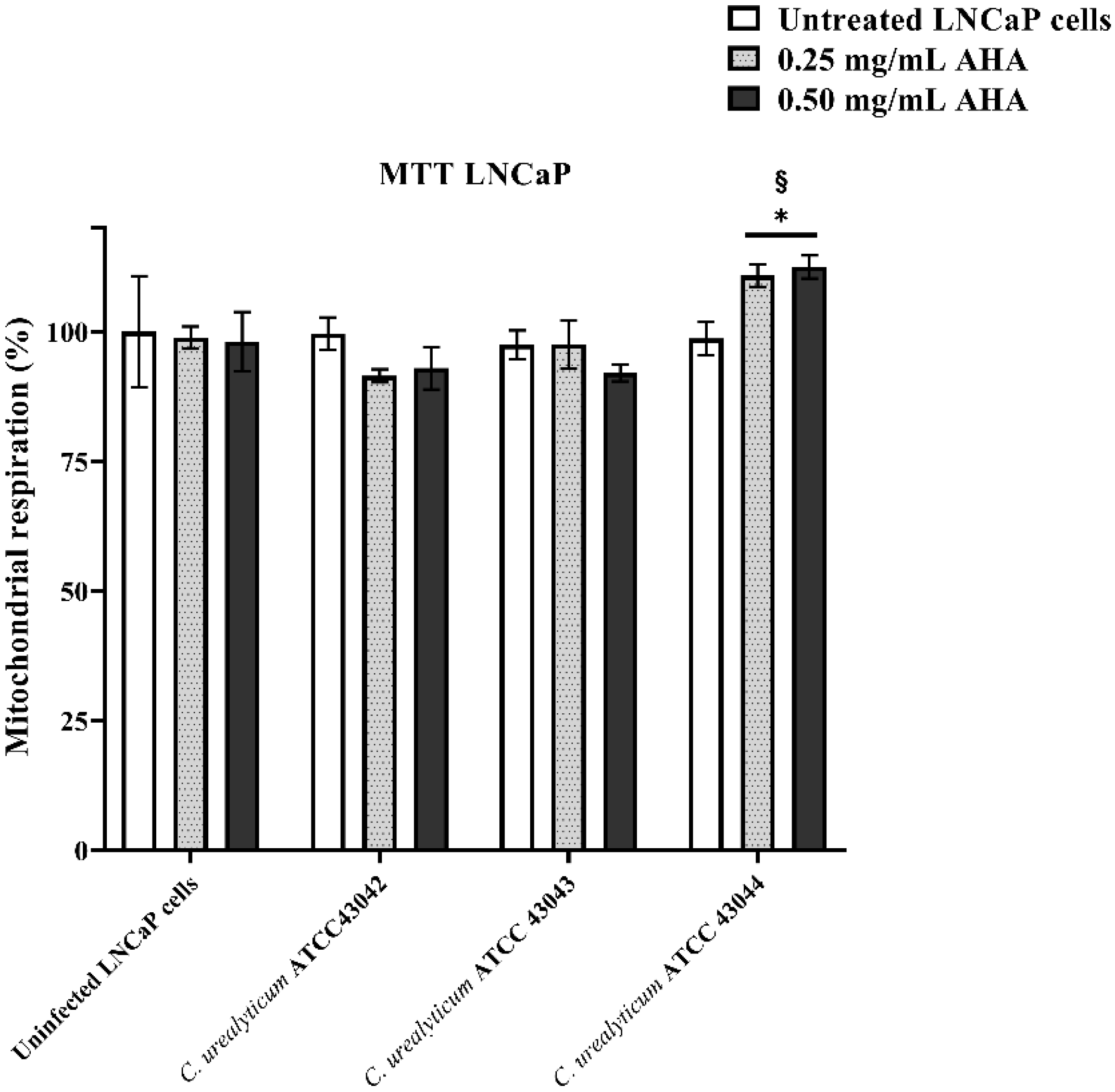
3.2. Effects of AHA Treatments and C. urealyticum Infections on LNCaP and HeLa Cell Viability
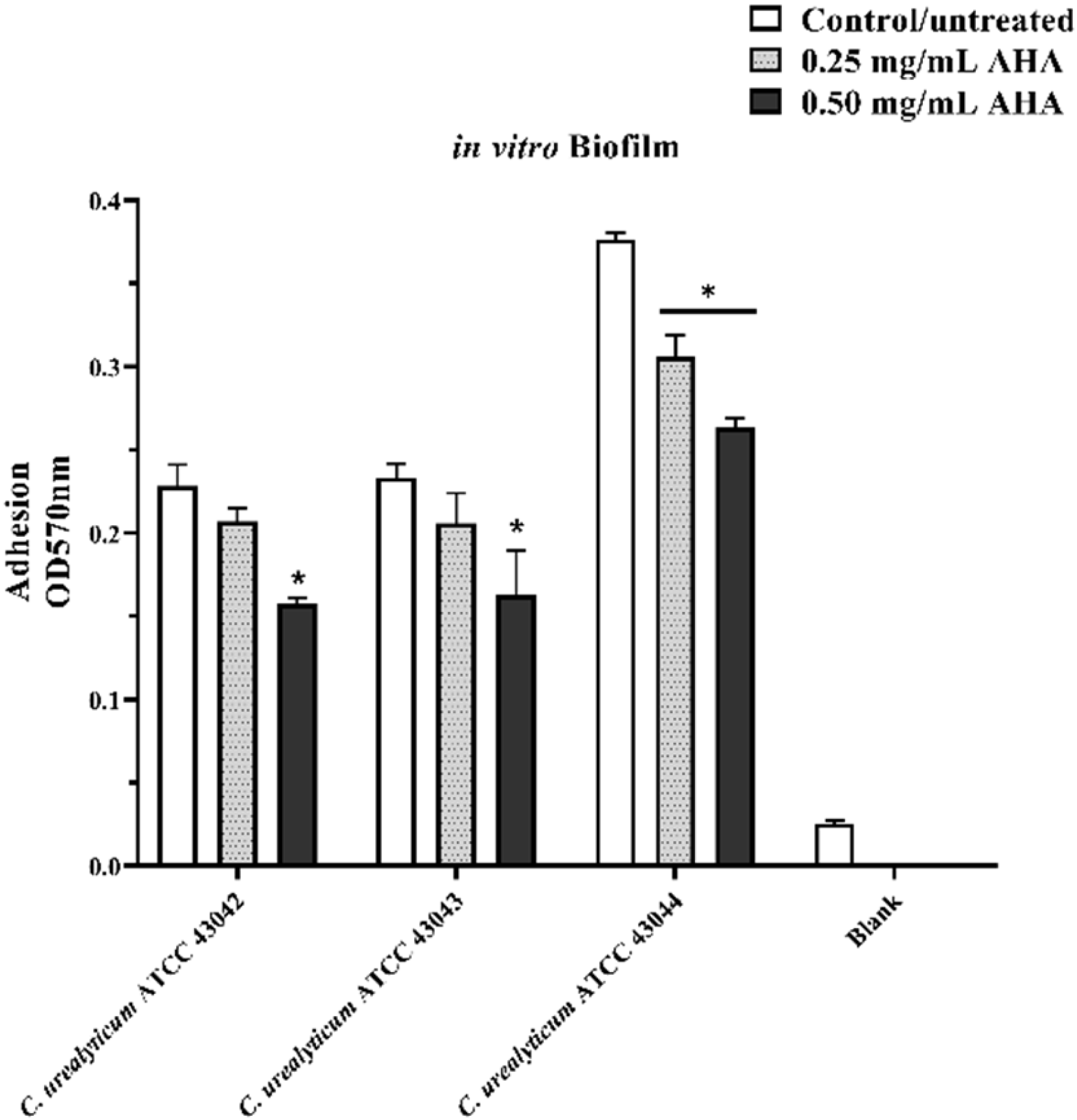
3.3. Effects of AHA Treatments on in Vitro Biofilm Formation Assay
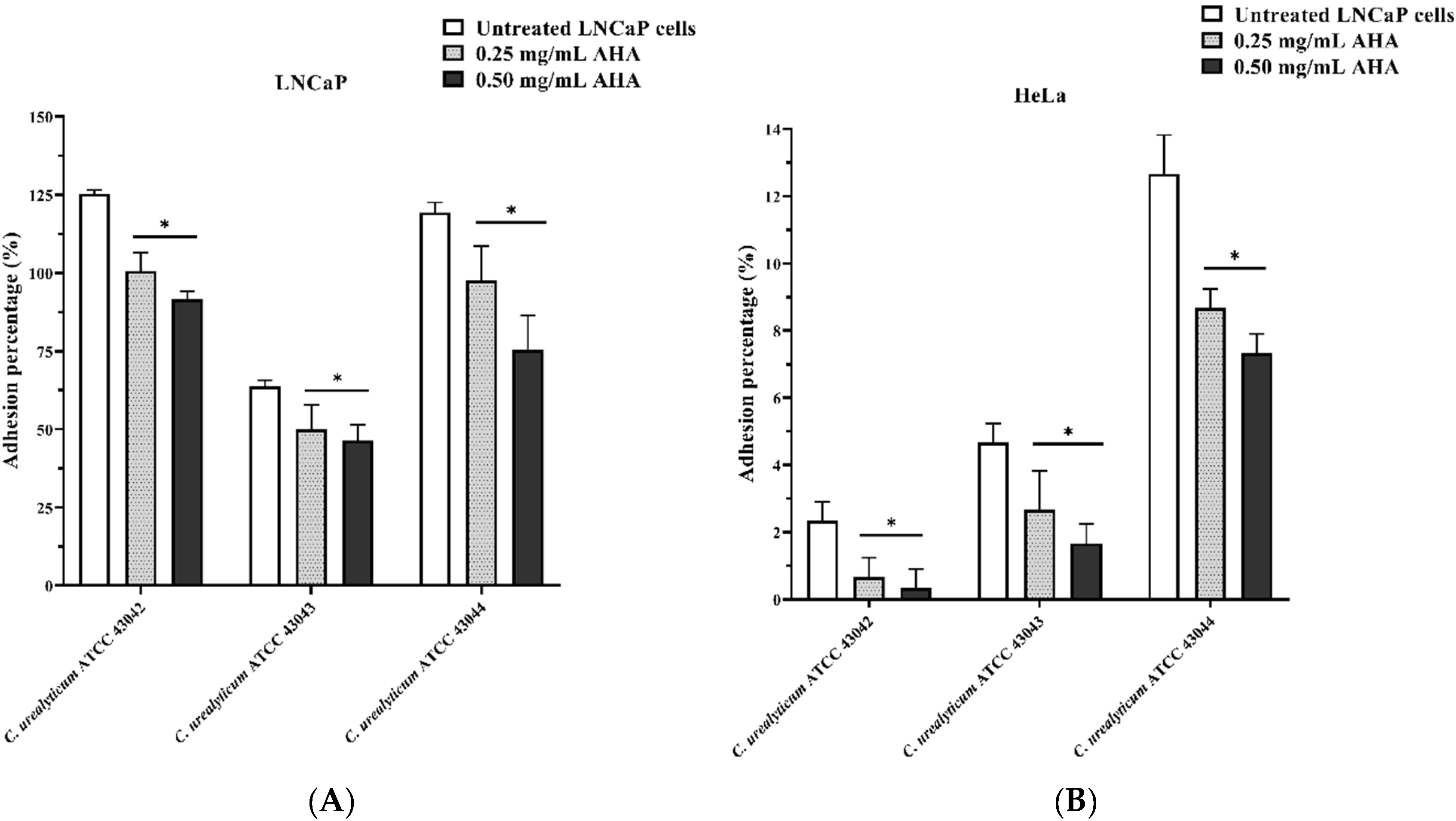
3.4. Effect of AHA on C. urealyticum Adhesion to Human LNCaP and HeLa Cell Lines
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Bernard, K.A.; Funke, G. Corynebacterium. In Bergey’s Manual of Systematics of Archaea and Bacteria; Wiley: Hoboken, NJ, USA, 2015; pp. 1–70. [Google Scholar]
- González-Enguita, C.; Vela-Navarrete, R. Corynebacterium urealyticum: The historical importance of its discovery. Actas Urol. Esp. 2017, 41, 140. [Google Scholar] [CrossRef]
- Bernard, K. The genus Corynebacterium and other medically relevant coryneform-like bacteria. J. Clin. Microbiol. 2012, 50, 3152–3158. [Google Scholar] [CrossRef] [PubMed]
- Salem, N.; Salem, L.; Saber, S.; Ismail, G.; Bluth, M.H. Corynebacterium urealyticum: A comprehensive review of an understated organism. Infect. Drug Resist. 2015, 8, 129. [Google Scholar] [PubMed]
- Costales, J.; Alsyouf, M.; Napolitan, P.; Wang, S.; Hu, B. Corynebacterium urealyticum: Rare urinary tract infection with serious complications. Can. J. Urol. 2019, 26, 9680–9682. [Google Scholar] [PubMed]
- François, J. La cystite incrustée. J. Urol. Med. Chair 1914, 5, 35–52. [Google Scholar]
- El Sayegh, H.; Elouardani, M.; Iken, A.; Nouini, Y.; Lachkar, A.; Benslimane, L.; Belahnech, Z.; Faik, M. Cystite incrustante à Corynebacterium urealyticum. La Revue de Médecine Interne 2008, 29, 328–330. [Google Scholar] [CrossRef]
- Pardo, M.A.; Rosino, S.A.; Rivero, G.Á.; Barceló, B.I. Encrusted prostatitis by Corynebacterium urealyticum: The importance of clinical suspicion. Actas Urol. Esp. 2018. [Google Scholar] [CrossRef]
- López-Medrano, F.; García-Bravo, M.; Morales, J.; András, A.; San Juan, R.; Lizasoain, M.; Aguado, J. Urinary tract infection due to Corynebacterium urealyticum in kidney transplant recipients: An underdiagnosed etiology for obstructive uropathy and graft dysfunction—results of a prospective cohort study. Clin. Infect. Dis. 2008, 46, 825–830. [Google Scholar] [CrossRef]
- Riegel, P. Actualités de l′ épidémiologie et du rôle pathogène des corynébactéries. Antibiotiques 2006, 8, 153–161. [Google Scholar] [CrossRef]
- Soriano, F.; Tauch, A. Microbiological and clinical features of Corynebacterium urealyticum: Urinary tract stones and genomics as the Rosetta Stone. Clin. Microbiol. Infect. 2008, 14, 632–643. [Google Scholar] [CrossRef]
- Gómez-Garcés, J.-L.; Alos, J.-I.; Tamayo, J. In vitro activity of linezolid and 12 other antimicrobials against coryneform bacteria. Int. J. Antimicrob. Agents 2007, 29, 688–692. [Google Scholar] [CrossRef] [PubMed]
- Soriano, F.; Huelves, L.; Naves, P.; Rodríguez-Cerrato, V.; del Prado, G.; Ruiz, V.; Ponte, C. In vitro activity of ciprofloxacin, moxifloxacin, vancomycin and erythromycin against planktonic and biofilm forms of Corynebacterium urealyticum. J. Antimicrob. Chemother. 2009, 63, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.N.; Valadão, T.B.; Baio, P.V.P.; Mattos-Guaraldi, A.L.; Vieira, V.V. Novel mutations in the QRDR region gyrA gene in multidrug-resistance Corynebacterium spp. isolates from intravenous sites. Antonie Van Leeuwenhoek 2019, 1–4. [Google Scholar] [CrossRef]
- Souza, C.D.; Faria, Y.V.; Sant’Anna, L.D.O.; Viana, V.G.; Seabra, S.H.; Souza, M.C.D.; Vieira, V.V.; Hirata Júnior, R.; Moreira, L.D.O.; Mattos-Guaraldi, A.L.D. Biofilm production by multiresistant Corynebacterium striatumassociated with nosocomial outbreak. Memorias do Instituto Oswaldo Cruz 2015, 110, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Charalabopoulos, K.; Karachalios, G.; Baltogiannis, D.; Charalabopoulos, A.; Giannakopoulos, X.; Sofikitis, N. Penetration of antimicrobial agents into the prostate. Chemotherapy 2003, 49, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Elyasi, S.; Khalili, H.; Dashti-Khavidaki, S.; Mohammadpour, A. Vancomycin-induced nephrotoxicity: Mechanism, incidence, risk factors and special populations. A literature review. Eur. J. Clin. Pharmacol. 2012, 68, 1243–1255. [Google Scholar] [CrossRef]
- Wang, H.; Chen, Z.; Zhu, Y.; Wang, T.; Wu, X. Penetrability and therapeutic effect of vancomycin to the prostates of rats with bacterial prostatitis (BP) or BPH-BP. Zhonghua Nan Ke Xue Natl. J. Androl. 2006, 12, 490–495. [Google Scholar]
- Marty, N.; Agueda, L.; Lapchine, L.; Clave, D.; Henry-Ferry, S.; Chabanon, G. Adherence and hemagglutination ofCorynebacterium group D2. Eur. J. Clin. Microbiol. Infect. Dis. 1991, 10, 20–24. [Google Scholar] [CrossRef]
- Mandlik, A.; Swierczynski, A.; Das, A.; Ton-That, H. Corynebacterium diphtheriae employs specific minor pilins to target human pharyngeal epithelial cells. Mol. Microbiol. 2007, 64, 111–124. [Google Scholar] [CrossRef]
- Tauch, A.; Trost, E.; Tilker, A.; Ludewig, U.; Schneiker, S.; Goesmann, A.; Arnold, W.; Bekel, T.; Brinkrolf, K.; Brune, I. The lifestyle of Corynebacterium urealyticum derived from its complete genome sequence established by pyrosequencing. J. Biotechnol. 2008, 136, 11–21. [Google Scholar] [CrossRef]
- Richmond, S.; Yep, A. Quantification of Urease Activity. In Proteus Mirabilis; Springer: Berlin/Heidelberg, Germany, 2019; pp. 85–96. [Google Scholar]
- Genovese, C.; Davinelli, S.; Mangano, K.; Tempera, G.; Nicolosi, D.; Corsello, S.; Vergalito, F.; Tartaglia, E.; Scapagnini, G.; Di Marco, R. Effects of a new combination of plant extracts plus d-mannose for the management of uncomplicated recurrent urinary tract infections. J. Chemother. 2018, 30, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Sabiote, L.; Emiliani, E.; Kanashiro, A.K.; Balañà, J.; Mosquera, L.; Sánchez-Martín, F.M.; Millán, F.; Alonso, C.; Palou, J.; Angerri, O. Oral Acidification with l-Methionine as a Noninvasive Treatment for Encrusted Uropathy. J. Endourol. Case Rep. 2020. [Google Scholar] [CrossRef]
- Miano, R.; Germani, S.; Vespasiani, G. Stones and urinary tract infections. Urol. Int. 2007, 79, 32–36. [Google Scholar] [CrossRef]
- Rahman, N.U.; Meng, M.V.; Stoller, M.L. Infections and urinary stone disease. Curr. Pharm. Des. 2003, 9, 975–981. [Google Scholar] [CrossRef]
- Cappuccino, L.; Bottino, P.; Torricella, A.; Pontremoli, R. Nephrolithiasis by Corynebacterium urealyticum infection: Literature review and case report. J. Nephrol. 2014, 27, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Fuochi, V.; Li Volti, G.; Camiolo, G.; Tiralongo, F.; Giallongo, C.; Distefano, A.; Petronio Petronio, G.; Barbagallo, I.; Viola, M.; Furneri, P.M. Antimicrobial and anti-proliferative effects of skin mucus derived from Dasyatis pastinaca (Linnaeus, 1758). Mar. Drugs 2017, 15, 342. [Google Scholar] [CrossRef]
- Gomes, D.L.; Martins, C.A.; Faria, L.M.; Santos, L.S.; Santos, C.S.; Sabbadini, P.S.; Souza, M.C.; Alves, G.B.; Rosa, A.C.; Nagao, P.E. Corynebacterium diphtheriae as an emerging pathogen in nephrostomy catheter-related infection: Evaluation of traits associated with bacterial virulence. J. Med. Microbiol. 2009, 58, 1419–1427. [Google Scholar] [CrossRef]
- Lieten, S.; Schelfaut, D.; Wissing, K.M.; Geers, C.; Tielemans, C. Alkaline-encrusted pyelitis and cystitis: An easily missed and life-threatening urinary infection. Case Rep. 2011, 2011, bcr1220103613. [Google Scholar] [CrossRef]
- Sánchez-Martín, F.; López-Martínez, J.; Kanashiro-Azabache, A.; Moncada, E.; Angerri-Feu, O.; Millán-Rodríguez, F.; Villavicencio-Mavrich, H. Corinebacterium urealyticum: Increased incidence of infection and encrusted uropathy. Actas Urológicas Españolas 2016, 40, 102–107. [Google Scholar] [CrossRef]
- Netzel, T. Prostatitis associated with Corynebacterium glucuronolyticum. Clin. Microbiol. Newslett. 2002, 24, 94–95. [Google Scholar] [CrossRef]
- Magri, V.; Trinchieri, A.; Ceriani, I.; Marras, E.; Perletti, G. Eradication of unusual pathogens by combination pharmacological therapy is paralleled by improvement of signs and symptoms of chronic prostatitis syndrome. Arch. Ital. Urol. Androl. 2007, 79, 93. [Google Scholar] [PubMed]
- Ivanov, I.B.; Kuzmin, M.D.; Gritsenko, V.A. Microflora of the seminal fluid of healthy men and men suffering from chronic prostatitis syndrome. Int. J. Androl. 2009, 32, 462–467. [Google Scholar] [CrossRef] [PubMed]
- Türk, S.; Punab, M.; Mändar, R. Antimicrobial susceptibility patterns of coryneform bacteria isolated from semen. Open Infect. Dis. J. 2009, 3. [Google Scholar] [CrossRef]
- Türk, S.; Mazzoli, S.; Štšepetova, J.; Kuznetsova, J.; Mändar, R. Coryneform bacteria in human semen: Inter-assay variability in species composition detection and biofilm production ability. Microb. Ecol. Health Dis. 2014, 25, 22701. [Google Scholar] [CrossRef]
- Lummus, W.E.; Thompson, I. Prostatitis. Emerg. Med. Clin. N. Am. 2001, 19, 691–707. [Google Scholar] [CrossRef]
- Khan, F.U.; Ihsan, A.U.; Khan, H.U.; Jana, R.; Wazir, J.; Khongorzul, P.; Waqar, M.; Zhou, X. Comprehensive overview of prostatitis. Biomed. Pharmacother. 2017, 94, 1064–1076. [Google Scholar] [CrossRef]
- Davis, N.G.; Silberman, M. Bacterial Acute Prostatitis. StatPearls: St. Petersburg, FI, USA, 2019. [Google Scholar]
- Delcaru, C.; Alexandru, I.; Podgoreanu, P.; Grosu, M.; Stavropoulos, E.; Chifiriuc, M.C.; Lazar, V. Microbial biofilms in urinary tract infections and prostatitis: Etiology, pathogenicity, and combating strategies. Pathogens 2016, 5, 65. [Google Scholar] [CrossRef]
- Ram, Y.; Dellus-Gur, E.; Bibi, M.; Karkare, K.; Obolski, U.; Feldman, M.W.; Cooper, T.F.; Berman, J.; Hadany, L. Predicting microbial growth in a mixed culture from growth curve data. Proc. Natl. Acad. Sci. USA 2019, 116, 14698–14707. [Google Scholar] [CrossRef]
- Walter, B.; Hänssler, E.; Kalinowski, J.; Burkovski, A. Nitrogen metabolism and nitrogen control in corynebacteria: Variations of a common theme. J. Mol. Microbiol. Biotechnol. 2007, 12, 131–138. [Google Scholar] [CrossRef]
- Nervig, R.M.; Kadis, S. Effect of hydroxamic acids on growth and urease activity in Corynebacterium renale. Can. J. Microbiol. 1976, 22, 544–551. [Google Scholar] [CrossRef]
- Kappaun, K.; Piovesan, A.R.; Carlini, C.R.; Ligabue-Braun, R. Ureases: Historical aspects, catalytic, and non-catalytic properties—A review. J. Adv. Res. 2018, 13, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Rinaldo, S.; Giardina, G.; Mantoni, F.; Paone, A.; Cutruzzolà, F. Beyond nitrogen metabolism: Nitric oxide, cyclic-di-GMP and bacterial biofilms. FEMS Microbiol. Lett. 2018, 365, fny029. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Corynebacterium urealyticum ATCC 43042 | Corynebacterium urealyticum ATCC 43043 | Corynebacterium urealyticum ATCC 43044 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| a 0.50 | a 0.25 | b C+ | a 0.50 | a 0.25 | b C+ | a 0.50 | a 0.25 | b C+ | |
| Goodness of Fit R2 | 0.9990 | 0.9997 | 0.9983 | 0.9992 | 0.9992 | 0.9994 | 0.9974 | 0.9973 | 0.9984 |
| Ytop (CFU/mL) at 72 h | 9.38 × 108 | 9.54 × 108 | 9.78 × 108 | 8.19 × 108 | 8.43 × 108 | 9.78 × 108 | 9.48 × 108 | 9.53 × 108 | 9.58 × 108 |
| Ytop ± SD c (CFU/mL) at 72 h | 1.01 × 106 | 1.74 × 106 | 4.56 × 106 | 6.39 × 106 | 3.93 × 106 | 3.58 × 106 | 2.91 × 106 | 2.90 × 106 | 2.23 × 106 |
| Inhibition (%) | 4.09 | 2.45 | / | 16.26 | 13.80 | / | 1.04 | 0.52 | / |
| XCFU50 (h) | 41.85 | 40.52 | 38.22 | 58.50 | 52.28 | 47.12 | 20.31 | 20.02 | 19.22 |
| XCFU50 ± S.D. c (h) | 0.02 | 0.05 | 0.15 | 0.12 | 0.09 | 0.09 | 0.01 | 0.01 | 0.07 |
| ∆XCFU50 (h) | 3.63 | 2.30 | / | 11.38 | 5.16 | / | 1.09 | 0.80 | / |
| C. urealyticum strain | Growth | Biofilm Formation | LNCaP Adhesion | HeLa Adhesion | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| a C+ | AHA | a C+ | AHA | a C+ | AHA | a C+ | AHA | |||||
| b 0.25 | b 0.50 | b 0.25 | b 0.50 | b 0.50 | b 0.25 | b 0.25 | b 0.50 | |||||
| ATCC 43042 | ++ | ++ | ++ | ++ | ++ | ++ | +++ | ++ | ++ | + | ++ | ++ |
| ATCC 43043 | + | +++ | +++ | ++ | ++ | ++ | + | ++ | ++ | + | ++ | ++ |
| ATCC 43044 | +++ | + | + | +++ | ++ | ++ | +++ | ++ | ++ | + | ++ | ++ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nicolosi, D.; Genovese, C.; Cutuli, M.A.; D’Angeli, F.; Pietrangelo, L.; Davinelli, S.; Petronio Petronio, G.; Di Marco, R. Preliminary In Vitro Studies on Corynebacterium urealyticum Pathogenetic Mechanisms, a Possible Candidate for Chronic Idiopathic Prostatitis? Microorganisms 2020, 8, 463. https://doi.org/10.3390/microorganisms8040463
Nicolosi D, Genovese C, Cutuli MA, D’Angeli F, Pietrangelo L, Davinelli S, Petronio Petronio G, Di Marco R. Preliminary In Vitro Studies on Corynebacterium urealyticum Pathogenetic Mechanisms, a Possible Candidate for Chronic Idiopathic Prostatitis? Microorganisms. 2020; 8(4):463. https://doi.org/10.3390/microorganisms8040463
Chicago/Turabian StyleNicolosi, Daria, Carlo Genovese, Marco Alfio Cutuli, Floriana D’Angeli, Laura Pietrangelo, Sergio Davinelli, Giulio Petronio Petronio, and Roberto Di Marco. 2020. "Preliminary In Vitro Studies on Corynebacterium urealyticum Pathogenetic Mechanisms, a Possible Candidate for Chronic Idiopathic Prostatitis?" Microorganisms 8, no. 4: 463. https://doi.org/10.3390/microorganisms8040463
APA StyleNicolosi, D., Genovese, C., Cutuli, M. A., D’Angeli, F., Pietrangelo, L., Davinelli, S., Petronio Petronio, G., & Di Marco, R. (2020). Preliminary In Vitro Studies on Corynebacterium urealyticum Pathogenetic Mechanisms, a Possible Candidate for Chronic Idiopathic Prostatitis? Microorganisms, 8(4), 463. https://doi.org/10.3390/microorganisms8040463