Targeted Genome Mining—From Compound Discovery to Biosynthetic Pathway Elucidation

Abstract
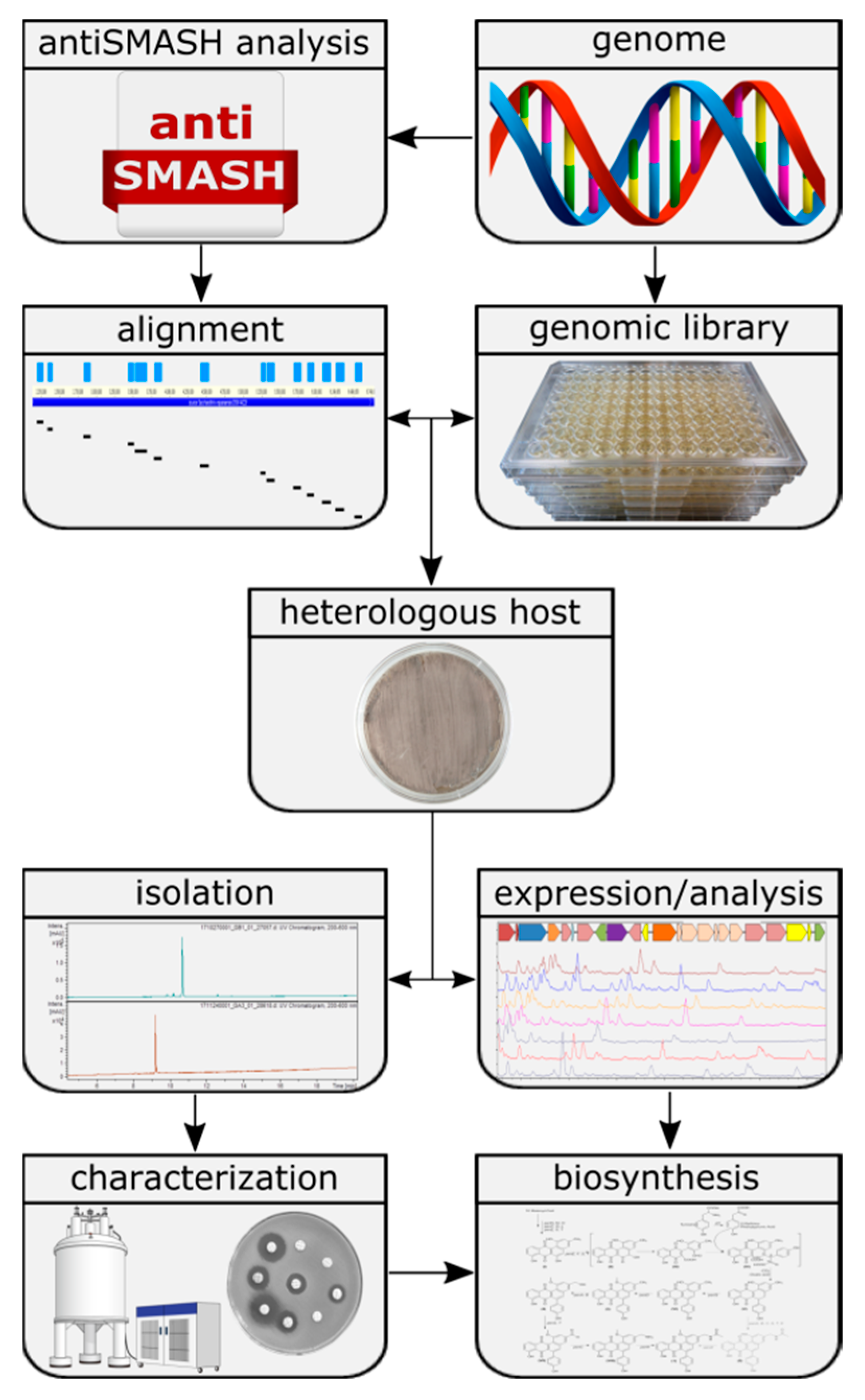
1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. In-Silico Analysis of Gene Clusters
2.3. Extraction, Dereplication and Isolation
2.4. Modification of BACs
2.5. Construction of Plasmids
Synthesis of 1E5_CMP1 and 1E5_CMP2
3. Results and Discussion
3.1. Selection of Putative Biosynthetic Gene Clusters
3.2. Success Rate of the Expression System
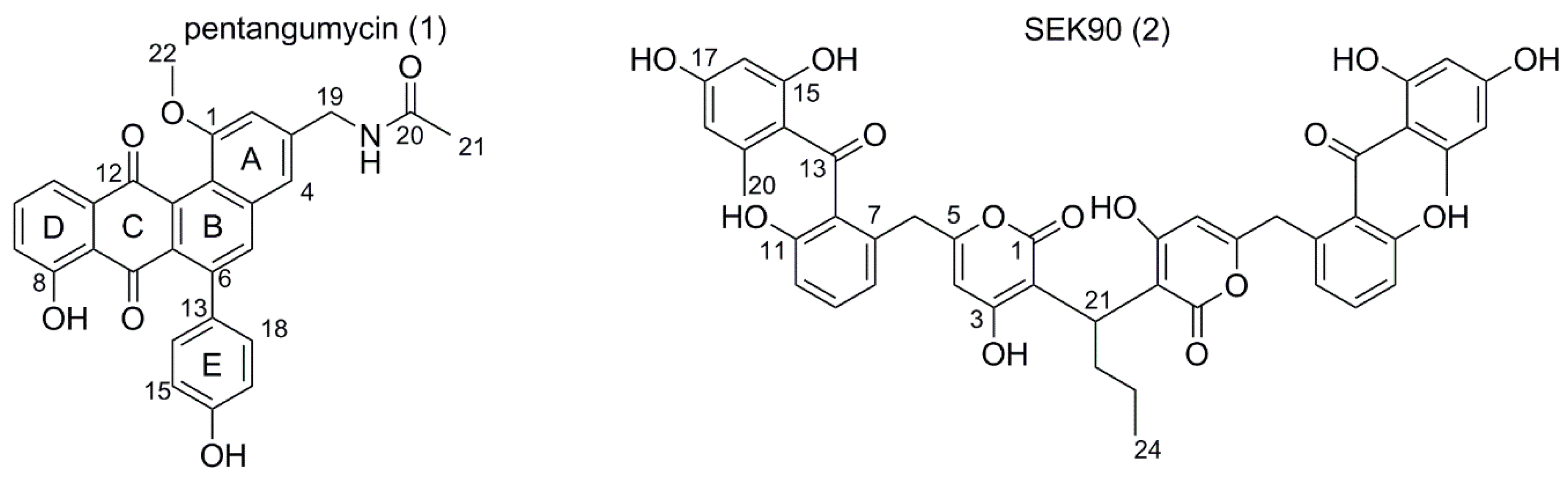
3.3. Isolation of Pentangumycin and SEK90
3.4. Both 1 and 2 Are the Result of Type II PKS Gene Cluster Expression
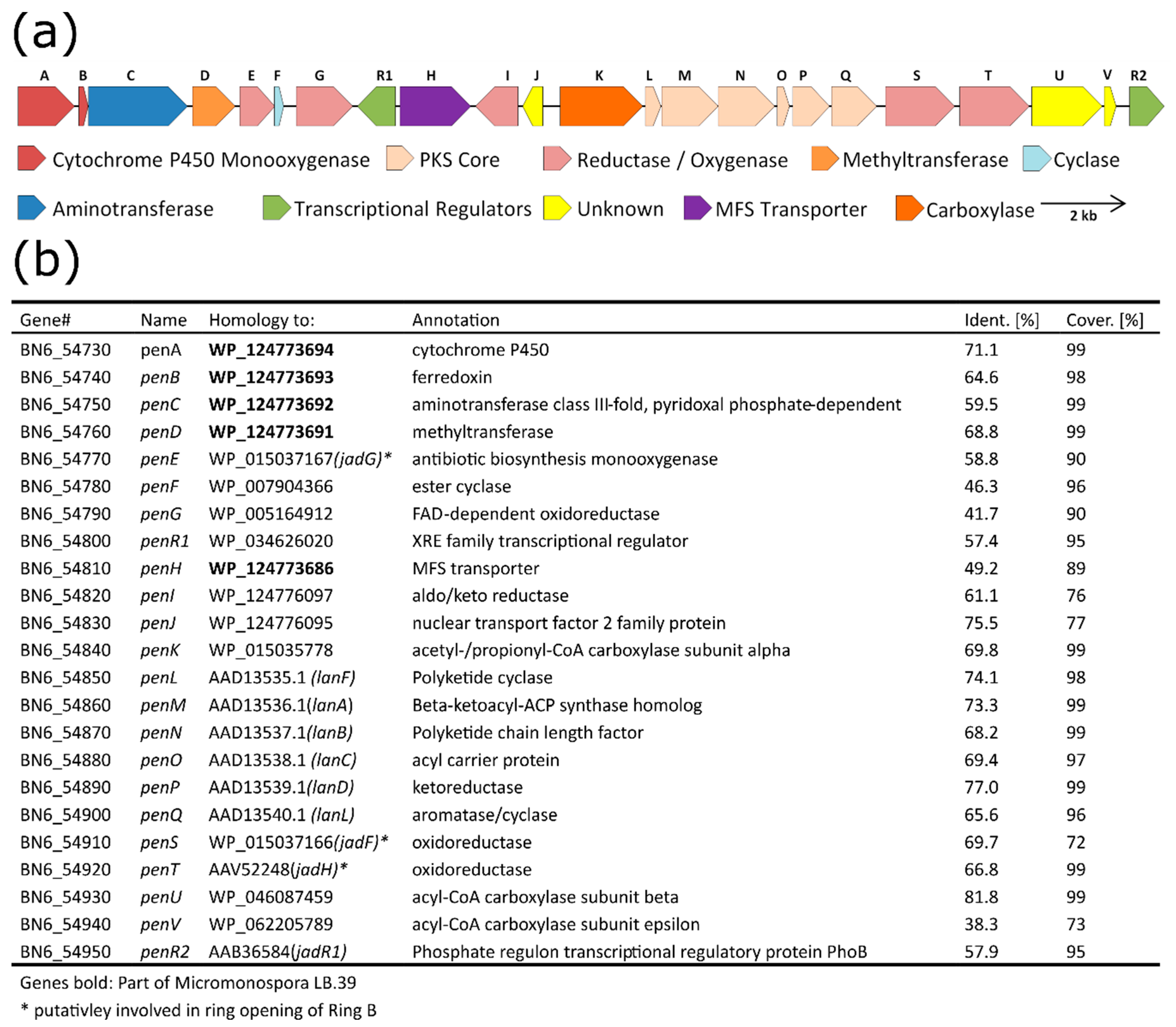
3.5. Detailed Analysis and Border Prediction of the Biosynthetic Gene Cluster of Pentangumycin
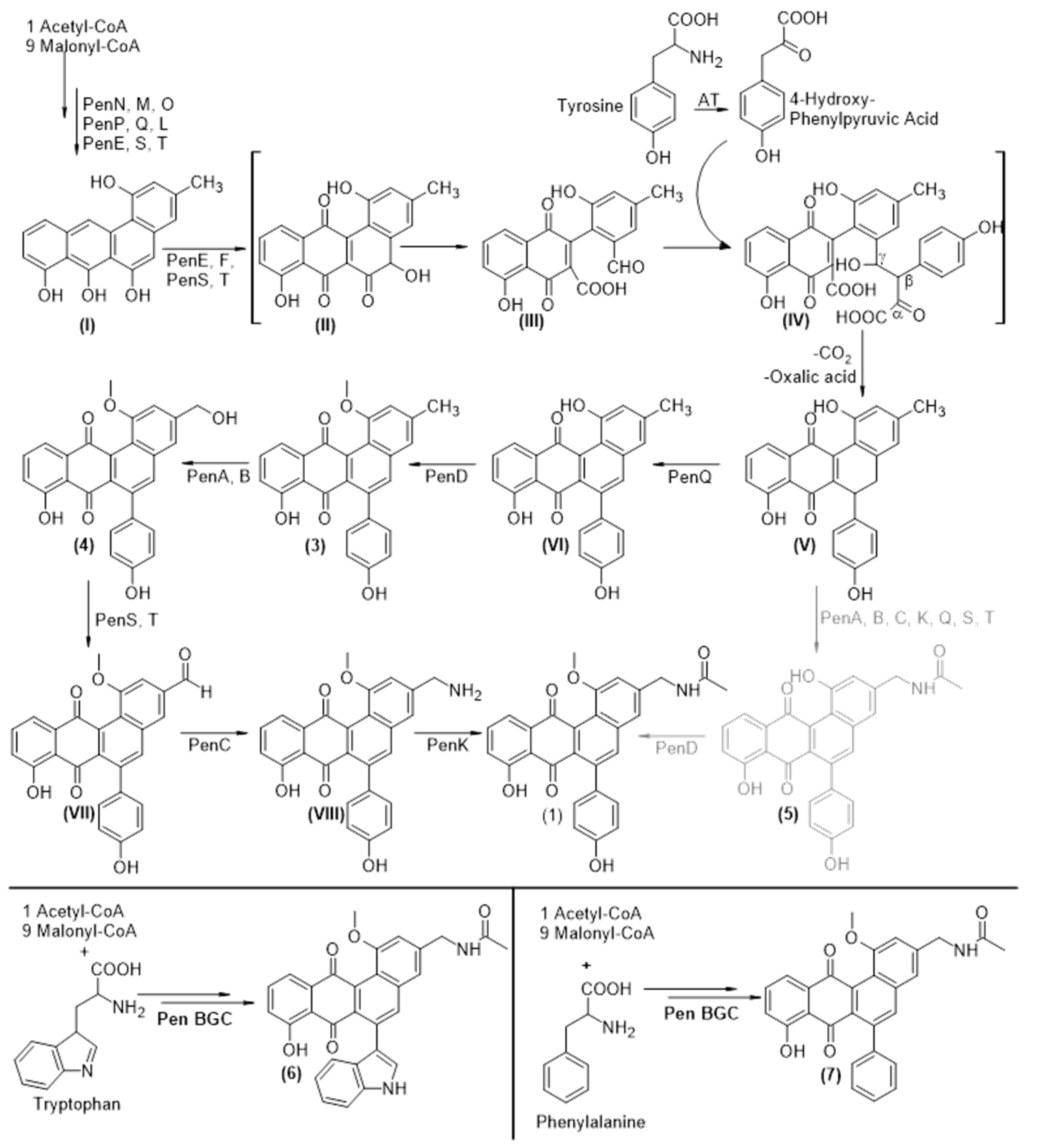
3.6. Biosynthesis of Pentangumycins Core Structure
3.7. Tailoring Steps of Pentangumycin’s Biosynthesis
3.8. Analysis of Regulatory Gene Functions
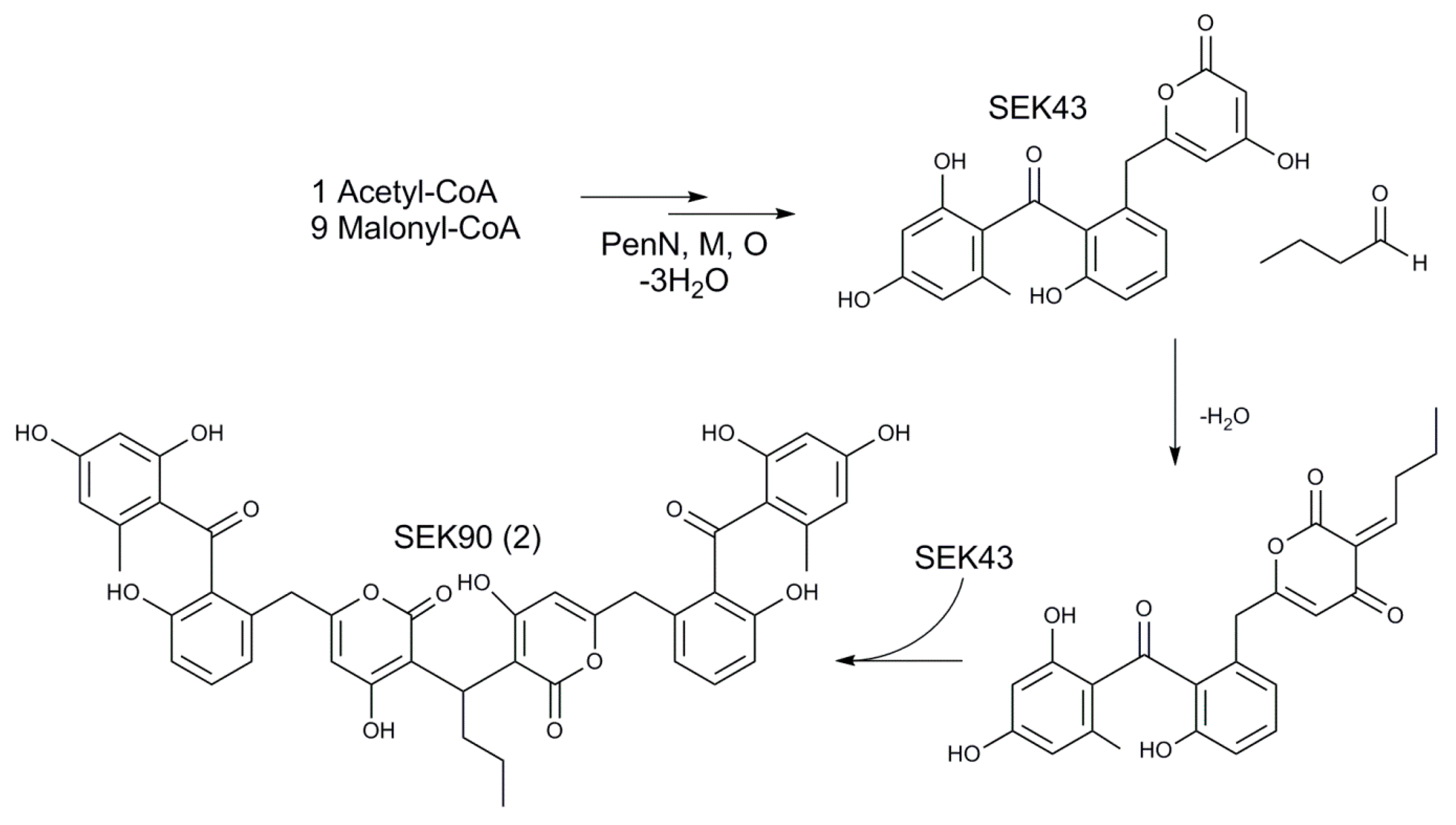
3.9. Origin of SEK90
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Berdy, J. Bioactive microbial metabolites. J. Antibiot. (Tokyo) 2005, 58, 1–26. [Google Scholar] [CrossRef]
- Walsh, C.T. A chemocentric view of the natural product inventory. Nat. Chem. Biol. 2015, 11, 620–624. [Google Scholar] [CrossRef]
- Kong, D.X.; Guo, M.Y.; Xiao, Z.H.; Chen, L.L.; Zhang, H.Y. Historical variation of structural novelty in a natural product library. Chem. Biodivers 2011, 8, 1968–1977. [Google Scholar] [CrossRef]
- Cech, N.B.; Yu, K. Mass spectrometry for natural products research: Challenges, pitfalls, and opportunities. LCGC N. Am. 2013, 31, 938–947. [Google Scholar]
- Zhang, Y.; Pan, G.; Zou, Z.; Fan, K.; Yang, K.; Tan, H. Jadr*-mediated feed-forward regulation of cofactor supply in jadomycin biosynthesis. Mol. Microbiol. 2013, 90, 884–897. [Google Scholar] [CrossRef] [PubMed]
- National-Human-Genome-Research-Institute; Wetterstrand, K.A. DNA Sequencing Costs: Data from the Nhgri Genome Sequencing Program (gsp). 2019. Available online: www.genome.gov/sequencingcostsdata (accessed on 16 April 2019).
- Ahmed, N. A flood of microbial genomes-do we need more? PLoS ONE 2009, 4, e5831. [Google Scholar] [CrossRef] [PubMed]
- Doroghazi, J.R.; Albright, J.C.; Goering, A.W.; Ju, K.S.; Haines, R.R.; Tchalukov, K.A.; Labeda, D.P.; Kelleher, N.L.; Metcalf, W.W. A roadmap for natural product discovery based on large-scale genomics and metabolomics. Nat. Chem. Biol. 2014, 10, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Tokovenko, B.; Rebets, Y.; Luzhetskyy, A. Automating assessment of the undiscovered biosynthetic potential of actinobacteria. bioRxiv 2016, 036087. [Google Scholar] [CrossRef]
- Weber, T.; Blin, K.; Duddela, S.; Krug, D.; Kim, H.U.; Bruccoleri, R.; Lee, S.Y.; Fischbach, M.A.; Muller, R.; Wohlleben, W.; et al. Antismash 3.0-a comprehensive resource for the genome mining of biosynthetic gene clusters. Nucleic Acids Res. 2015, 43, W237–W243. [Google Scholar] [CrossRef]
- Myronovskyi, M.; Rosenkranzer, B.; Nadmid, S.; Pujic, P.; Normand, P.; Luzhetskyy, A. Generation of a cluster-free streptomyces albus chassis strains for improved heterologous expression of secondary metabolite clusters. Metab. Eng. 2018, 49, 316–324. [Google Scholar] [CrossRef]
- Gomez-Escribano, J.P.; Bibb, M.J. Engineering streptomyces coelicolor for heterologous expression of secondary metabolite gene clusters. Microb. Biotechnol. 2011, 4, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Mazodier, P.; Petter, R.; Thompson, C. Intergeneric conjugation between escherichia coli and streptomyces species. J. Bacteriol. 1989, 171, 3583–3585. [Google Scholar] [CrossRef] [PubMed]
- Gust, B.; Chandra, G.; Jakimowicz, D.; Yuqing, T.; Bruton, C.J.; Chater, K.F. Lambda red-mediated genetic manipulation of antibiotic-producing streptomyces. Adv. Appl. Microbiol. 2004, 54, 107–128. [Google Scholar] [PubMed]
- Blin, K.; Wolf, T.; Chevrette, M.G.; Lu, X.; Schwalen, C.J.; Kautsar, S.A.; Suarez Duran, H.G.; de Los Santos, E.L.C.; Kim, H.U.; Nave, M.; et al. Antismash 4.0-improvements in chemistry prediction and gene cluster boundary identification. Nucleic Acids Res. 2017, 45, W36–W41. [Google Scholar] [CrossRef] [PubMed]
- Skinnider, M.A.; Merwin, N.J.; Johnston, C.W.; Magarvey, N.A. Prism 3: Expanded prediction of natural product chemical structures from microbial genomes. Nucleic Acids Res. 2017, 45, W49–W54. [Google Scholar] [CrossRef] [PubMed]
- Labeda, D.P.; Lechevalier, M.P. Amendment of the genus saccharothrix labeda et al. 1984 and descriptions of saccharothrix espanaensis sp. Nov., saccharothrix cryophilis sp. Nov., and saccharothrix mutabilis comb. Nov. Int. Syst. Evol. Microbiol. 1989, 39, 420–423. [Google Scholar] [CrossRef]
- Strobel, T.; Al-Dilaimi, A.; Blom, J.; Gessner, A.; Kalinowski, J.; Luzhetska, M.; Puhler, A.; Szczepanowski, R.; Bechthold, A.; Ruckert, C. Complete genome sequence of saccharothrix espanaensis dsm 44229(t) and comparison to the other completely sequenced pseudonocardiaceae. BMC Genom. 2012, 13, 465. [Google Scholar] [CrossRef]
- Singh, M.P.; Petersen, P.J.; Weiss, W.J.; Kong, F.; Greenstein, M. Saccharomicins, novel heptadecaglycoside antibiotics produced by saccharothrix espanaensis: Antibacterial and mechanistic activities. Antimicrob. Agents Chemother. 2000, 44, 2154–2159. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012. [Google Scholar]
- Kieser, T.; Bibb, M.J.; Buttner, M.J.; Chater, K.F.; Hopwood, D.A. Practical Streptomyces Genetics; John Innes Foundation: Norwich, UK, 2010; p. 613. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Myronovskyi, M.; Rosenkranzer, B.; Luzhetskyy, A. Iterative marker excision system. Appl. Microbiol. Biotechnol. 2014, 98, 4557–4570. [Google Scholar] [CrossRef]
- Siegl, T.; Tokovenko, B.; Myronovskyi, M.; Luzhetskyy, A. Design, construction and characterisation of a synthetic promoter library for fine-tuned gene expression in actinomycetes. Metab. Eng. 2013, 19, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Brenner, M. Molekularbiologische Untersuchungen zur Biosynthese der Oligosaccharid-Antibiotika Saccharomicin a und b in Saccharothrix Espanaensis. Ph.D. Thesis, Albert-Ludwig-Universität Freiburg, Freiburg im Breisgau, Germany, 2016. [Google Scholar]
- Berner, M.; Krug, D.; Bihlmaier, C.; Vente, A.; Muller, R.; Bechthold, A. Genes and enzymes involved in caffeic acid biosynthesis in the actinomycete saccharothrix espanaensis. J. Bacteriol. 2006, 188, 2666–2673. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P.; Erehman, J.; Gohlke, B.O.; Wilhelm, T.; Preissner, R.; Dunkel, M. Super natural ii—A database of natural products. Nucleic Acids Res. 2015, 43, D935–D939. [Google Scholar] [CrossRef] [PubMed]
- Klementz, D.; Doring, K.; Lucas, X.; Telukunta, K.K.; Erxleben, A.; Deubel, D.; Erber, A.; Santillana, I.; Thomas, O.S.; Bechthold, A.; et al. Streptomedb 2.0—An extended resource of natural products produced by streptomycetes. Nucleic Acids Res. 2016, 44, D509–D514. [Google Scholar] [CrossRef]
- Pait, I.G.U.; Kitani, S.; Roslan, F.W.; Ulanova, D.; Arai, M.; Ikeda, H.; Nihira, T. Discovery of a new diol-containing polyketide by heterologous expression of a silent biosynthetic gene cluster from streptomyces lavendulae fri-5. J. Ind. Microbiol. Biotechnol. 2018, 45, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Lithwick, G.; Margalit, H. Hierarchy of sequence-dependent features associated with prokaryotic translation. Genome Res. 2003, 13, 2665–2673. [Google Scholar] [CrossRef] [PubMed]
- Gustafsson, C.; Govindarajan, S.; Minshull, J. Codon bias and heterologous protein expression. Trends Biotechnol. 2004, 22, 346–353. [Google Scholar] [CrossRef] [PubMed]
- Kane, J.F. Effects of rare codon clusters on high-level expression of heterologous proteins in escherichia coli. Curr. Opin. Chem. Biol. 1995, 6, 494–500. [Google Scholar] [CrossRef]
- Liu, S.; Wang, M.; Du, G.; Chen, J. Improving the active expression of transglutaminase in streptomyces lividans by promoter engineering and codon optimization. BMC Biotechnol. 2016, 16, 75. [Google Scholar] [CrossRef]
- Olano, C.; Garcia, I.; Gonzalez, A.; Rodriguez, M.; Rozas, D.; Rubio, J.; Sanchez-Hidalgo, M.; Brana, A.F.; Mendez, C.; Salas, J.A. Activation and identification of five clusters for secondary metabolites in streptomyces albus j1074. Microb. Biotechnol. 2014, 7, 242–256. [Google Scholar] [CrossRef]
- Freudl, R.; Braun, G.; Hindennach, I.; Henning, U. Lethal mutations in the structural gene of an outer membrane protein (ompa) of escherichia coli k12. Mol. Gen. Genet. 1985, 201, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Qiu, J.; Zhuo, Y.; Zhu, D.; Zhou, X.; Zhang, L.; Bai, L.; Deng, Z. Overexpression of the abc transporter avtab increases avermectin production in streptomyces avermitilis. Appl. Microbiol. Biotechnol. 2011, 92, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Doull, J.L.; Singh, A.K.; Hoare, M.; Ayer, S.W. Conditions for the production of jadomycin b by streptomyces venezuelae isp5230: Effects of heat shock, ethanol treatment and phage infection. J. Ind. Microbiol. Biotechnol. 1994, 13, 120–125. [Google Scholar]
- Pishchany, G.; Mevers, E.; Ndousse-Fetter, S.; Horvath, D.J., Jr.; Paludo, C.R.; Silva-Junior, E.A.; Koren, S.; Skaar, E.P.; Clardy, J.; Kolter, R. Amycomicin is a potent and specific antibiotic discovered with a targeted interaction screen. Proc. Natl. Acad. Sci. USA 2018, 115, 10124–10129. [Google Scholar] [CrossRef] [PubMed]
- Moon, K.; Xu, F.; Zhang, C.; Seyedsayamdost, M.R. Bioactivity-hites unveils cryptic antibiotics encoded in actinomycete bacteria. ACS Chem. Biol. 2019, 14, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Seyedsayamdost, M.R. Toward a global picture of bacterial secondary metabolism. J. Ind. Microbiol. Biotechnol. 2019, 46, 301–311. [Google Scholar] [CrossRef]
- Doull, J.L.; Ayer, S.W.; Singh, A.K.; Thibault, P. Production of a novel polyketide antibiotic, jadomycin b, by streptomyces venezuelae following heat shock. J. Antibiot. (Tokyo) 1993, 46, 869–871. [Google Scholar] [CrossRef]
- Drautz, H.; Zahner, H.; Rohr, J.; Zeeck, A. Metabolic products of microorganisms. 234. Urdamycins, new angucycline antibiotics from streptomyces fradiae. I. Isolation, characterization and biological properties. J. Antibiot. (Tokyo) 1986, 39, 1657–1669. [Google Scholar] [CrossRef]
- Robertson, A.W.; MacLeod, J.M.; MacIntyre, L.W.; Forget, S.M.; Hall, S.R.; Bennett, L.G.; Correa, H.; Kerr, R.G.; Goralski, K.B.; Jakeman, D.L. Post polyketide synthase carbon-carbon bond formation in type-ii pks-derived natural products from streptomyces venezuelae. J. Org. Chem. 2018, 83, 1876–1890. [Google Scholar] [CrossRef]
- Qin, Z.; Munnoch, J.T.; Devine, R.; Holmes, N.A.; Seipke, R.F.; Wilkinson, K.A.; Wilkinson, B.; Hutchings, M.I. Formicamycins, antibacterial polyketides produced by streptomyces formicae isolated from african tetraponera plant-ants. Chem. Sci. 2017, 8, 3218–3227. [Google Scholar] [CrossRef]
- Potterat, O.; Puder, C.; Wagner, K.; Bolek, W.; Vettermann, R.; Kauschke, S.G. Chlorocyclinones a-d, chlorinated angucyclinones from streptomyces sp. Strongly antagonizing rosiglitazone-induced ppar-gamma activation. J. Nat. Prod. 2007, 70, 1934–1938. [Google Scholar] [CrossRef] [PubMed]
- Louden, B.C.; Haarmann, D.; Lynne, A.M. Use of blue agar cas assay for siderophore detection. J. Microbiol. Biol. Educ. 2011, 12, 51–53. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, R.; Ebert-Khosla, S.; Hopwood, D.A.; Khosla, C. Rational design of aromatic polyketide natural products by recombinant assembly of enzymatic subunits. Nature 1995, 375, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Girvan, H.M.; Munro, A.W. Applications of microbial cytochrome p450 enzymes in biotechnology and synthetic biology. Curr. Opin. Chem. Biol. 2016, 31, 136–145. [Google Scholar] [CrossRef]
- Waskell, L.; Kim, J.-J.P. Electron transfer partners of cytochrome p450. In Cytochrome p450: Structure, Mechanism, and Biochemistry; de Ortiz Montellano, P.R., Ed.; Springer: Cham, Switzerland, 2015; pp. 33–68. [Google Scholar]
- Sasaki, M.; Akahira, A.; Oshiman, K.; Tsuchido, T.; Matsumura, Y. Purification of cytochrome p450 and ferredoxin, involved in bisphenol a degradation, from sphingomonas sp. Strain ao1. Appl. Environ. Microbiol. 2005, 71, 8024–8030. [Google Scholar] [CrossRef]
- Chen, Y.H.; Wang, C.C.; Greenwell, L.; Rix, U.; Hoffmeister, D.; Vining, L.C.; Rohr, J.; Yang, K.Q. Functional analyses of oxygenases in jadomycin biosynthesis and identification of jadh as a bifunctional oxygenase/dehydrase. J. Biol. Chem. 2005, 280, 22508–22514. [Google Scholar] [CrossRef]
- Wang, L.; Tian, X.; Wang, J.; Yang, H.; Fan, K.; Xu, G.; Yang, K.; Tan, H. Autoregulation of antibiotic biosynthesis by binding of the end product to an atypical response regulator. Proc. Natl. Acad. Sci. USA 2009, 106, 8617–8622. [Google Scholar] [CrossRef]
- Novakova, R.; Homerova, D.; Feckova, L.; Kormanec, J. Characterization of a regulatory gene essential for the production of the angucycline-like polyketide antibiotic auricin in streptomyces aureofaciens ccm 3239. Microbiology 2005, 151, 2693–2706. [Google Scholar] [CrossRef]
- Ostash, I.; Ostash, B.; Walker, S.; Fedorenko, V. Proton-dependent transporter gene lndj confers resistance to landomycin e in streptomyces globisporus 1912. Russ. J. Genet. 2007, 43, 852–857. [Google Scholar] [CrossRef]
- Rebets, Y.; Dutko, L.; Ostash, B.; Luzhetskyy, A.; Kulachkovskyy, O.; Yamaguchi, T.; Nakamura, T.; Bechthold, A.; Fedorenko, V. Function of lani in regulation of landomycin a biosynthesis in streptomyces cyanogenus s136 and cross-complementation studies with streptomyces antibiotic regulatory proteins encoding genes. Arch. Microbiol. 2008, 189, 111–120. [Google Scholar] [CrossRef]
- Pickens, L.B.; Tang, Y. Oxytetracycline biosynthesis. J. Biol. Chem. 2010, 285, 27509–27515. [Google Scholar] [CrossRef]
- Westrich, L.; Domann, S.; Faust, B.; Bedford, D.; Hopwood, D.A.; Bechthold, A. Cloning and characterization of a gene cluster from streptomyces cyanogenus s136 probably involved in landomycin biosynthesis. FEMS Microbiol. Lett. 1999, 170, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Rix, U.; Zheng, J.; Remsing Rix, L.L.; Greenwell, L.; Yang, K.; Rohr, J. The dynamic structure of jadomycin b and the amino acid incorporation step of its biosynthesis. J. Am. Chem. Soc. 2004, 126, 4496–4497. [Google Scholar] [CrossRef] [PubMed]
- Sharif, E.U.; O’Doherty, G.A. Biosynthesis and total synthesis studies on the jadomycin family of natural products. Eur. J. Org. Chem. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Yushchuk, O.; Kharel, M.; Ostash, I.; Ostash, B. Landomycin biosynthesis and its regulation in streptomyces. Appl. Microbiol. Biotechnol. 2019, 103, 1659–1665. [Google Scholar] [CrossRef]
- Dickens, M.L.; Ye, J.; Strohl, W.R. Analysis of clustered genes encoding both early and late steps in daunomycin biosynthesis by streptomyces sp. Strain c5. J. Bacteriol. 1995, 177, 536–543. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Olano, C.; Moss, S.J.; Brana, A.F.; Sheridan, R.M.; Math, V.; Weston, A.J.; Mendez, C.; Leadlay, P.F.; Wilkinson, B.; Salas, J.A. Biosynthesis of the angiogenesis inhibitor borrelidin by streptomyces parvulus tu4055: Insights into nitrile formation. Mol. Microbiol. 2004, 52, 1745–1756. [Google Scholar] [CrossRef]
- Liu, W.C.; Parker, L.; Slusarchyk, S.; Greenwood, G.L.; Grahm, S.F.; Meyers, E. Isolation, characterization, and structure of rabelomycin, a new antibiotic. J. Antibiot. (Tokyo) 1970, 23, 437–441. [Google Scholar] [CrossRef]
- Herrmann, S.; Siegl, T.; Luzhetska, M.; Petzke, L.; Jilg, C.; Welle, E.; Erb, A.; Leadlay, P.F.; Bechthold, A.; Luzhetskyy, A. Site-specific recombination strategies for engineering actinomycete genomes. Appl. Environ. Microbiol. 2012, 78, 1804–1812. [Google Scholar] [CrossRef]
- Horbal, L.; Marques, F.; Nadmid, S.; Mendes, M.V.; Luzhetskyy, A. Secondary metabolites overproduction through transcriptional gene cluster refactoring. Metab. Eng. 2018, 49, 299–315. [Google Scholar] [CrossRef]
- Meurer, G.; Gerlitz, M.; Wendt-Pienkowski, E.; Vining, L.C.; Rohr, J.; Hutchinson, C.R. Iterative type ii polyketide synthases, cyclases and ketoreductases exhibit context-dependent behavior in the biosynthesis of linear and angular decapolyketides. Chem. Biol. 1997, 4, 433–443. [Google Scholar] [CrossRef][Green Version]
- Zawada, R.J.; Khosla, C. Domain analysis of the molecular recognition features of aromatic polyketide synthase subunits. J. Biol. Chem. 1997, 272, 16184–16188. [Google Scholar] [CrossRef] [PubMed]
- Bao, W.; Sheldon, P.J.; Wendt-Pienkowski, E.; Hutchinson, C.R. The streptomyces peucetius dpsc gene determines the choice of starter unit in biosynthesis of the daunorubicin polyketide. J. Bacteriol. 1999, 181, 4690–4695. [Google Scholar] [CrossRef] [PubMed]
- Nachtigall, J. Strukturaufklärung von Naturstoffen aus Bakterien der Gattung Streptomyces. Ph.D. Thesis, Technischen Universität Berlin, Fakultät II—Mathematik und Naturwissenschaften der Technischen, Berlin, Germany, 2012. [Google Scholar]
- Huang, C.; Yang, C.; Zhu, Y.; Zhang, W.; Yuan, C.; Zhang, C. Marine bacterial aromatic polyketides from host-dependent heterologous expression and fungal mode of cyclization. Front. Chem. 2018, 6, 528. [Google Scholar] [CrossRef] [PubMed]
- de March, P.; Moreno-Mañas, M.; Casado, J.; Pleixats, R.; Roca, J.L.; Trius, A. The reactivity of 4-hydroxy-6-methyl-2-pyrone towards aliphatic saturated and α,β-unsaturated aldehydes. J. Heterocycl. Chem. 1984, 21, 85–89. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pentangumycin (1) | SEK90 (2) | ||||||
|---|---|---|---|---|---|---|---|
| Pos. | δC | δH (J in Hz) | HMBC | Pos. | δC | δH (J in Hz) | HMBC |
| 1 | 156.7 C | - | - | 1 | 166.5 C | - | - |
| 2 | 108.7 CH | 7.10 s | 1, 4, 12b, 19 | 2 | 102.7 C | - | - |
| 3 | 143.2 C | - | - | 3 | 165.5 C | - | - |
| 4 | 117.5 CH | 7.44 s | 1, 2, 4a, 5, 12b, 19 | 4 | 101.9 CH | 5.68 (s br) | 2, 5, 6 |
| 4a | 136.6 C | - | - | 5 | 160.6 C | - | - |
| 5 | 135.1 CH | 7.88 s | 1, 4, 4a, 6a, 7, 12a, 12b, 13 | 6 | 36.1 CH2 | 3.55 (s br) | 4, 5, 7, 8, 12 |
| 6 | 139.1 C | - | - | 7 | 132.5 C | - | - |
| 6a | 131.2 C | - | - | 8 | 120.1 CH | 6.74 dd (7.7, N/A) | 9, 10, 11, 12, 13 |
| 7 | 188.0 C | - | - | 9 | 130.9 CH | 7.2 dd (7.9, N/A) | 7, 8, 10, 11, 12 |
| 7a | 115.8 C | - | - | 10 | 114.4 CH | 6.77 dd (8.1, N/A) | 8, 9, 11, 12, 13 |
| 8 | 160.5 C | - | - | 11 | 153.3 C | - | - |
| 9 | 123.0 CH | 7.26 m | 7a, 11 | 12 | 130.6 C | - | - |
| 10 | 136.7 CH | 7.73 t (7.5) | 8, 11a | 13 | 199.9 C | - | - |
| 11 | 117.2 CH | 7.50 dd (7.5, 0.93) | 7a, 9, 10, 8, 12 | 14 | 115.3 C | - | - |
| 11a | 135.5 C | - | - | 15 | 164.9 C | - | - |
| 12 | 185.5 C | - | - | 16 | 100.5 CH | 6.11 d (2.4) | 14, 15, 17 |
| 12a | 140.0 C | - | - | 17 | 166.3 C | - | - |
| 12b | 119.5 C | - | - | 18 | 111.4 CH | 6.04 dd (0.64, 2.4) | 13, 14, 15, 16, 20 |
| 13 | 132.2 C | - | - | 19 | 142.7 C | - | - |
| 14/18 | 129.9 CH | 7.26 m | 6, 14/18, 16 | 20 | 21.2 CH3 | 1.8 s | 14, 18, 19 |
| 15/17 | 114.8 CH | 6.78 dt (8.5, 2) | 13, 15/17, 16 | 21 | 29.5 CH | 4.36 t (8.2) | 1, 2, 3, 22, 23 |
| 16 | 156.6 C | - | - | 22 | 31.5 CH2 | 1.75 dt (7.3, 8.2) | 2, 21, 23, 24 |
| 19 | 42.4 CH2 | 4.43 d (6) | 2, 3, 4, 20 | 23 | 20.6 CH2 | 1.05 dq (7.3) | 21, 22, 24 |
| 20 | 169.6 C | - | - | 24 | 13.8 CH3 | 0.89 t (7.3) | 22, 23 |
| 21 | 22.8 CH3 | 1.93 s | 20 | - | - | - | - |
| 22 | 56.1 CH3 | 3.88 s | 1 | - | - | - | - |
| NH | - | 8.53 t (br) | 19, 20 | - | - | - | - |
| OH1 | - | - | - | - | - | 12.67 s | 14, 15, 16 |
| OH2 | - | - | - | - | - | 11.49 s (br) | - |
| OH3 | - | - | - | - | - | 10.38 s | 16, 18 |
| OH4 | - | - | - | - | - | 9.78 s | 10, 11, 12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gummerlich, N.; Rebets, Y.; Paulus, C.; Zapp, J.; Luzhetskyy, A. Targeted Genome Mining—From Compound Discovery to Biosynthetic Pathway Elucidation. Microorganisms 2020, 8, 2034. https://doi.org/10.3390/microorganisms8122034
Gummerlich N, Rebets Y, Paulus C, Zapp J, Luzhetskyy A. Targeted Genome Mining—From Compound Discovery to Biosynthetic Pathway Elucidation. Microorganisms. 2020; 8(12):2034. https://doi.org/10.3390/microorganisms8122034
Chicago/Turabian StyleGummerlich, Nils, Yuriy Rebets, Constanze Paulus, Josef Zapp, and Andriy Luzhetskyy. 2020. "Targeted Genome Mining—From Compound Discovery to Biosynthetic Pathway Elucidation" Microorganisms 8, no. 12: 2034. https://doi.org/10.3390/microorganisms8122034
APA StyleGummerlich, N., Rebets, Y., Paulus, C., Zapp, J., & Luzhetskyy, A. (2020). Targeted Genome Mining—From Compound Discovery to Biosynthetic Pathway Elucidation. Microorganisms, 8(12), 2034. https://doi.org/10.3390/microorganisms8122034