Human Cytomegalovirus (HCMV) Reactivation in the Mammary Gland Induces a Proinflammatory Cytokine Shift in Breast Milk

Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Proximity Extension Assay (PEA)
2.3. Enzyme-Linked Immunosorbent Assay (ELISA) for Cytokine Quantification
2.4. DNA Extraction and Determination of DNA Viral Load via qPCR
2.5. Statistics
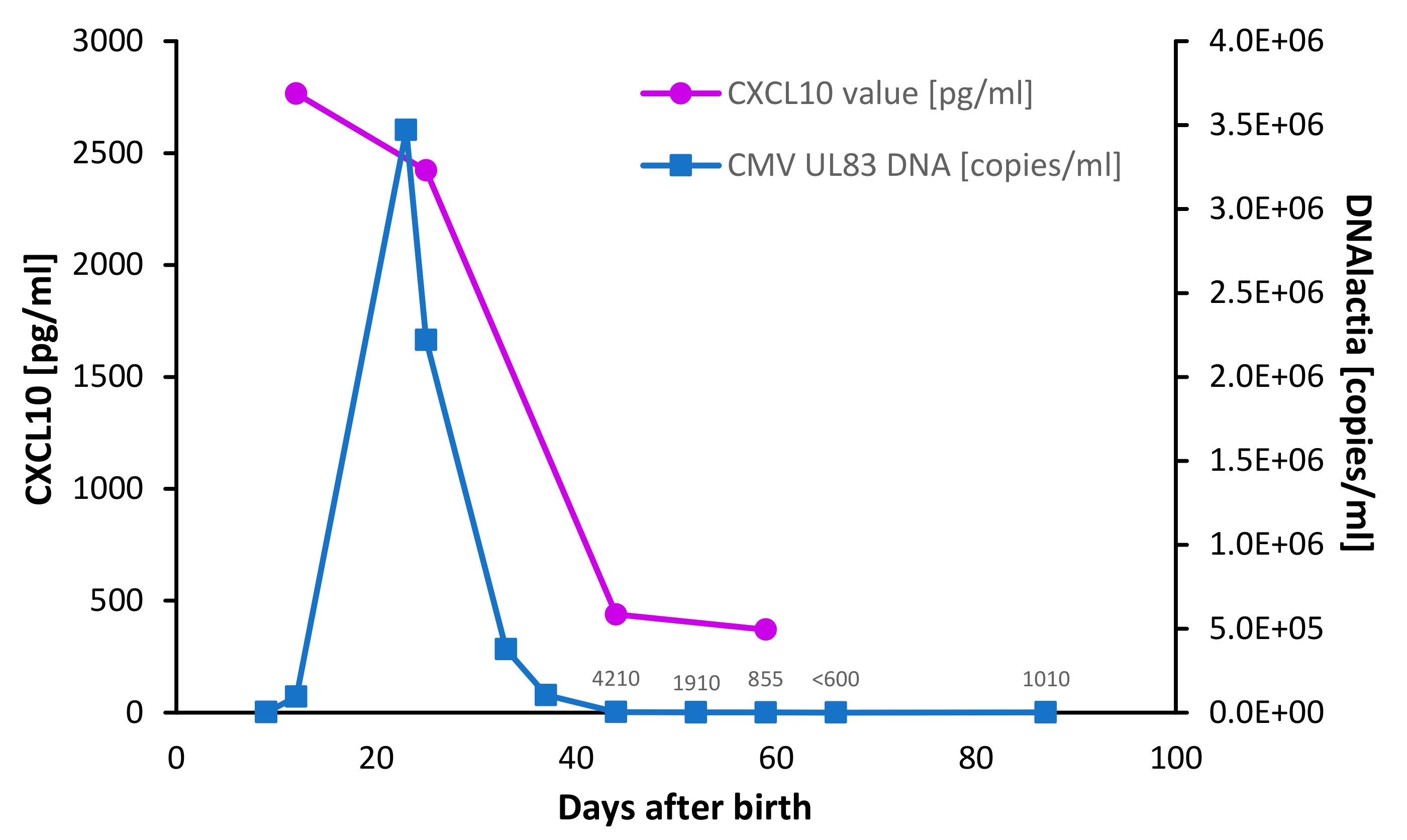
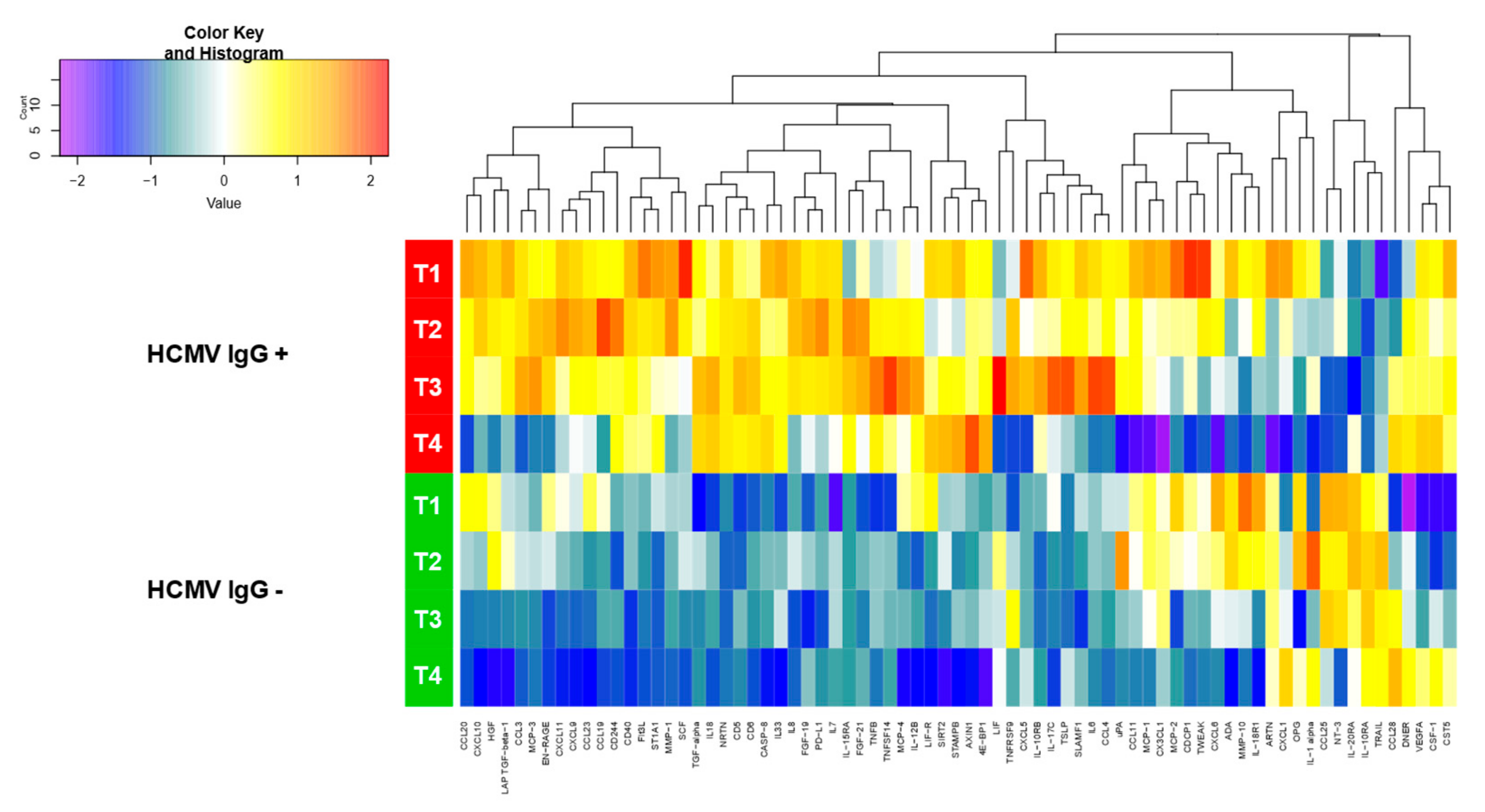
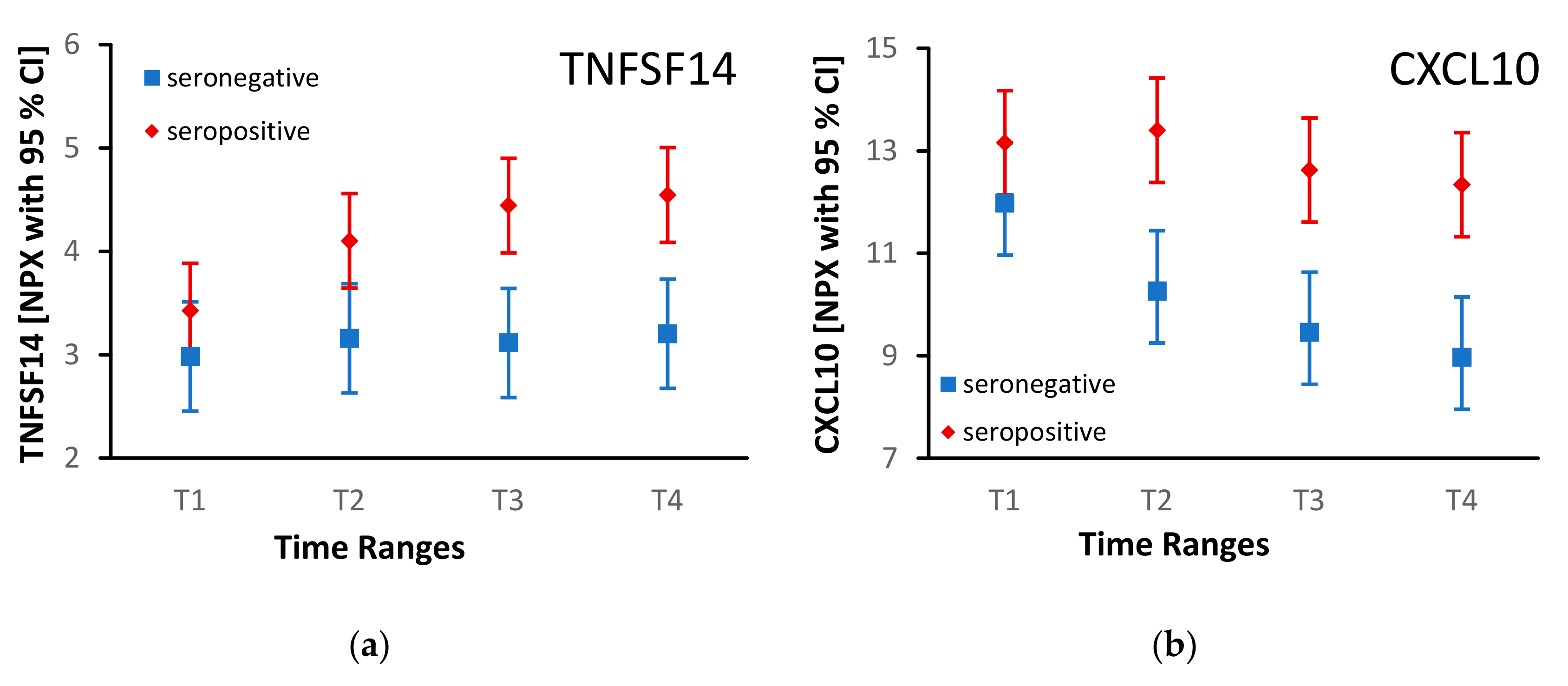
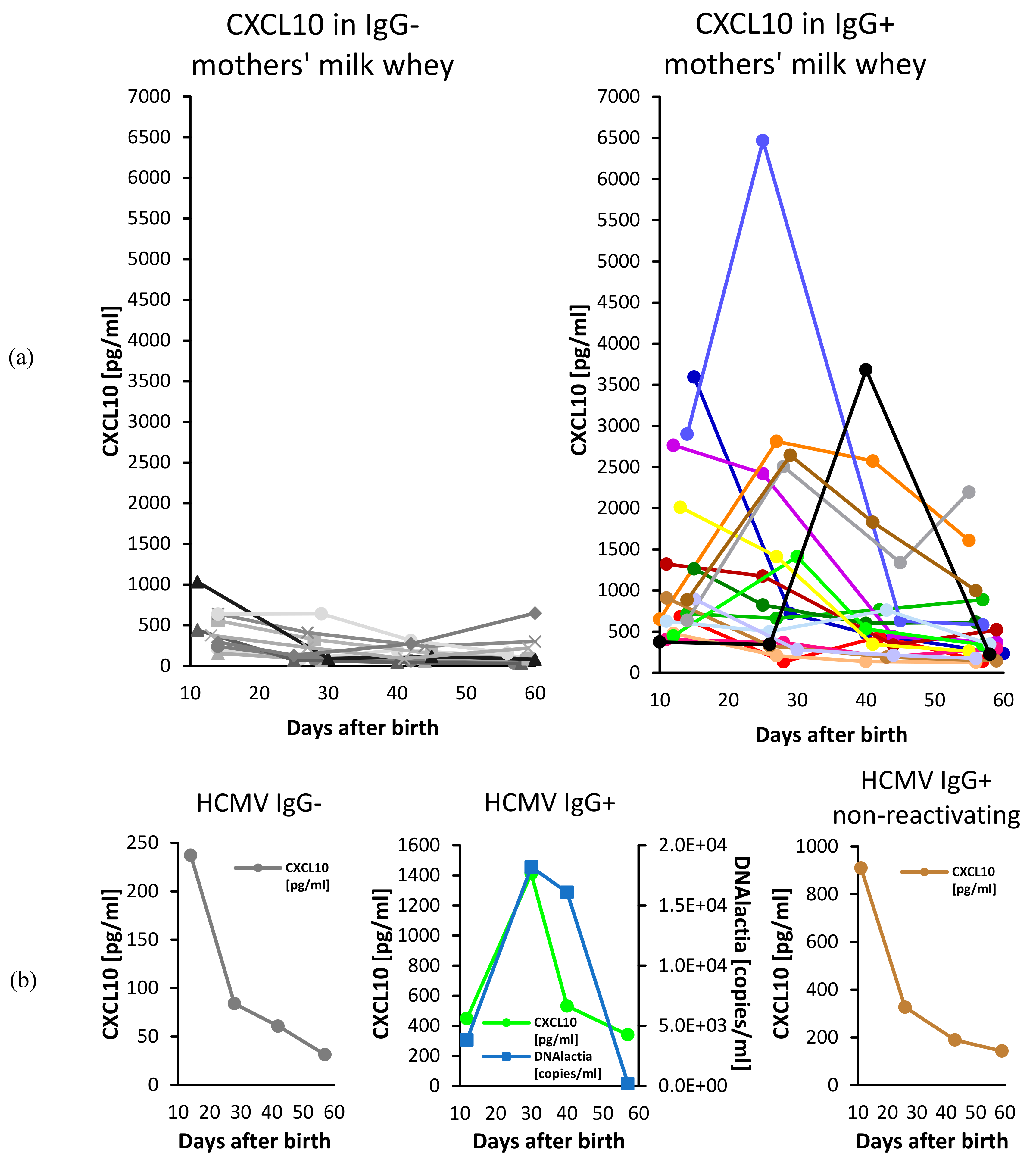
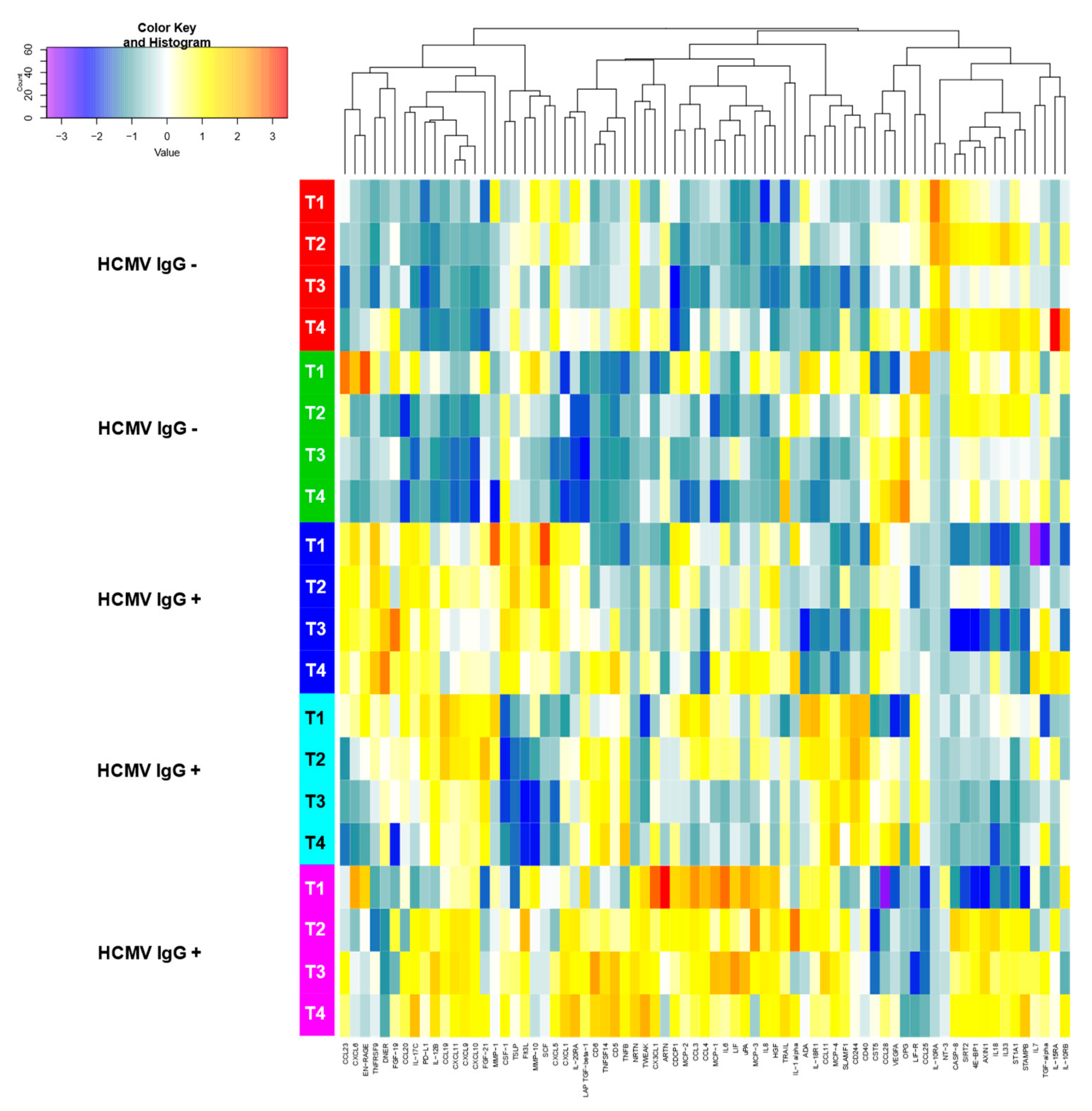
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Hamprecht, K.; Maschmann, J.; Vochem, M.; Dietz, K.; Speer, C.P.; Jahn, G. Epidemiology of transmission of cytomegalovirus from mother to preterm infant by breastfeeding. Lancet 2001, 357, 513–518. [Google Scholar] [CrossRef]
- Hamprecht, K.; Witzel, S.; Maschmann, J.; Dietz, K.; Baumeister, A.; Mikeler, E.; Goelz, R.; Speer, C.P.; Jahn, G. Rapid detection and quantification of cell free cytomegalovirus by a high-speed centrifugation-based microculture assay: Comparison to longitudinally analyzed viral DNA load and pp67 late transcript during lactation. J. Clin. Virol. 2003, 28, 303–316. [Google Scholar] [CrossRef]
- Hamprecht, K.; Goelz, R. Postnatal cytomegalovirus infection through human milk in preterm infants: Transmission, clinical presentation, and prevention. Clin. Perinatol. 2017, 44, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Maschmann, J.; Hamprecht, K.; Dietz, K.; Jahn, G.; Speer, C.P. Cytomegalovirus infection of extremely low—Birth weight infants via breast milk. Clin. Infect. Dis. 2001, 33, 1998–2003. [Google Scholar] [CrossRef]
- Garofalo, R. Cytokines in human milk. J. Pediatr. 2010, 156, S36–S40. [Google Scholar] [CrossRef]
- Srivastava, M.D.; Srivastava, A.; Brouhard, B.; Saneto, R.; Groh-Wargo, S.; Kubit, J. Cytokines in human milk. Res. Commun. Mol. Pathol. Pharm. 1996, 93, 263–287. [Google Scholar]
- Yilmaz, H.L.; Saygili-Yilmaz, E.S.; Gunesacar, R. Interleukin-10 and -12 in human milk at 3 stages of lactation: A longitudinal study. Adv. Ther. 2007, 24, 603–610. [Google Scholar] [CrossRef]
- Erbağcı, A.B.; Çekmen, M.B.; Balat, Ö.; Balat, A.; Aksoy, F.; Tarakçıoğlu, M. Persistency of high proinflammatory cytokine levels from colostrum to mature milk in preeclampsia. Clin. Biochem. 2005, 38, 712–716. [Google Scholar] [CrossRef]
- Goldman, A.S.; Chheda, S.; Garofalo, R.; Schmalstieg, F.C. Cytokines in human milk: Properties and potential effects upon the mammary gland and the neonate. J. Mammary Gland Biol. Neoplasia 1996, 1, 251–258. [Google Scholar] [CrossRef]
- Böttcher, M.F.; Jenmalm, M.C.; Garofalo, R.P.; Björkstén, B. Cytokines in breast milk from allergic and nonallergic mothers. Pediatr. Res. 2000, 47, 157. [Google Scholar] [CrossRef]
- Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E. Maternal weight and excessive weight gain during pregnancy modify the immunomodulatory potential of breast milk. Pediatr. Res. 2012, 72, 77. [Google Scholar] [CrossRef] [PubMed]
- Burch, J.; Karmaus, W.; Gangur, V.; Soto-Ramírez, N.; Yousefi, M.; Goetzl, L.M. Pre-and perinatal characteristics and breast milk immune markers. Pediatr. Res. 2013, 74, 615. [Google Scholar] [CrossRef] [PubMed]
- Reeves, M.B.; Compton, T. Inhibition of inflammatory interleukin-6 activity via extracellular signal-regulated kinase–mitogen-activated protein kinase signaling antagonizes human cytomegalovirus reactivation from dendritic cells. J. Virol. 2011, 85, 12750–12758. [Google Scholar] [CrossRef] [PubMed]
- Kline, J.N.; Hunninghake, G.M.; He, B.; Monick, M.M.; Hunninghake, G.W. Synergistic activation of the human cytomegalovirus major immediate early promoter by prostaglandin E2 and cytokines. Exp. Lung Res. 1998, 24, 3–14. [Google Scholar] [CrossRef]
- Stein, J.; Volk, H.-D.; Liebenthal, C.; Krüger, D.H.; Prösch, S. Tumour necrosis factor α stimulates the activity of the human cytomegalovirus major immediate early enhancer/promoter in immature monocytic cells. J. Gen. Virol. 1993, 74, 2333–2338. [Google Scholar] [CrossRef]
- Lazar, K.; Rabe, T.; Goelz, R.; Hamprecht, K. Human cytomegalovirus reactivation during lactation: Impact of antibody kinetics and neutralization in blood and breast milk. Nutrients 2020, 12, 338. [Google Scholar] [CrossRef]
- Assarsson, E.; Lundberg, M.; Holmquist, G.; Björkesten, J.; Bucht Thorsen, S.; Ekman, D.; Eriksson, A.; Rennel Dickens, E.; Ohlsson, S.; Edfeldt, G.; et al. Homogenous 96-Plex PEA immunoassay exhibiting high sensitivity, specificity, and excellent scalability. PLoS ONE 2014, 9, e95192. [Google Scholar] [CrossRef]
- Baggiolini, M.; Dewald, B.; Moser, B. Human chemokines: An update. Annu. Rev. Immunol. 1997, 15, 675–705. [Google Scholar] [CrossRef]
- Hassiotou, F.; Hepworth, A.R.; Metzger, P.; Tat Lai, C.; Trengove, N.; Hartmann, P.E.; Filgueira, L. Maternal and infant infections stimulate a rapid leukocyte response in breastmilk. Clin. Transl. Immunol. 2013, 2, e3. [Google Scholar] [CrossRef]
- Witkowska-Zimny, M.; Kaminska-El-Hassan, E. Cells of human breast milk. Cell. Mol. Biol. Lett. 2017, 22, 11. [Google Scholar] [CrossRef]
- Takahata, Y.; Takada, H.; Nomura, A.; Nakayama, H.; Ohshima, K.; Hara, T. Detection of interferon-γ-inducible chemokines in human milk. Acta Paediatr. 2003, 92, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Moylan, D.C.; Pati, S.K.; Ross, S.A.; Fowler, K.B.; Boppana, S.B.; Sabbaj, S. Breast milk human cytomegalovirus (CMV) viral load and the establishment of breast milk CMV-pp65-specific CD8 T cells in human CMV infected mothers. J. Infect. Dis. 2017, 216, 1176–1179. [Google Scholar] [CrossRef] [PubMed]
- Maschmann, J.; Goelz, R.; Witzel, S.; Strittmatter, U.; Steinmassl, M.; Jahn, G.; Hamprecht, K. Characterization of human breast milk leukocytes and their potential role in cytomegalovirus transmission to newborns. Neonatology 2015, 107, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Whitbeck, J.C.; Peng, C.; Lou, H.; Xu, R.; Willis, S.H.; De Leon, M.P.; Peng, T.; Nicola, A.V.; Montgomery, R.I.; Warner, M.S. Glycoprotein D of herpes simplex virus (HSV) binds directly to HVEM, a member of the tumor necrosis factor receptor superfamily and a mediator of HSV entry. J. Virol. 1997, 71, 6083–6093. [Google Scholar] [CrossRef]
- Cheung, T.C.; Humphreys, I.R.; Potter, K.G.; Norris, P.S.; Shumway, H.M.; Tran, B.R.; Patterson, G.; Jean-Jacques, R.; Yoon, M.; Spear, P.G. Evolutionarily divergent herpesviruses modulate T cell activation by targeting the herpesvirus entry mediator cosignaling pathway. Proc. Natl. Acad. Sci. USA 2005, 102, 13218–13223. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.M.; Nelson, C.A.; Šedý, J.R. Balancing co-stimulation and inhibition with BTLA and HVEM. Nat. Rev. Immunol. 2006, 6, 671–681. [Google Scholar] [CrossRef]
- Weseslindtner, L.; Nachbagauer, R.; Kundi, M.; Jaksch, P.; Kerschner, H.; Simon, B.; Hatos-Agyi, L.; Scheed, A.; Aberle, J.; Klepetko, W. Human cytomegalovirus infection in lung transplant recipients triggers a CXCL-10 response. Am. J. Transpl. 2011, 11, 542–552. [Google Scholar] [CrossRef]
- Evers, D.L.; Wang, X.; Huang, E.-S. Cellular stress and signal transduction responses to human cytomegalovirus infection. Microbes Infect. 2004, 6, 1084–1093. [Google Scholar] [CrossRef]
- DeMeritt, I.B.; Milford, L.E.; Yurochko, A.D. Activation of the NF-κB pathway in human cytomegalovirus-infected cells is necessary for efficient transactivation of the major immediate-early promoter. J. Virol. 2004, 78, 4498–4507. [Google Scholar] [CrossRef]
- Pahl, H.L. Activators and target genes of Rel/NF-κB transcription factors. Oncogene 1999, 18, 6853. [Google Scholar] [CrossRef]
- Luganini, A.; Terlizzi, M.E.; Gribaudo, G. Bioactive molecules released from cells infected with the human cytomegalovirus. Front. Microbiol. 2016, 7, 715. [Google Scholar] [CrossRef]
- Hamprecht, K.; Maschmann, J.; Müller, D.; Dietz, K.; Besenthal, I.; Goelz, R.; Middeldorp, J.M.; Speer, C.P.; Jahn, G. Cytomegalovirus (CMV) inactivation in breast milk: Reassessment of pasteurization and freeze-thawing. Pediatr. Res. 2004, 56, 529. [Google Scholar] [CrossRef] [PubMed]
- Bapistella, S.; Hamprecht, K.; Thomas, W.; Speer, C.P.; Dietz, K.; Maschmann, J.; Poets, C.F.; Goelz, R. Short-term pasteurization of breast milk to prevent postnatal cytomegalovirus transmission in very preterm infants. Clin. Infect. Dis. 2019, 69, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Maschmann, J.; Müller, D.; Lazar, K.; Goelz, R.; Hamprecht, K. New short-term heat inactivation method of cytomegalovirus (CMV) in breast milk: Impact on CMV inactivation, CMV antibodies and enzyme activities. Arch. Dis. Child. Fetal Neonatal Ed. 2019, 104, F604–F608. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Karmaus, W.; Davis, S.; Gangur, V. Immune markers in breast milk and fetal and maternal body fluids: A systematic review of perinatal concentrations. J. Hum. Lact. 2011, 27, 171–186. [Google Scholar] [CrossRef] [PubMed]
- Ustundag, B.; Yilmaz, E.; Dogan, Y.; Akarsu, S.; Canatan, H.; Halifeoglu, I.; Cikim, G.; Aygun, A.D. Levels of cytokines (IL-1β, IL-2, IL-6, IL-8, TNF-α) and trace elements (Zn, Cu) in breast milk from mothers of preterm and term infants. Mediat. Inflamm. 2005, 2005, 331–336. [Google Scholar] [CrossRef]
- Dupont, L.; Reeves, M.B. Cytomegalovirus latency and reactivation: Recent insights into an age old problem. Rev. Med. Virol. 2016, 26, 75–89. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mother | Age | HCMV Serostatus | Gestational Age at Birth [Weeks + Days] | Onset of HCMV DNAlactia 1 [Day after Birth] | Peak Viral Load [Copies/ml] (Time Range 2)3 |
|---|---|---|---|---|---|
| 1 | 30 | positive | 30 + 2 | 5 | 183 000 (T2) |
| 2 | 28 | positive | 33 + 2 | 5.5 | 11 000 (T2) |
| 3 | 33 | positive | 24 + 2 | 4.5 | 3 470 000 (T2) |
| 4 | 27 | positive | 30 + 6 | 1.5 | 238 000 (T2) |
| 5 | 27 | negative | 26 + 3 | - | - |
| 6 | 31 | negative | 26 + 2 | - | - |
| 7 | 33 | negative | 24 + 5 | - | - |
| Category | Cytokines, Chemokines | Mean NPX Values with 95 % CI (IgG-Seropositive) | Mean NPX Values with 95 % CI (IgG-Seronegative) | P-Values 1 |
|---|---|---|---|---|
| CC-chemokines | CCL19 | 7.6 [6.9; 8.3] | 4.8 [4.0; 5.7] | 6.28E-04 |
| CCL20 | 11.9 [11.5; 12.4] | 10.7 [10.2; 11.2] | 3.13E-03 | |
| MCP-2 | 10.6 [10.2; 11.1] | 9.5 [9.0; 10.0] | 5.59E-03 | |
| CXC-chemokines | CXCL9 | 9.6 [8.6; 10.5] | 5.9 [4.7; 7.0] | 7.51E-04 |
| CXCL11 | 10.7 [9.8; 11.5] | 7.6 [6.6; 8.6] | 7.61E-04 | |
| IL-8 | 9.6 [9.1; 10.2] | 7.9 [7.3; 8.5] | 1.47E-03 | |
| CXCL10 | 12.9 [11.9; 13.8] | 10.2 [9.1; 11.3] | 3.19E-03 | |
| Others | IL-17C | 4.5 [4.1; 4.8] | 3.4 [3.0; 3.8] | 2.37E-03 |
| CD5 | 1.9 [1.5; 2.3] | 0.8 [0.3; 1.3] | 3.97E-03 | |
| TNFSF14 | 4.1 [3.7; 4.6] | 3.1 [2.6; 3.6] | 8.86E-03 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabe, T.; Lazar, K.; Cambronero, C.; Goelz, R.; Hamprecht, K. Human Cytomegalovirus (HCMV) Reactivation in the Mammary Gland Induces a Proinflammatory Cytokine Shift in Breast Milk. Microorganisms 2020, 8, 289. https://doi.org/10.3390/microorganisms8020289
Rabe T, Lazar K, Cambronero C, Goelz R, Hamprecht K. Human Cytomegalovirus (HCMV) Reactivation in the Mammary Gland Induces a Proinflammatory Cytokine Shift in Breast Milk. Microorganisms. 2020; 8(2):289. https://doi.org/10.3390/microorganisms8020289
Chicago/Turabian StyleRabe, Tabea, Katrin Lazar, Christoffer Cambronero, Rangmar Goelz, and Klaus Hamprecht. 2020. "Human Cytomegalovirus (HCMV) Reactivation in the Mammary Gland Induces a Proinflammatory Cytokine Shift in Breast Milk" Microorganisms 8, no. 2: 289. https://doi.org/10.3390/microorganisms8020289
APA StyleRabe, T., Lazar, K., Cambronero, C., Goelz, R., & Hamprecht, K. (2020). Human Cytomegalovirus (HCMV) Reactivation in the Mammary Gland Induces a Proinflammatory Cytokine Shift in Breast Milk. Microorganisms, 8(2), 289. https://doi.org/10.3390/microorganisms8020289