Microbial Degradation of Different Hydrocarbon Fuels with Mycoremediation of Volatiles



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. General Setup
2.2. Soil Analyses
2.3. Experimental Setup for Fungal Growth on Volatiles
2.4. Respiration Measurement
2.5. Modeling
2.6. GC-MS Analysis of Hydrocarbons in Soil
2.7. Identification of Fungal Species
2.8. Statistical Analysis
3. Results and Discussion
3.1. Soil Characteristics
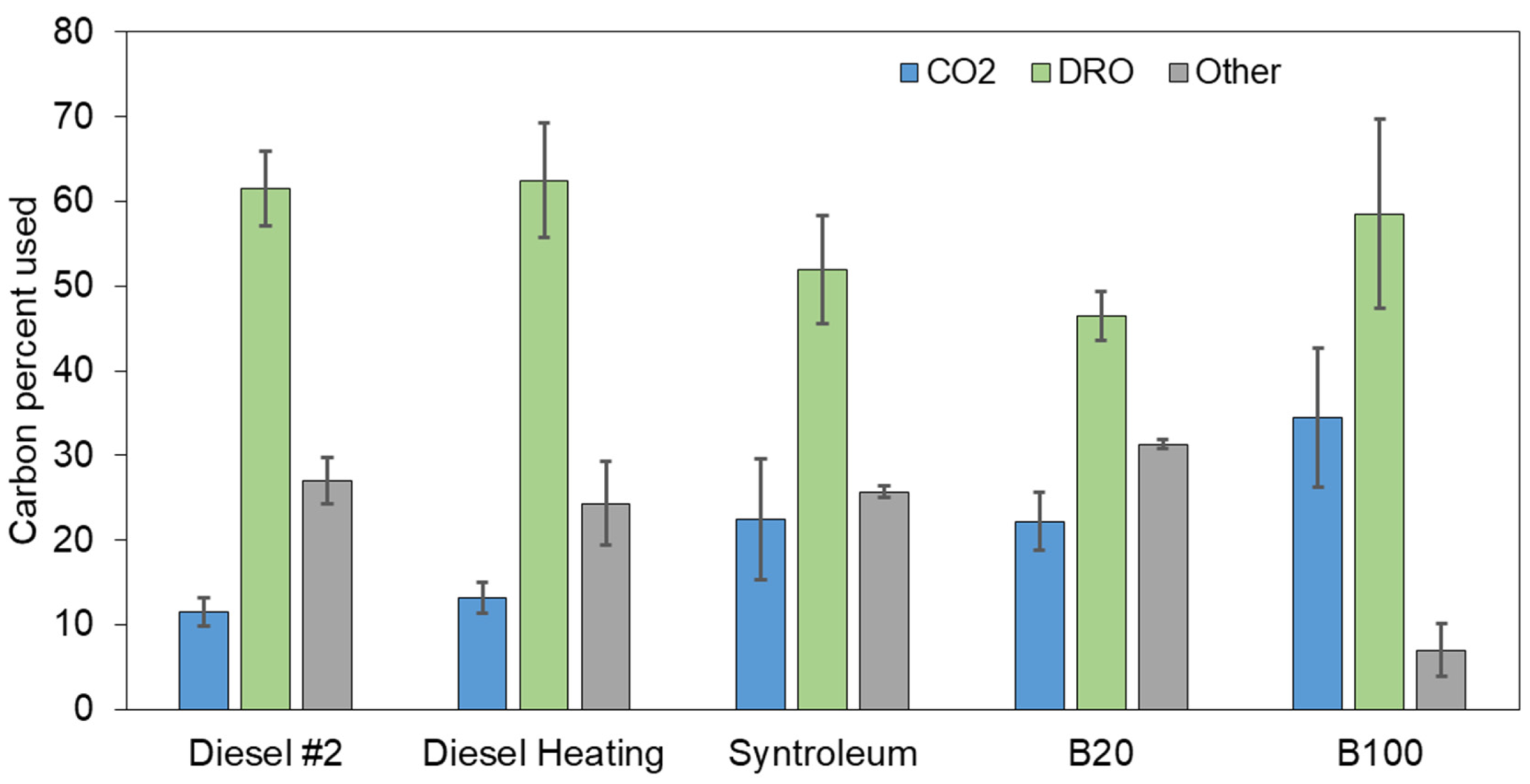
3.2. Hydrocarbon Degradation Rate Constants and Mass Balance for Different Fuels
3.3. Nutrient Availability and Utilization
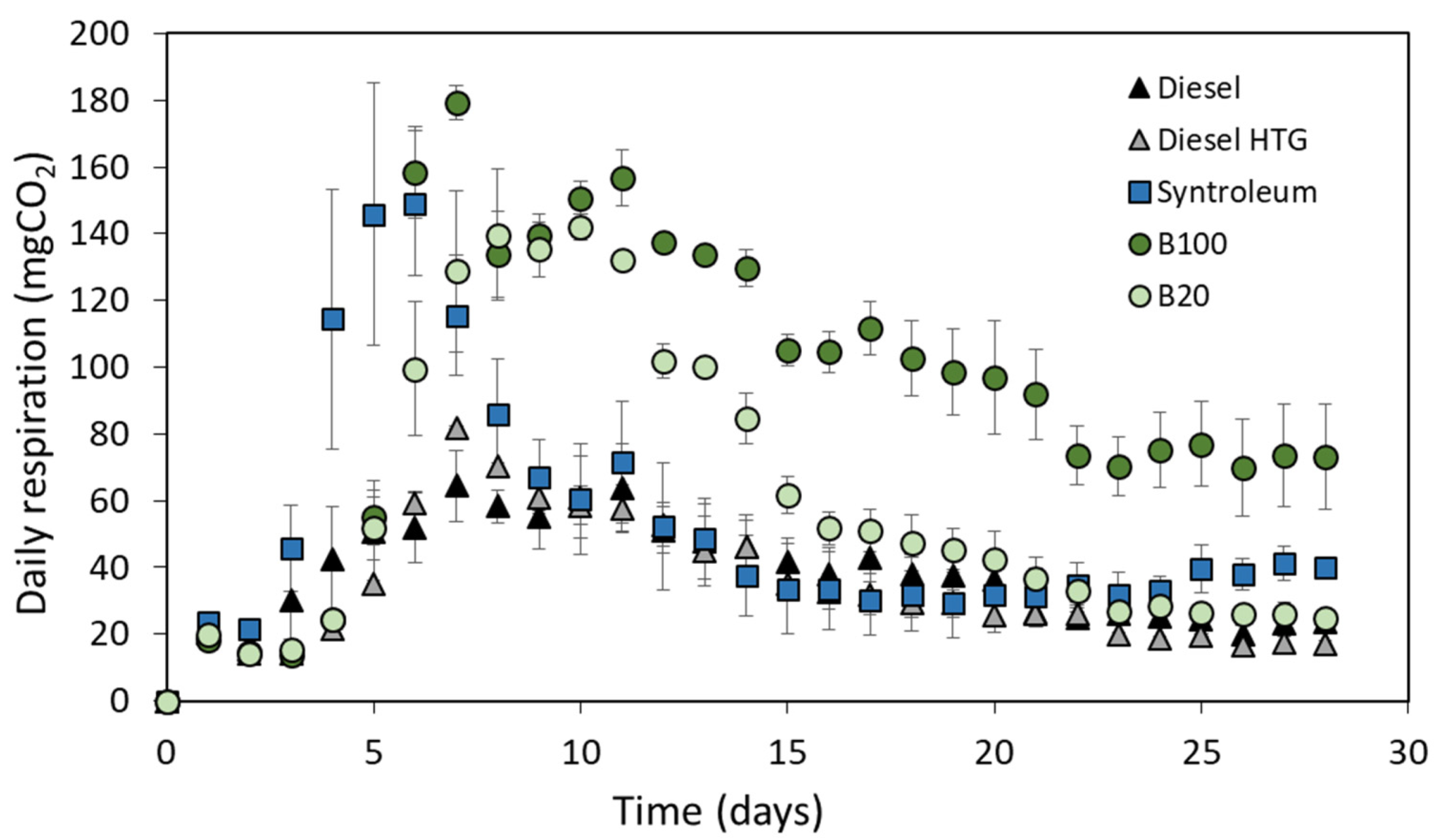
3.4. Degradation of Volatile Compounds by Fungal Communities
3.5. Identification of Fungal Species
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marwood, T.M.; Knoke, K.; Yau, K.; Lee, H.; Trevors, J.T.; Suchorski-Tremblay, A.; Flemming, C.A.; Hodge, V.; Liu, D.L.; Seech, A.G. Comparison of toxicity detected by five bioassays during bioremediation of diesel fuel-spiked soils. Environ. Toxicol. Water Qual. 1998, 13, 117–126. [Google Scholar] [CrossRef]
- Horel, A.; Schiewer, S. Investigation of the physical and chemical parameters affecting biodegradation of diesel and synthetic diesel fuel contaminating Alaskan soils. Cold Reg. Sci. Technol. 2009, 58, 113–119. [Google Scholar] [CrossRef]
- Horel, A.; Schiewer, S. Influence of constant and fluctuating temperature on biodegradation rates of fish biodiesel blends contaminating Alaskan sand. Chemosphere 2011, 83, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Rizzo, C.; Malavenda, R.; Gerçe, B.; Papale, M.; Syldatk, C.; Hausmann, R.; Bruni, V.; Michaud, L.; Lo Giudice, A.; Amalfitano, S. Effects of a simulated acute oil spillage on bacterial communities from Arctic and Antarctic marine sediments. Microorganisms 2019, 7, 632. [Google Scholar] [CrossRef]
- Fritsche, W.; Hofrichter, M. Aerobic degradation by microorganisms. In Environmental Processes-Soil Decontamination; Klein, J., Ed.; Wiley-VCH: Weinheim, Germany, 2000; pp. 146–155. [Google Scholar]
- Atlas, R.M.; Bartha, R. Microbial Ecology: Fundamentals and Applications, 3rd ed.; Benjamin/Cummings Pub. Co.: Redwood City, CA, USA, 1993; 563p. [Google Scholar]
- Huesemann, M.H. Incomplete hydrocarbon biodegradation in contaminated soils: Limitations in bioavailability or inherent recalcitrance? Biorem. J. 1997, 1, 27–39. [Google Scholar] [CrossRef]
- Gruiz, K.; Kriston, E. In situ bioremediation of hydrocarbon in soil. J. Soil Contam. 1995, 4, 163–173. [Google Scholar] [CrossRef]
- Maddela, N.R.; Burgos, R.; Kadiyala, V.; Carrion, A.R.; Bangeppagari, M. Removal of petroleum hydrocarbons from crude oil in solid and slurry phase by mixed soil microorganisms isolated from Ecuadorian oil fields. Int. Biodeterior. Biodegrad. 2016, 108, 85–90. [Google Scholar] [CrossRef]
- Harms, H.; Schlosser, D.; Wick, L.Y. Untapped potential: Exploiting fungi in bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 2011, 9, 177–192. [Google Scholar] [CrossRef]
- Dickson, U.J.; Coffey, M.; Mortimer, R.J.G.; Di Bonito, M.; Ray, N. Mycoremediation of petroleum contaminated soils: Progress, prospects and perspectives. Environ. Sci. Process. Impacts 2019, 21, 1446–1458. [Google Scholar] [CrossRef]
- Prenafeta-Boldú, F.X.; de Hoog, G.S.; Summerbell, R.C. Fungal Communities in Hydrocarbon Degradation. In Microbial Communities Utilizing Hydrocarbons and Lipids: Members, Metagenomics and Ecophysiology; McGenity, T.J., Ed.; Springer International Publishing: Cham, Switzerland, 2018; pp. 1–36. [Google Scholar]
- Wu, Y.-R.; Luo, Z.-H.; Kwok-Kei Chow, R.; Vrijmoed, L.L.P. Purification and characterization of an extracellular laccase from the anthracene-degrading fungus Fusarium solani MAS2. Bioresour. Technol. 2010, 101, 9772–9777. [Google Scholar] [CrossRef]
- Schiewer, S.; Horel, A. Biodiesel addition influences biodegradation rates of fresh and artificially weathered diesel fuel in Alaskan sand. J. Cold Reg. Eng. 2017, 31, 04017012. [Google Scholar] [CrossRef]
- Federal Transit Administration. Demonstration of Fischer-Tropsch Diesel Fuel in Cold Climates; USDOT: Washington, DC, USA, 2007. Available online: http://www.fta.dot.gov/documents/Fischer_Tropsch_Synthetic_Diesel_Demonstration_Project.pdf (accessed on 17 April 2008).
- Steigers, J.A.; Seshadri, G.; Crimp, P.M. Demonstrating the use of Alaska fish oil as a feedstock for the commercial production of biodiesel. In World Renewable Energy Congress VIII; Elsevier Ltd.: Denver, Colorado, USA, 2004; pp. 1–5. [Google Scholar]
- Alaska Energy Authority. Development and Demonstration of Mobile Fish Oil Processing Module. 2007. Available online: ftp://www.aidea.org/REFund/Round%202/Applications/267_Mobile%20Biodiesel%20Processing%20Plant_CCTH/Grant%20Packet/Legal%20issues%20and%20Previous%20Grants%20and%20Trends/Appendix%20L-%20AEA08-013FishOilGrantRFA.pdf.pdf (accessed on 17 May 2017).
- Horel, A.; Schiewer, S. Influence of inocula with prior hydrocarbon exposure on biodegradation rates of diesel, synthetic diesel, and fish-biodiesel in soil. Chemosphere 2014, 109, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Niemeyer, T.K. Soil Heating and Nutrient Supply for the Improvement of Bioremediation Performance in Cold Climates; University of Alaska Fairbanks: Fairbanks, AK, USA, 2003. [Google Scholar]
- Anderson, J. Soil respiration. In Methods of Soil Analysis; Page, A.L., Miller, R.H., Keeney, D.R., Eds.; American Society of Agronomy: Madison, WI, USA, 1982; pp. 143–156. [Google Scholar]
- Horel, A.; Schiewer, S. Impact of VOC removal by activated carbon on biodegradation rates of diesel, Syntroleum and biodiesel in contaminated sand. Sci. Total Environ. 2016, 573, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Gardes, M.; Bruns, T.D. Its primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics; Academic Press, Inc.: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Page, R.D.M. TREEVIEW: An application to display phylogenetic trees on personal computers. Comput. Appl. Biosci. 1996, 12, 357–358. [Google Scholar]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Saadoun, I.M.K.; Al–Ghzawi, Z.D. Bioremediation of Petroleum Contamination; Science Publishers Inc: Enfield, NH, USA, 2005; pp. 173–212. [Google Scholar]
- Bell, T.; Yergeau, E.; Maynard, C.; Juck, D.; Whyte, L.; Greer, C. Predictable bacterial composition and hydrocarbon degradation in Arctic soils following diesel and nutrient disturbance. ISME J. 2013, 7, 1200–1210. [Google Scholar] [CrossRef]
- Bajpai, R.; Kim, J.; Qasim, M. Bioremediation. In Encyclopedia of Agricultural, Food, and Biological Engineering; Heldman, D.R., Ed.; CRC Press: Boca Raton, FL, USA, 2003; pp. 108–118. [Google Scholar]
- Leahy, J.G.; Colwell, R.R. Microbial-degradation of hydrocarbons in the environment. Microbiol. Rev. 1990, 54, 305–315. [Google Scholar] [CrossRef]
- Selker, J.S.; Keller, C.K.; McCord, J.T. Vadose biogeochemical processes. In Vadose Zone Processes; Lewis Publisher: Washington, DC, USA, 1999; pp. 147–227. [Google Scholar]
- Bergin, S. Annual Report for the Ultra-Clean Fischer-Tropsch Fuels Production and Demonstration Project; Integrated Concepts and Research Corporation: Sterling Heights, MI, USA, 2004; Available online: http://www.fischer-tropsch.org/DOE/DOE_reports/41099/41099-05/41099-05.pdf (accessed on 11 July 2006).
- Lotter, S.; Heerenklage, J.; Stegmann, R. Carbon balance and modelling of oil degradation in soil bioreactors. In Treatment of Contaminated Soil: Fundamentals, Analysis, Applications; Stegmann, R., Brunner, G., Calmano, W., Matz, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2001; pp. 355–363. [Google Scholar]
- Prince, R.C.; Haitmanek, C.; Lee, C.C. The primary aerobic biodegradation of biodiesel B20. Chemosphere 2008, 71, 1446–1451. [Google Scholar] [CrossRef]
- Sendzikiene, E.; Makareviciene, V.; Janulis, P.; Makareviciute, D. Biodegradability of biodiesel fuel of animal and vegetable origin. Eur. J. Lipid Sci. Technol. 2007, 109, 493–497. [Google Scholar] [CrossRef]
- Lisiecki, P.; Chrzanowski, L.; Szulc, A.; Lawniczak, L.; Bialas, W.; Dziadas, M.; Owsianiak, M.; Staniewski, J.; Cyplik, P.; Marecik, R.; et al. Biodegradation of diesel/biodiesel blends in saturated sand microcosms. Fuel 2014, 116, 321–327. [Google Scholar] [CrossRef]
- Ferguson, S.H.; Franzmann, P.D.; Revill, A.T.; Snape, I.; Rayner, J.L. The effects of nitrogen and water on mineralisation of hydrocarbons in diesel-contaminated terrestrial Antarctic. Cold Reg. Sci. Technol. 2003, 37, 197–212. [Google Scholar] [CrossRef]
- Walworth, J.L.; Woolard, C.R.; Harris, K.C. Nutrient amendments for contaminated peri-glacial soils: Use of cod bone meal as a controlled release nutrient source. Cold Reg. Sci. Technol. 2003, 37, 81–88. [Google Scholar] [CrossRef]
- Sakamoto, K.; Oba, Y. Effect of fungal to bacterial biomass ratio on the relationship between CO2 evolution and total soil microbial biomass. Biol. Fertil. Soils 1994, 17, 39–44. [Google Scholar] [CrossRef]
- Morgan, P.; Watkinson, R.J. Hydrocarbon degradation in soils and methods for soil biotreatment. Crit. Rev. Biotechnol. 1989, 8, 305–333. [Google Scholar] [CrossRef]
- Margesin, R.; Schinner, F. Laboratory bioremediation experiments with soil from a diesel-oil contaminated site—Significant role of cold-adapted microorganisms and fertilizers. J. Chem. Technol. Biotechnol. 1997, 70, 92–98. [Google Scholar] [CrossRef]
- Brook, T.R.; Stiver, W.H.; Zytner, R.G. Biodegradation of diesel fuel in soil under various nitrogen addition regimes. Soil Sediment Contam. 2001, 10, 539–553. [Google Scholar] [CrossRef]
- Walworth, J.; Pond, A.; Snape, I.; Rayner, J.; Ferguson, S.; Harvey, P. Nitrogen requirements for maximizing petroleum bioremediation in a sub-Antarctic soil. Cold Reg. Sci. Technol. 2007, 48, 84–91. [Google Scholar] [CrossRef]
- Prenafeta-Boldú, F.X.; Kuhn, A.; Luykx, D.M.A.M.; Anke, H.; van Groenestijn, J.W.; de Bont, J.A.M. Isolation and characterisation of fungi growing on volatile aromatic hydrocarbons as their sole carbon and energy source. Mycological Research 2001, 105, 477–484. [Google Scholar] [CrossRef]
- Nikolova, N.; Nenov, V. BTEX degradation by fungi. Water Sci. Technol. 2005, 51, 87–93. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cox, H.H.J.; Moerman, R.E.; van Baalen, S.; van Heiningen, W.N.M.; Doddema, H.J.; Harder, W. Performance of a styrene-degrading biofilter containing the yeast Exophiala jeanselmei. Biotechnol. Bioeng. 1997, 53, 259–266. [Google Scholar] [CrossRef]
- Mineki, S.; Suzuki, K.; Iwata, K.; Nakajima, D.; Goto, S. Degradation of Polyaromatic Hydrocarbons by Fungi Isolated from Soil in Japan. Polycycl. Aromat. Compd. 2015, 35, 120–128. [Google Scholar] [CrossRef]
- Weber, F.J.; Hage, K.C.; de Bont, J.A. Growth of the fungus Cladosporium sphaerospermum with toluene as the sole carbon and energy source. Appl. Environ. Microbiol. 1995, 61, 3562–3566. [Google Scholar] [CrossRef] [PubMed]
- Singh, H. Mycorrhizal fungi in rhizosphere remediation. In Mycoremediation: Fungal Bioremediation; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 533–572. [Google Scholar]
- Bajaj, S.; Singh, D.K. Biodegradation of persistent organic pollutants in soil, water and pristine sites by cold-adapted microorganisms: Mini review. Int. Biodeterior. Biodegrad. 2015, 100, 98–105. [Google Scholar] [CrossRef]
- Al-Nasrawi, H. Biodegradation of crude oil by fungi isolated from Gulf of Mexico. J. Bioremediation Biodegrad. 2012, 3, 1–6. [Google Scholar]
- Wang, S.; Nomura, N.; Nakajima, T.; Uchiyama, H. Case study of the relationship between fungi and bacteria associated with high-molecular-weight polycyclic aromatic hydrocarbon degradation. J. Biosci. Bioeng. 2012, 113, 624–630. [Google Scholar] [CrossRef]
- Varjani, S.J. Microbial degradation of petroleum hydrocarbons. Bioresour. Technol. 2017, 223, 277–286. [Google Scholar] [CrossRef]
- April, T.M.; Foght, J.M.; Currah, R.S. Hydrocarbon-degrading filamentous fungi isolated from flare pit soils in northern and western Canada. Can. J. Microbiol. 2000, 46, 38–49. [Google Scholar] [CrossRef]
- Gestel, K.V.; Mergaert, J.; Swings, J.; Coosemans, J.; Ryckeboer, J. Bioremediation of diesel-contaminated soil by composting with biowaste. Environ. Pollut. 2003, 125, 361–368. [Google Scholar] [CrossRef]
- Husaini, A.; Roslan, H.A.; Hii, K.S.Y.; Ang, C.H. Biodegradation of aliphatic hydrocarbon by indigenous fungi isolated from used motor oil contaminated sites. World J. Microbiol. Biotechnol. 2008, 24, 2789–2797. [Google Scholar] [CrossRef]
- Mancera-Lopez, M.; Esparza-Garcia, F.; Chavez-Gomez, B.; Rodriguez-Vazquez, R.; Saucedo-Castaneda, G.; Barrera-Cortes, J. Bioremediation of an aged hydrocarbon-contaminated soil by a combined system of biostimulation-bioaugmentation with filamentous fungi. Int. Biodeterior. Biodegrad. 2008, 61, 151–160. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horel, A.; Schiewer, S. Microbial Degradation of Different Hydrocarbon Fuels with Mycoremediation of Volatiles. Microorganisms 2020, 8, 163. https://doi.org/10.3390/microorganisms8020163
Horel A, Schiewer S. Microbial Degradation of Different Hydrocarbon Fuels with Mycoremediation of Volatiles. Microorganisms. 2020; 8(2):163. https://doi.org/10.3390/microorganisms8020163
Chicago/Turabian StyleHorel, Agota, and Silke Schiewer. 2020. "Microbial Degradation of Different Hydrocarbon Fuels with Mycoremediation of Volatiles" Microorganisms 8, no. 2: 163. https://doi.org/10.3390/microorganisms8020163
APA StyleHorel, A., & Schiewer, S. (2020). Microbial Degradation of Different Hydrocarbon Fuels with Mycoremediation of Volatiles. Microorganisms, 8(2), 163. https://doi.org/10.3390/microorganisms8020163