Galleria mellonella for the Evaluation of Antifungal Efficacy against Medically Important Fungi, a Narrative Review




Abstract
1. Introduction
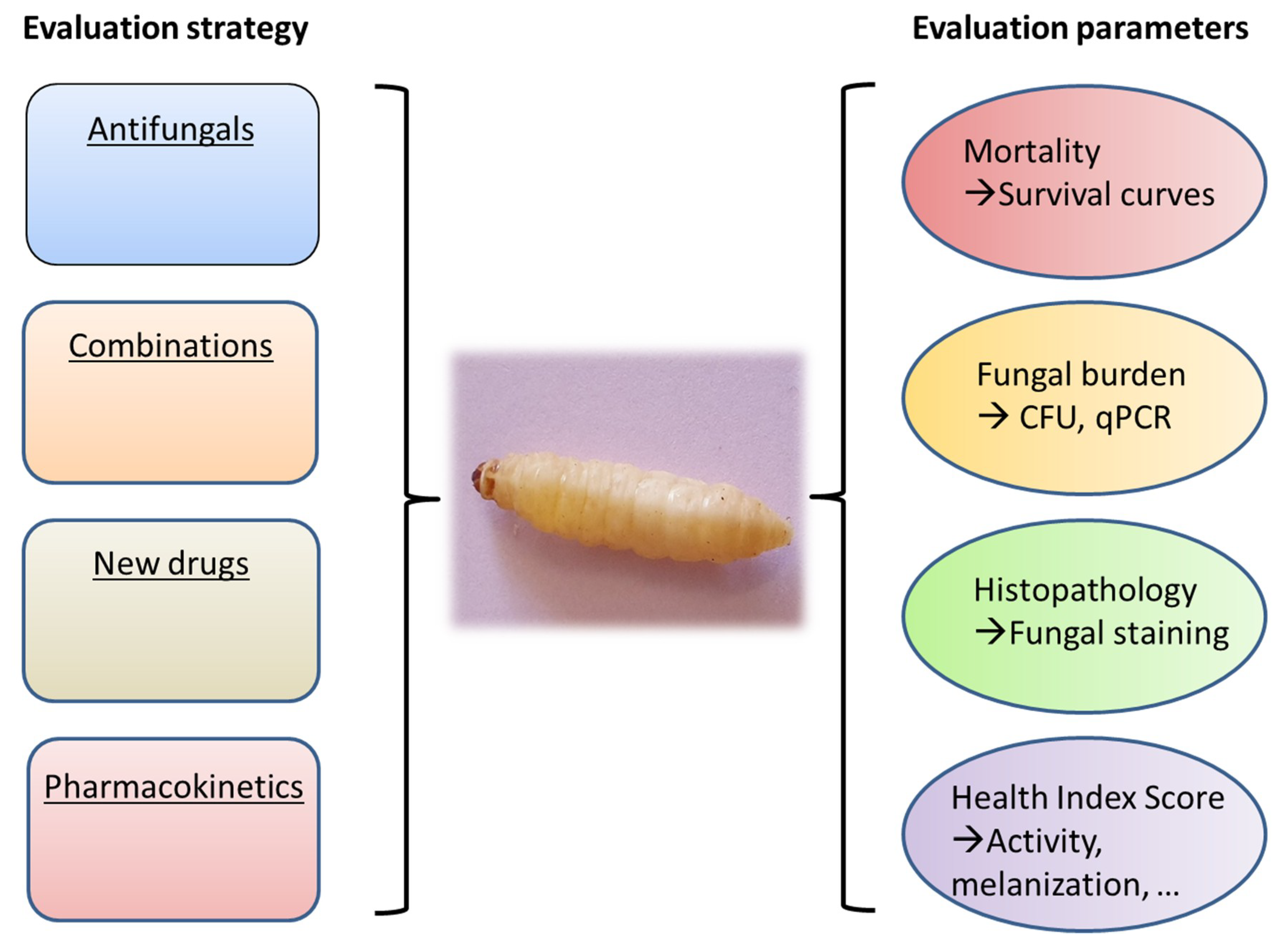
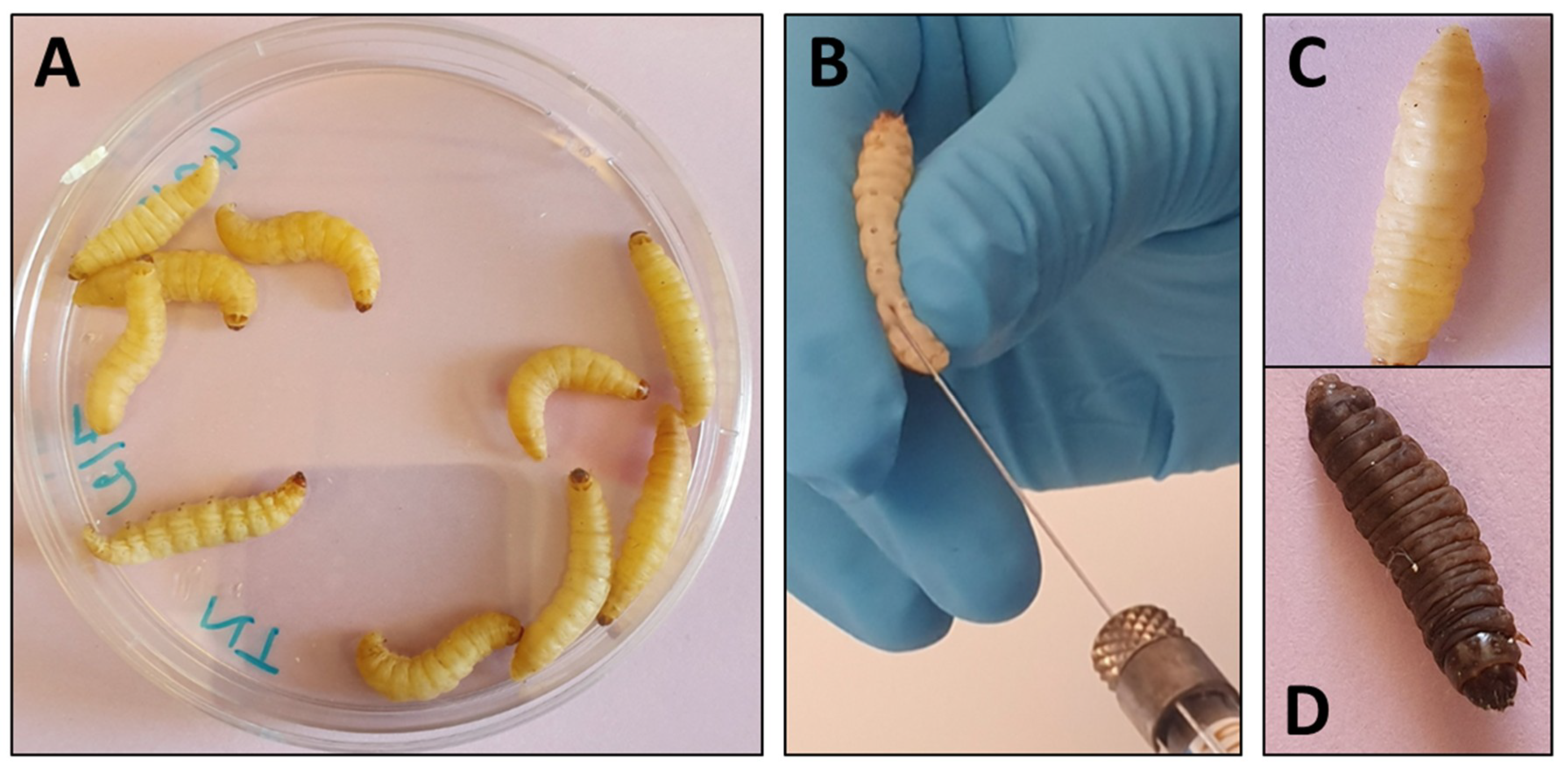
2. Methods and Endpoints Used for Evaluation of Antifungal Activity in Galleria mellonella
3. Evaluation of Antifungal Efficacy in Galleria mellonella
3.1. Candida spp.
3.1.1. Conventional Antifungals
3.1.2. Combinations
3.1.3. New Drugs
3.2. Cryptococcus spp.
3.3. Trichosporon spp.
3.4. Aspergillus spp.
3.5. Mucorales
3.6. Madurella Mycetomatis
4. Pharmacokinetics of Antifungals in G. mellonella
5. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Hohl, T.M. Overview of vertebrate animal models of fungal infection. J. Immunol. Methods 2014, 410, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Arvanitis, M.; Glavis-Bloom, J.; Mylonakis, E. Invertebrate models of fungal infection. Biochim. Biophys. Acta 2013, 1832, 1378–1383. [Google Scholar] [CrossRef]
- Chamilos, G.; Lionakis, M.S.; Lewis, R.E.; Kontoyiannis, D.P. Role of mini-host models in the study of medically important fungi. Lancet Infect. Dis. 2007, 7, 42–55. [Google Scholar] [CrossRef]
- Canteri de Souza, P.; Custodio Caloni, C.; Wilson, D.; Sergio Almeida, R. An invertebrate host to study fungal infections, mycotoxins and antifungal drugs: Tenebrio molitor. J. Fungi (Basel) 2018, 4, 125. [Google Scholar] [CrossRef] [PubMed]
- Pukkila-Worley, R.; Holson, E.; Wagner, F.; Mylonakis, E. Antifungal drug discovery through the study of invertebrate model hosts. Curr. Med. Chem. 2009, 16, 1588–1595. [Google Scholar] [CrossRef] [PubMed]
- Andrea, A.; Krogfelt, K.A.; Jenssen, H. Methods and challenges of using the Greater Wax moth (Galleria mellonella) as a model organism in antimicrobial compound discovery. Microorganisms 2019, 7, 85. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.J.; Loh, J.M.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef]
- Ribeiro, C.; Brehelin, M. Insect haemocytes: What type of cell is that? J. Insect Physiol. 2006, 52, 417–429. [Google Scholar] [CrossRef]
- Wojda, I. Immunity of the greater wax moth Galleria mellonella. Insect Sci. 2017, 24, 342–357. [Google Scholar] [CrossRef]
- Kavanagh, K.; Sheehan, G. The use of Galleria mellonella larvae to identify novel antimicrobial agents against fungal species of medical interest. J. Fungi (Basel) 2018, 4, 113. [Google Scholar] [CrossRef]
- Lionakis, M.S. Drosophila and Galleria insect model hosts: New tools for the study of fungal virulence, pharmacology and immunology. Virulence 2011, 2, 521–527. [Google Scholar] [CrossRef] [PubMed]
- Champion, O.L.; Titball, R.W.; Bates, S. Standardization of G. mellonella larvae to provide reliable and reproducible results in the study of fungal pathogens. J. Fungi (Basel) 2018, 4, 108. [Google Scholar] [CrossRef] [PubMed]
- Ames, L.; Duxbury, S.; Pawlowska, B.; Ho, H.L.; Haynes, K.; Bates, S. Galleria mellonella as a host model to study Candida glabrata virulence and antifungal efficacy. Virulence 2017, 8, 1909–1917. [Google Scholar] [CrossRef] [PubMed]
- Binder, U.; Aigner, M.; Risslegger, B.; Hortnagl, C.; Lass-Florl, C.; Lackner, M. Minimal inhibitory concentration (MIC)-phenomena in Candida albicans and their impact on the diagnosis of antifungal resistance. J. Fungi (Basel) 2019, 5, 83. [Google Scholar] [CrossRef]
- Forastiero, A.; Mesa-Arango, A.C.; Alastruey-Izquierdo, A.; Alcazar-Fuoli, L.; Bernal-Martinez, L.; Pelaez, T.; Lopez, J.F.; Grimalt, J.O.; Gomez-Lopez, A.; Cuesta, I.; et al. Candida tropicalis antifungal cross-resistance is related to different azole target (Erg11p) modifications. Antimicrob. Agents Chemother. 2013, 57, 4769–4781. [Google Scholar] [CrossRef]
- Fuchs, B.B.; Li, Y.; Li, D.; Johnston, T.; Hendricks, G.; Li, G.; Rajamuthiah, R.; Mylonakis, E. Micafungin elicits an immunomodulatory effect in Galleria mellonella and mice. Mycopathologia 2016, 181, 17–25. [Google Scholar] [CrossRef]
- Kelly, J.; Kavanagh, K. Caspofungin primes the immune response of the larvae of Galleria mellonella and induces a non-specific antimicrobial response. J. Med. Microbiol. 2011, 60, 189–196. [Google Scholar] [CrossRef]
- Li, D.D.; Deng, L.; Hu, G.H.; Zhao, L.X.; Hu, D.D.; Jiang, Y.Y.; Wang, Y. Using Galleria mellonella-Candida albicans infection model to evaluate antifungal agents. Biol. Pharm. Bull. 2013, 36, 1482–1487. [Google Scholar] [CrossRef]
- Mesa-Arango, A.C.; Forastiero, A.; Bernal-Martinez, L.; Cuenca-Estrella, M.; Mellado, E.; Zaragoza, O. The non-mammalian host Galleria mellonella can be used to study the virulence of the fungal pathogen Candida tropicalis and the efficacy of antifungal drugs during infection by this pathogenic yeast. Med. Mycol. 2013, 51, 461–472. [Google Scholar] [CrossRef]
- Morio, F.; Lombardi, L.; Binder, U.; Loge, C.; Robert, E.; Graessle, D.; Bodin, M.; Lass-Florl, C.; Butler, G.; Le Pape, P. Precise genome editing using a CRISPR-Cas9 method highlights the role of CoERG11 amino acid substitutions in azole resistance in Candida orthopsilosis. J. Antimicrob. Chemother. 2019, 74, 2230–2238. [Google Scholar] [CrossRef]
- Scorzoni, L.; de Lucas, M.P.; Mesa-Arango, A.C.; Fusco-Almeida, A.M.; Lozano, E.; Cuenca-Estrella, M.; Mendes-Giannini, M.J.; Zaragoza, O. Antifungal efficacy during Candida krusei infection in non-conventional models correlates with the yeast in vitro susceptibility profile. PLoS ONE 2013, 8, e60047. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.N.; Campos-Silva, R.; Ramos, L.S.; Trentin, D.S.; Macedo, A.J.; Branquinha, M.H.; Santos, A.L.S. Virulence of Candida haemulonii complex in Galleria mellonella and efficacy of classical antifungal drugs: A comparative study with other clinically relevant non-albicans Candida species. Fems Yeast Res. 2018, 18, foy082. [Google Scholar] [CrossRef] [PubMed]
- Souza, A.C.; Fuchs, B.B.; Pinhati, H.M.; Siqueira, R.A.; Hagen, F.; Meis, J.F.; Mylonakis, E.; Colombo, A.L. Candida parapsilosis resistance to fluconazole: Molecular mechanisms and in vivo impact in infected Galleria mellonella larvae. Antimicrob. Agents Chemother. 2015, 59, 6581–6587. [Google Scholar] [CrossRef] [PubMed]
- Ajdidi, A.; Sheehan, G.; Abu Elteen, K.; Kavanagh, K. Assessment of the in vitro and in vivo activity of atorvastatin against Candida albicans. J. Med. Microbiol. 2019, 68, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Browne, N.; Hackenberg, F.; Streciwilk, W.; Tacke, M.; Kavanagh, K. Assessment of in vivo antimicrobial activity of the carbene silver(I) acetate derivative SBC3 using Galleria mellonella larvae. Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 2014, 27, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.I.B.; Lopes, L.F.F.; de Camargo Ribeiro, F.; de Sa, N.P.; Lino, C.I.; Tharmalingam, N.; de Oliveira, R.B.; Rosa, C.A.; Mylonakis, E.; Fuchs, B.B.; et al. Anti-Candida albicans activity of thiazolylhydrazone derivatives in invertebrate and murine models. J. Fungi (Basel) 2018, 4, 134. [Google Scholar] [CrossRef]
- Favre-Godal, Q.; Dorsaz, S.; Queiroz, E.F.; Conan, C.; Marcourt, L.; Wardojo, B.P.; Voinesco, F.; Buchwalder, A.; Gindro, K.; Sanglard, D.; et al. Comprehensive approach for the detection of antifungal compounds using a susceptible strain of Candida albicans and confirmation of in vivo activity with the Galleria mellonella model. Phytochemistry 2014, 105, 68–78. [Google Scholar] [CrossRef]
- Nile, C.; Falleni, M.; Cirasola, D.; Alghamdi, A.; Anderson, O.F.; Delaney, C.; Ramage, G.; Ottaviano, E.; Tosi, D.; Bulfamante, G.; et al. Repurposing pilocarpine hydrochloride for treatment of Candida albicans infections. mSphere 2019, 4, e00689. [Google Scholar] [CrossRef]
- Osmanov, A.; Wise, A.; Denning, D.W. In vitro and in vivo efficacy of miramistin against drug-resistant fungi. J. Med. Microbiol. 2019, 68, 1047–1052. [Google Scholar] [CrossRef]
- Rossoni, R.D.; de Barros, P.P.; Lopes, L.; Ribeiro, F.C.; Nakatsuka, T.; Kasaba, H.; Junqueira, J.C. Effects of surface pre-reacted glass-ionomer (S-PRG) eluate on Candida spp.: Antifungal activity, anti-biofilm properties, and protective effects on Galleria mellonella against C. albicans infection. Biofouling 2019, 35, 997–1006. [Google Scholar] [CrossRef]
- Rowan, R.; Moran, C.; McCann, M.; Kavanagh, K. Use of Galleria mellonella larvae to evaluate the in vivo anti-fungal activity of [Ag2(mal)(phen)3]. Biometals Int. J. Role Met. Ions Biol. Biochem. Med. 2009, 22, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Sa, N.P.; Lima, C.M.; JR, A.D.S.; Costa, M.C.; de Barros, P.P.; Junqueira, J.C.; Vaz, J.A.; Oliveira, R.B.; Fuchs, B.B.; Mylonakis, E.; et al. A phenylthiazole derivative demonstrates efficacy on treatment of the cryptococcosis & candidiasis in animal models. Future Sci. Oa 2018, 4, dFso305. [Google Scholar] [CrossRef]
- Spadari, C.C.; de Bastiani, F.; Lopes, L.B.; Ishida, K. Alginate nanoparticles as non-toxic delivery system for miltefosine in the treatment of candidiasis and cryptococcosis. Int. J. Nanomed. 2019, 14, 5187–5199. [Google Scholar] [CrossRef]
- Staniszewska, M.; Gizinska, M.; Mikulak, E.; Adamus, K.; Koronkiewicz, M.; Lukowska-Chojnacka, E. New 1,5 and 2,5-disubstituted tetrazoles-dependent activity towards surface barrier of Candida albicans. Eur. J. Med. Chem. 2018, 145, 124–139. [Google Scholar] [CrossRef] [PubMed]
- Cowen, L.E.; Singh, S.D.; Kohler, J.R.; Collins, C.; Zaas, A.K.; Schell, W.A.; Aziz, H.; Mylonakis, E.; Perfect, J.R.; Whitesell, L.; et al. Harnessing Hsp90 function as a powerful, broadly effective therapeutic strategy for fungal infectious disease. Proc. Natl. Acad. Sci. USA 2009, 106, 2818–2823. [Google Scholar] [CrossRef]
- Gu, W.; Yu, Q.; Yu, C.; Sun, S. In vivo activity of fluconazole/tetracycline combinations in Galleria mellonella with resistant Candida albicans infection. J. Glob. Antimicrob. Resist. 2018, 13, 74–80. [Google Scholar] [CrossRef]
- Li, X.; Wu, X.; Gao, Y.; Hao, L. Synergistic effects and mechanisms of combined treatment with harmine hydrochloride and azoles for resistant Candida albicans. Front. Microbiol. 2019, 10, 2295. [Google Scholar] [CrossRef]
- Li, X.; Zhao, Y.; Huang, X.; Yu, C.; Yang, Y.; Sun, S. Ambroxol hydrochloride combined with fluconazole reverses the resistance of Candida albicans to fluconazole. Front. Cell Infect. Microbiol. 2017, 7, 124. [Google Scholar] [CrossRef]
- Li, Y.; Jiao, P.; Li, Y.; Gong, Y.; Chen, X.; Sun, S. The synergistic antifungal effect and potential mechanism of D-penicillamine combined with fluconazole against Candida albicans. Front. Microbiol. 2019, 10, 2853. [Google Scholar] [CrossRef]
- Liu, X.; Li, T.; Wang, D.; Yang, Y.; Sun, W.; Liu, J.; Sun, S. Synergistic antifungal effect of fluconazole combined with licofelone against resistant Candida albicans. Front. Microbiol. 2017, 8, 2101. [Google Scholar] [CrossRef]
- Lu, M.; Yang, X.; Yu, C.; Gong, Y.; Yuan, L.; Hao, L.; Sun, S. Linezolid in combination with azoles induced synergistic effects against Candida albicans and protected Galleria mellonella against experimental candidiasis. Front. Microbiol. 2019, 9, 3142. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Yu, C.; Cui, X.; Shi, J.; Yuan, L.; Sun, S. Gentamicin synergises with azoles against drug-resistant Candida albicans. Int. J. Antimicrob. Agents 2018, 51, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Wang, D.; Yu, C.; Huang, X.; Li, X.; Sun, S. Strong synergism of dexamethasone in combination with fluconazole against resistant Candida albicans mediated by inhibiting drug efflux and reducing virulence. Int. J. Antimicrob. Agents 2017, 50, 399–405. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Yan, H.; Lu, M.; Wang, D.; Sun, S. Antifungal activity of ribavirin used alone or in combination with fluconazole against Candida albicans is mediated by reduced virulence. Int. J. Antimicrob. Agents 2020, 55, 105804. [Google Scholar] [CrossRef]
- de Castro Spadari, C.; da Silva de Bastiani, F.W.M.; Pisani, P.B.B.; de Azevedo Melo, A.S.; Ishida, K. Efficacy of voriconazole in vitro and in invertebrate model of cryptococcosiss. Arch. Microbiol. 2019. [Google Scholar] [CrossRef]
- de Sa, N.P.; de Barros, P.P.; Junqueira, J.C.; Vaz, J.A.; de Oliveira, R.B.; Rosa, C.A.; Santos, D.A.; Johann, S. Thiazole derivatives act on virulence factors of Cryptococcus spp. Med. Mycol. 2019, 57, 84–91. [Google Scholar] [CrossRef]
- Mylonakis, E.; Moreno, R.; El Khoury, J.B.; Idnurm, A.; Heitman, J.; Calderwood, S.B.; Ausubel, F.M.; Diener, A. Galleria mellonella as a model system to study Cryptococcus neoformans pathogenesis. Infect. Immun. 2005, 73, 3842–3850. [Google Scholar] [CrossRef]
- Palanco, A.C.; Lacorte Singulani, J.; Costa-Orlandi, C.B.; Gullo, F.P.; Strohmayer Lourencetti, N.M.; Gomes, P.C.; Ayusso, G.M.; Dutra, L.A.; Silva Bolzani, V.D.; Regasini, L.O.; et al. Activity of 3’-hydroxychalcone against Cryptococcus gattii and toxicity, and efficacy in alternative animal models. Future Microbiol. 2017, 12, 1123–1134. [Google Scholar] [CrossRef]
- Sangalli-Leite, F.; Scorzoni, L.; Alves de Paula e Silva, A.C.; da Silva, J.F.; de Oliveira, H.C.; de Lacorte Singulani, J.; Gullo, F.P.; Moraes da Silva, R.; Regasini, L.O.; Siqueira da Silva, D.H.; et al. Synergistic effect of pedalitin and amphotericin B against Cryptococcus neoformans by in vitro and in vivo evaluation. Int. J. Antimicrob. Agents 2016, 48, 504–511. [Google Scholar] [CrossRef]
- Singulani, J.L.; Galeane, M.C.; Ramos, M.D.; Gomes, P.C.; Dos Santos, C.T.; de Souza, B.M.; Palma, M.S.; Fusco Almeida, A.M.; Mendes Giannini, M.J.S. Antifungal activity, toxicity, and membranolytic action of a mastoparan analog peptide. Front. Cell Infect. Microbiol. 2019, 9, 419. [Google Scholar] [CrossRef]
- Vu, K.; Gelli, A. Astemizole and an analogue promote fungicidal activity of fluconazole against Cryptococcus neoformans var. grubii and Cryptococcus gattii. Med. Mycol. 2010, 48, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Blatzer, M.; Blum, G.; Jukic, E.; Posch, W.; Gruber, P.; Nagl, M.; Binder, U.; Maurer, E.; Sarg, B.; Lindner, H.; et al. Blocking Hsp70 enhances the efficiency of amphotericin B treatment against resistant Aspergillus terreus strains. Antimicrob. Agents Chemother. 2015, 59, 3778–3788. [Google Scholar] [CrossRef] [PubMed]
- Forastiero, A.; Bernal-Martinez, L.; Mellado, E.; Cendejas, E.; Gomez-Lopez, A. In vivo efficacy of voriconazole and posaconazole therapy in a novel invertebrate model of Aspergillus fumigatus infection. Int. J. Antimicrob. Agents 2015, 46, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rubio, R.; Alcazar-Fuoli, L.; Monteiro, M.C.; Monzon, S.; Cuesta, I.; Pelaez, T.; Mellado, E. Insight into the significance of Aspergillus fumigatus cyp51A polymorphisms. Antimicrob. Agents Chemother. 2018, 62, e00241. [Google Scholar] [CrossRef]
- Glampedakis, E.; Coste, A.T.; Aruanno, M.; Bachmann, D.; Delarze, E.; Erard, V.; Lamoth, F. Efficacy of antifungal monotherapies and combinations against Aspergillus calidoustus. Antimicrob. Agents Chemother. 2018, 62, e01137. [Google Scholar] [CrossRef]
- Maurer, E.; Browne, N.; Surlis, C.; Jukic, E.; Moser, P.; Kavanagh, K.; Lass-Florl, C.; Binder, U. Galleria mellonella as a host model to study Aspergillus terreus virulence and amphotericin B resistance. Virulence 2015, 6, 591–598. [Google Scholar] [CrossRef]
- Ben Yaakov, D.; Rivkin, A.; Mircus, G.; Albert, N.; Dietl, A.M.; Kovalerchick, D.; Carmeli, S.; Haas, H.; Kontoyiannis, D.P.; Osherov, N. Identification and characterization of haemofungin, a novel antifungal compound that inhibits the final step of haem biosynthesis. J. Antimicrob. Chemother. 2016, 71, 946–952. [Google Scholar] [CrossRef]
- Ben Yaakov, D.; Shadkchan, Y.; Albert, N.; Kontoyiannis, D.P.; Osherov, N. The quinoline bromoquinol exhibits broad-spectrum antifungal activity and induces oxidative stress and apoptosis in Aspergillus fumigatus. J. Antimicrob. Chemother. 2017, 72, 2263–2272. [Google Scholar] [CrossRef]
- Liu, F.F.; Pu, L.; Zheng, Q.Q.; Zhang, Y.W.; Gao, R.S.; Xu, X.S.; Zhang, S.Z.; Lu, L. Calcium signaling mediates antifungal activity of triazole drugs in the Aspergilli. Fungal Genet. Biol. 2015, 81, 182–190. [Google Scholar] [CrossRef]
- Mircus, G.; Albert, N.; Ben-Yaakov, D.; Chikvashvili, D.; Shadkchan, Y.; Kontoyiannis, D.P.; Osherov, N. Identification and characterization of a novel family of selective antifungal compounds (CANBEFs) that interfere with fungal protein synthesis. Antimicrob. Agents Chemother. 2015, 59, 5631–5640. [Google Scholar] [CrossRef]
- Trevino-Rangel, R.J.; Villanueva-Lozano, H.; Mendez-Galomo, K.S.; Solis-Villegas, E.M.; Becerril-Garcia, M.A.; Montoya, A.M.; Robledo-Leal, E.R.; Gonzalez, G.M. In vivo evaluation of the antifungal activity of sertraline against Aspergillus fumigatus. J. Antimicrob. Chemother. 2019, 74, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Bastidas, R.J.; Shertz, C.A.; Lee, S.C.; Heitman, J.; Cardenas, M.E. Rapamycin exerts antifungal activity in vitro and in vivo against Mucor circinelloides via FKBP12-dependent inhibition of Tor. Eukaryot Cell 2012, 11, 270–281. [Google Scholar] [CrossRef] [PubMed]
- Macedo, D.; Leonardelli, F.; Dudiuk, C.; Vitale, R.G.; Del Valle, E.; Giusiano, G.; Gamarra, S.; Garcia-Effron, G. In vitro and in vivo evaluation of voriconazole-containing antifungal combinations against Mucorales using a Galleria mellonella model of mucormycosis. J. Fungi (Basel) 2019, 5, 5. [Google Scholar] [CrossRef] [PubMed]
- Maurer, E.; Hortnagl, C.; Lackner, M.; Grassle, D.; Naschberger, V.; Moser, P.; Segal, E.; Semis, M.; Lass-Florl, C.; Binder, U. Galleria mellonella as a model system to study virulence potential of mucormycetes and evaluation of antifungal treatment. Med. Mycol. 2019, 57, 351–362. [Google Scholar] [CrossRef] [PubMed]
- Eadie, K.; Parel, F.; Helvert-van Poppel, M.; Fahal, A.; van de Sande, W. Combining two antifungal agents does not enhance survival of Galleria mellonella larvae infected with Madurella mycetomatis. Trop. Med. Int. Health 2017, 22, 696–702. [Google Scholar] [CrossRef] [PubMed]
- Kloezen, W.; Parel, F.; Bruggemann, R.; Asouit, K.; Helvert-van Poppel, M.; Fahal, A.; Mouton, J.; van de Sande, W. Amphotericin B and terbinafine but not the azoles prolong survival in Galleria mellonella larvae infected with Madurella mycetomatis. Med. Mycol. 2018, 56, 469–478. [Google Scholar] [CrossRef]
- Lim, W.; Melse, Y.; Konings, M.; Phat Duong, H.; Eadie, K.; Laleu, B.; Perry, B.; Todd, M.H.; Ioset, J.R.; van de Sande, W.W.J. Addressing the most neglected diseases through an open research model: The discovery of fenarimols as novel drug candidates for eumycetoma. Plos Negl. Trop. Dis. 2018, 12, e0006437. [Google Scholar] [CrossRef]
- Marine, M.; Bom, V.L.; de Castro, P.A.; Winkelstroter, L.K.; Ramalho, L.N.; Brown, N.A.; Goldman, G.H. The development of animal infection models and antifungal efficacy assays against clinical isolates of Trichosporon asahii, T. asteroides and T. inkin. Virulence 2015, 6, 476–486. [Google Scholar] [CrossRef]
- de Lacorte Singulani, J.; Scorzoni, L.; de Paula, E.S.A.C.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J. Evaluation of the efficacy of antifungal drugs against Paracoccidioides brasiliensis and Paracoccidioides lutzii in a Galleria mellonella model. Int. J. Antimicrob. Agents 2016, 48, 292–297. [Google Scholar] [CrossRef]
- Singulani, J.L.; Scorzoni, L.; de Oliveira, H.C.; Marcos, C.M.; Assato, P.A.; Fusco-Almeida, A.M.; Mendes-Giannini, M.J.S. Applications of invertebrate animal models to dimorphic fungal infections. J. Fungi (Basel) 2018, 4, 118. [Google Scholar] [CrossRef]
- Jacobsen, I.D. Galleria mellonella as a model host to study virulence of Candida. Virulence 2014, 5, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Segal, E.; Frenkel, M. Experimental in vivo models of candidiasis. J. Fungi (Basel) 2018, 4, 21. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Yan, H.; Yu, C.; Yuan, L.; Sun, S. Proton pump inhibitors act synergistically with fluconazole against resistant Candida albicans. Sci. Rep. 2020, 10, 498. [Google Scholar] [CrossRef] [PubMed]
- Diekema, D.; Arbefeville, S.; Boyken, L.; Kroeger, J.; Pfaller, M. The changing epidemiology of healthcare-associated candidemia over three decades. Diagn. Microbiol. Infect. Dis. 2012, 73, 45–48. [Google Scholar] [CrossRef] [PubMed]
- MacCallum, D.M.; Desbois, A.P.; Coote, P.J. Enhanced efficacy of synergistic combinations of antimicrobial peptides with caspofungin versus Candida albicans in insect and murine models of systemic infection. Eur. J. Clin. Microbiol. Infect. Dis. Off. Publ. Eur. Soc. Clin. Microbiol. 2013, 32, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Cornely, O.A.; Bassetti, M.; Calandra, T.; Garbino, J.; Kullberg, B.J.; Lortholary, O.; Meersseman, W.; Akova, M.; Arendrup, M.C.; Arikan-Akdagli, S.; et al. ESCMID* guideline for the diagnosis and management of Candida diseases 2012: Non-neutropenic adult patients. Clin. Microbiol. Infect. 2012, 18, 19–37. [Google Scholar] [CrossRef] [PubMed]
- Bidaud, A.L.; Botterel, F.; Chowdhary, A.; Dannaoui, E. In vitro antifungal combination of flucytosine with amphotericin B, voriconazole, or micafungin against Candida auris shows no antagonism. Antimicrob. Agents Chemother. 2019. [Google Scholar] [CrossRef]
- Bidaud, A.L.; Djenontin, E.; Botterel, F.; Chowdhary, A.; Dannaoui, E. Colistin interacts synergistically with echinocandins against Candida auris. Int. J. Antimicrob. Agents 2020. [Google Scholar] [CrossRef]
- Apidianakis, Y.; Rahme, L.G.; Heitman, J.; Ausubel, F.M.; Calderwood, S.B.; Mylonakis, E. Challenge of Drosophila melanogaster with Cryptococcus neoformans and role of the innate immune response. Eukaryot Cell 2004, 3, 413–419. [Google Scholar] [CrossRef]
- Wang, Y.H.; Dong, H.H.; Zhao, F.; Wang, J.; Yan, F.; Jiang, Y.Y.; Jin, Y.S. The synthesis and synergistic antifungal effects of chalcones against drug resistant Candida albicans. Bioorg. Med. Chem. Lett. 2016, 26, 3098–3102. [Google Scholar] [CrossRef]
- de Almeida Junior, J.N.; Hennequin, C. Invasive Trichosporon infection: A systematic review on a re-emerging fungal pathogen. Front. Microbiol. 2016, 7, 1629. [Google Scholar] [CrossRef] [PubMed]
- Arendrup, M.C.; Boekhout, T.; Akova, M.; Meis, J.F.; Cornely, O.A.; Lortholary, O.; Escmid Efisg study group; Ecmm. ESCMID and ECMM joint clinical guidelines for the diagnosis and management of rare invasive yeast infections. Clin. Microbiol. Infect. 2014, 20, 76–98. [Google Scholar] [CrossRef] [PubMed]
- Alcazar-Fuoli, L.; Buitrago, M.; Gomez-Lopez, A.; Mellado, E. An alternative host model of a mixed fungal infection by azole susceptible and resistant Aspergillus spp strains. Virulence 2015, 6, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Mellado, E.; Alcazar-Fuoli, L.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Role of Aspergillus lentulus 14-alpha sterol demethylase (Cyp51A) in azole drug susceptibility. Antimicrob. Agents Chemother. 2011, 55, 5459–5468. [Google Scholar] [CrossRef]
- Lestrade, P.P.A.; Meis, J.F.; Melchers, W.J.G.; Verweij, P.E. Triazole resistance in Aspergillus fumigatus: Recent insights and challenges for patient management. Clin. Microbiol. Infect. 2019, 25, 799–806. [Google Scholar] [CrossRef]
- Rhein, J.; Morawski, B.M.; Hullsiek, K.H.; Nabeta, H.W.; Kiggundu, R.; Tugume, L.; Musubire, A.; Akampurira, A.; Smith, K.D.; Alhadab, A.; et al. Efficacy of adjunctive sertraline for the treatment of HIV-associated cryptococcal meningitis: An open-label dose-ranging study. Lancet Infect. Dis. 2016, 16, 809–818. [Google Scholar] [CrossRef]
- Cornely, O.A.; Alastruey-Izquierdo, A.; Arenz, D.; Chen, S.C.A.; Dannaoui, E.; Hochhegger, B.; Hoenigl, M.; Jensen, H.E.; Lagrou, K.; Lewis, R.E.; et al. Global guideline for the diagnosis and management of mucormycosis: An initiative of the European Confederation of Medical Mycology in cooperation with the Mycoses Study Group Education and Research Consortium. Lancet Infect. Dis. 2019, 19, e405–e421. [Google Scholar] [CrossRef]
- Prakash, H.; Chakrabarti, A. Global epidemiology of mucormycosis. J. Fungi (Basel) 2019, 5, 26. [Google Scholar] [CrossRef]
- Serris, A.; Danion, F.; Lanternier, F. Disease Entities in Mucormycosis. J. Fungi (Basel) 2019, 5, 23. [Google Scholar] [CrossRef]
- Bitar, D.; Van Cauteren, D.; Lanternier, F.; Dannaoui, E.; Che, D.; Dromer, F.; Desenclos, J.C.; Lortholary, O. Increasing incidence of zygomycosis (mucormycosis), France, 1997–2006. Emerg Infect. Dis. 2009, 15, 1395–1401. [Google Scholar] [CrossRef]
- Kaerger, K.; Schwartze, V.U.; Dolatabadi, S.; Nyilasi, I.; Kovacs, S.A.; Binder, U.; Papp, T.; Hoog, S.; Jacobsen, I.D.; Voigt, K. Adaptation to thermotolerance in Rhizopus coincides with virulence as revealed by avian and invertebrate infection models, phylogeny, physiological and metabolic flexibility. Virulence 2015, 6, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Semis, R.; Mendlovic, S.; Polacheck, I.; Segal, E. Activity of an intralipid formulation of nystatin in murine systemic candidiasis. Int. J. Antimicrob. Agents 2011, 38, 336–340. [Google Scholar] [CrossRef] [PubMed]
- Grizanova, E.V.; Krytsyna, T.I.; Surcova, V.S.; Dubovskiy, I.M. The role of midgut nonspecific esterase in the susceptibility of Galleria mellonella larvae to Bacillus thuringiensis. J. Invertebr. Pathol. 2019, 166, 107208. [Google Scholar] [CrossRef] [PubMed]
- Kloezen, W.; van Helvert-van Poppel, M.; Fahal, A.H.; van de Sande, W.W. A Madurella mycetomatis grain model in Galleria mellonella larvae. PLoS Negl. Trop. Dis. 2015, 9, e0003926. [Google Scholar] [CrossRef]
- Astvad, K.M.T.; Meletiadis, J.; Whalley, S.; Arendrup, M.C. Fluconazole pharmacokinetics in Galleria mellonella larvae and performance evaluation of a bioassay compared to liquid chromatography-tandem mass spectrometry for hemolymph specimens. Antimicrob. Agents Chemother. 2017, 61, e00895. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species | Antifungal | Dosage In Vivo (mg/kg) | In Vitro Phenotype | In vivo Efficacy (Gm) | Reference |
|---|---|---|---|---|---|
| C. albicans | FCZ | 3, 6, 12 | S | Yes | [13] |
| C. albicans | AMB | 2, 4 | S | Yes | [13] |
| C. albicans | CAS | 1, 2, 4 | S | Yes | [13] |
| C. albicans | FCZ | 1, 4, 16 | S | Yes | [18] |
| C. albicans | FCZ | 4 | R | No | [18] |
| C. albicans | AMB | 0.4, 1.6, 6.4 | S | Yes | [18] |
| C. albicans | 5FC | 1.25, 5, 20 | S | Yes | [18] |
| C. albicans | AMB | 1, 2, 4 | S | Yes | [21] |
| C. albicans | FCZ | 4, 12, 32, 64 | S | Yes | [21] |
| C. albicans | VRZ | 7.5, 10 | S | Yes | [21] |
| C. albicans | CAS | 1, 2, 4 | S | Yes | [21] |
| C. glabrata | FCZ | 3, 6, 12 | 32 | No | [13] |
| C. glabrata | AMB | 2, 4 | S | No at 2, Yes at 4 | [13] |
| C. glabrata | 5FC | 1, 2, 4 | S | No at 1, Yes at 2 and 4 | [13] |
| C. tropicalis | FCZ | 9 | S | Yes | [15] |
| C. tropicalis | FCZ | 9 | R | No | [15] |
| C. tropicalis | VRZ | 10 | S | Yes | [15] |
| C. tropicalis | VRZ | 10 | R | No | [15] |
| C. tropicalis | AMB | 3 | S | Yes | [15] |
| C. tropicalis | AMB | 3 | R | No | [15] |
| C. tropicalis | ANI | 10 | S | Yes | [15] |
| C. tropicalis | AMB | 1, 2, 4 | S | Yes | [19] |
| C. tropicalis | FCZ | ND | S | Yes high dose, No low dose | [19] |
| C. tropicalis | VRZ | ND | S | Yes high dose, No low dose | [19] |
| C. tropicalis | CAS | 1, 2, 4 | S | Yes | [19] |
| C. krusei | AMB | 1, 2, 4 | S | No at 1 and 2, Yes at 4 | [21] |
| C. krusei | FCZ | 4, 12, 32, 64 | R | No | [21] |
| C. krusei | VRZ | 7.5, 10 | S | No at 7.5, Yes at 10 | [21] |
| C. krusei | CAS | 1, 2, 4 | S | No at 1 and 2, Yes at 4 | [21] |
| C. orthopsilosis | FCZ | 2, 10 | S | Yes | [20] |
| C. orthopsilosis | FCZ | 2, 10 | R | No | [20] |
| C. parapsilosis | FCZ | 14 | S | Yes | [23] |
| C. parapsilosis | FCZ | 14 | R | No | [23] |
| C. haemulonii | FCZ | 6, 12 | R | No | [22] |
| C. haemulonii | AMB | 2.5, 5 | R | No | [22] |
| C. haemulonii | CAS | 0,5, 1 | S | Yes | [22] |
| C. duobushaemulonii | FCZ | 6, 12 | R | No | [22] |
| C. duobushaemulonii | AMB | 2.5, 5 | R | No | [22] |
| C. duobushaemulonii | CAS | 0,5, 1 | S | Yes | [22] |
| C. tropicalis | FCZ | 6, 12 | S | Yes | [22] |
| C. tropicalis | AMB | 2.5, 5 | S | Yes | [22] |
| C. tropicalis | CAS | 0,5, 1 | S | Yes | [22] |
| C. krusei | FCZ | 6, 12 | R | No | [22] |
| C. krusei | AMB | 2.5, 5 | S | Yes | [22] |
| C. krusei | CAS | 0,5, 1 | S | Yes | [22] |
| C. lusitaniae | FCZ | 6, 12 | S | Yes | [22] |
| C. lusitaniae | AMB | 2.5, 5 | R | No at 2.5, Yes at 5 | [22] |
| C. lusitaniae | CAS | 0,5, 1 | S | Yes | [22] |
| Species | Drugs in Combination | Efficacy of the Combination | Reference | ||
|---|---|---|---|---|---|
| Partner #1 | Partner #2 | In Vitro (SYN) | In Vivo (Gm) | ||
| C. albicans | AMB | 5FC | ND | Yes | [18] |
| C. albicans (Razole) | FCZ | Linezolid | Yes | Yes | [41] |
| C. albicans (Razole) | ITZ | Linezolid | Yes | Yes | [41] |
| C. albicans (Razole) | VRZ | Linezolid | Yes | Yes | [41] |
| C. albicans (Razole) | FCZ | Gentamicin | Yes | Yes | [42] |
| C. albicans (Razole) | FCZ | Minocycline | ND | Yes | [36] |
| C. albicans (Razole) | FCZ | Doxycycline | ND | Yes | [36] |
| C. albicans | CAS | Colistin | Yes | Yes | [75] |
| C. albicans (Razole) | FCZ | Dexamethasone | Yes | Yes | [43] |
| C. albicans (Razole) | FCZ | Licofelone | Yes | Yes | [59] |
| C. albicans (Razole) | FCZ | D-penicillamine | Yes | Yes | [39] |
| C. albicans (Razole) | FCZ | Harmine | Yes | Yes | [37] |
| C. albicans (Razole) | FCZ | Ambroxol | Yes | Yes | [38] |
| C. albicans (Razole) | FCZ | Ribavirin | Yes | Yes | [44] |
| C. albicans (Razole) | FCZ | Proton-pump inhibitors | Yes | Yes | [73] |
| C. albicans | FCZ | Hsp90 inhibitors | Yes | Yes | [35] |
| Species | Antifungals (doses [mg/kg]) | Combination | Main Results | Reference |
|---|---|---|---|---|
| C. neoformans | AMB (1.5) FCZ (14) 5-FC (20) | Yes | AMB or FC alone prolonged survival, FCZ prolonged survival (NS) AMB+FC more effective then AMB alone | [47] |
| C. neoformans C. gattii | VRZ (10, 20) AMB (1, 10, 20) | No | VRZ increased survival and decreased fungal burden | [45] |
| C. neoformans C. gattii | MFS* | No | MFS increased survival for C. gattii, and decreased fungal burden for both species | [33] |
| C. neoformans | AST, A2 FCZ | Yes | FCZ+AST and FCZ+A2 increased survival in larvae infected with FCZ-susceptible isolate | [51] |
| C. gattii | 3′-hydroxychalcone (2, 80, 160) AMB (2) | No | No in vitro–in vivo correlation. 3- hydroxychalcone fungicidal in vitro but no efficacy in vivo in terms of survival and fungal burden | [48] |
| C. gattii C. neoformans | CHT (5, 10) FCZ (10) | No | CHT increased survival for C. gattii and C. neoformans infected larvae. Correlation with a murine model | [32] |
| C. neoformans C. gattii | Compound 3 (5, 10) FCZ (5) | No | Compound 3 increased survival of infected larvae. Efficacy similar to that of FCZ | [46] |
| C. neoformans | PED (6.25 to 200) AMB (0.5 to 4) | Yes | AMB or PED increased survival. Better efficacy of the combination Good correlation with the murine model | [49] |
| C. neoformans | MK58911 (10 to 100) AMB (4) FCZ (10) | Yes | MK58911 increased survival. No benefit of MK58911+ AMB and MK58911+FCZ compared to monotherapies | [50] |
| Species | Antifungals (doses [mg/kg]) | Combination | Main Results | Reference |
|---|---|---|---|---|
| A. lentulus | VRZ (10) | No | No efficacy of VRZ against A. lentulus (azole-resistant) compared to A. fumigatus in term of survival and fungal burden | [83] |
| A. calidoustus | VRZ (10) AMB (5) TBF (5) | Yes | AMB not superior to VRZ in vivo, in contrast to in vitro. TBF combined with VRZ better than monotherapies. Combination synergistic in vitro. | [55] |
| A. terreus | L-AMB (1.6, 16.6) | No | Efficacy of L-AMB against AMB-susceptible isolates and no efficacy against AMB-resistant isolates. L-AMB administration increased hemocyte density. | [56] |
| A. terreus | AMB (5) Hsp70 inhibitor | Yes | AMB+Hsp70 inhibitor decreased MIC in vitro and increased survival in larvae infected with AMB-resistant isolate | [52] |
| A. fumigatus | VRZ (1.25, 2.5, 10, 40, 80) PSZ | No | VRZ at 10 mg/kg improved survival against VRC-susceptible strains (MIC ≤ 1 mg/L) but not against VRZ-resistant strains (MIC = 4 mg/L). | [53] |
| A. fumigatus | VRZ (10) | No | VRZ increased survival of larvae infected by either WT and mutant (isolates with SNPs in CYP51A and moderately elevated MICs) although mortality rate was higher for mutants. | [54] |
| A. fumigatus | CAS (1.5) GdA | Yes | Combination therapy (GdA + CAS) improved survival compared to each monotherapy. Correlation with in vitro results | [35] |
| A. fumigatus | ITZ (100) EGTA | Yes | The calcium chelator EGTA was synergistic in vivo when combined with ITZ. | [59] |
| A. fumigatus | AMB (2) Haemofungin | No | In vitro (CLSI), combination HMG+AMB not synergistic in contrast to HMG+CAS Efficacy of haemofungin (5.7 mg/kg) similar to that of amphotericin B at 2 mg/kg | [57] |
| A. fumigatus | AMB (1, 2) BMQ | No | Similar efficacy of BMQ (8 mg/kg) and AMB in G. mellonella. No correlation with a murine model. | [58] |
| A. fumigatus | CANBEF-24 (1.8, to 14.4) | No | No in vivo efficacy despite in vitro activity. | [60] |
| A. fumigatus | Miramistin | No | In vivo efficacy of miramistin | [29] |
| A. fumigatus | VRZ (10) AMB (3) sertraline (3, 10) | No | Survival of 50% for AMB and VRZ and 25% for sertraline. Correlation with murine model. | [61] |
| Species | Antifungals (Doses [mg/kg]) | Combination | Main Results | Reference |
|---|---|---|---|---|
| 6 speciesa | AMB (1), VRZ (10), CAS (0.5), PSZ (10) | Yes | No efficacy of monotherapies except for CAS vs. R. microsporus VRZ+AMB and VRZ+CAS increased survival compared to AMB alone for R. microsporus but not for R. oryzae | [63] |
| 6 speciesb | AMB (15), CAS (15), PSZ (15), NYS-L (15) | No | NYS-L has the best efficacy except for R. arrhizus L-AMB has low efficacy except for L. corymbifera ISA not effective except for M. circinelloides | [64] |
| M. circinelloides | Rapamycin (33) | No | Rapamycin increased survival | [62] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jemel, S.; Guillot, J.; Kallel, K.; Botterel, F.; Dannaoui, E. Galleria mellonella for the Evaluation of Antifungal Efficacy against Medically Important Fungi, a Narrative Review. Microorganisms 2020, 8, 390. https://doi.org/10.3390/microorganisms8030390
Jemel S, Guillot J, Kallel K, Botterel F, Dannaoui E. Galleria mellonella for the Evaluation of Antifungal Efficacy against Medically Important Fungi, a Narrative Review. Microorganisms. 2020; 8(3):390. https://doi.org/10.3390/microorganisms8030390
Chicago/Turabian StyleJemel, Sana, Jacques Guillot, Kalthoum Kallel, Françoise Botterel, and Eric Dannaoui. 2020. "Galleria mellonella for the Evaluation of Antifungal Efficacy against Medically Important Fungi, a Narrative Review" Microorganisms 8, no. 3: 390. https://doi.org/10.3390/microorganisms8030390
APA StyleJemel, S., Guillot, J., Kallel, K., Botterel, F., & Dannaoui, E. (2020). Galleria mellonella for the Evaluation of Antifungal Efficacy against Medically Important Fungi, a Narrative Review. Microorganisms, 8(3), 390. https://doi.org/10.3390/microorganisms8030390