Specialisation of Yeast Genera in Different Phases of Bee Bread Maturation


Abstract
1. Introduction
2. Materials and Methods
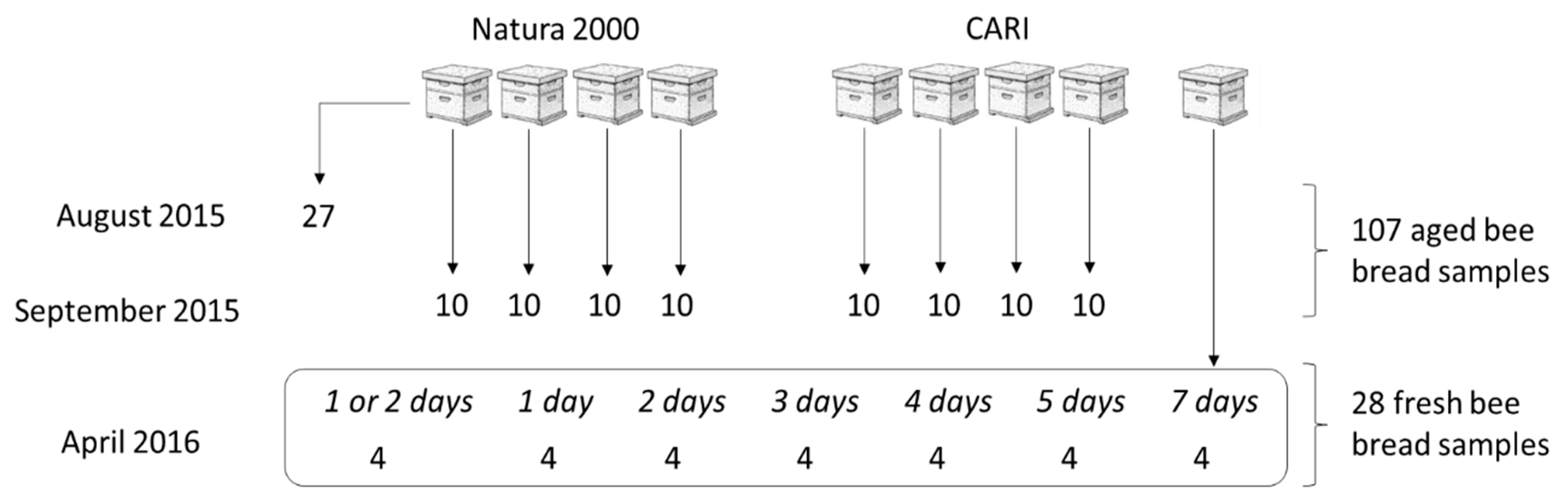
2.1. Samples
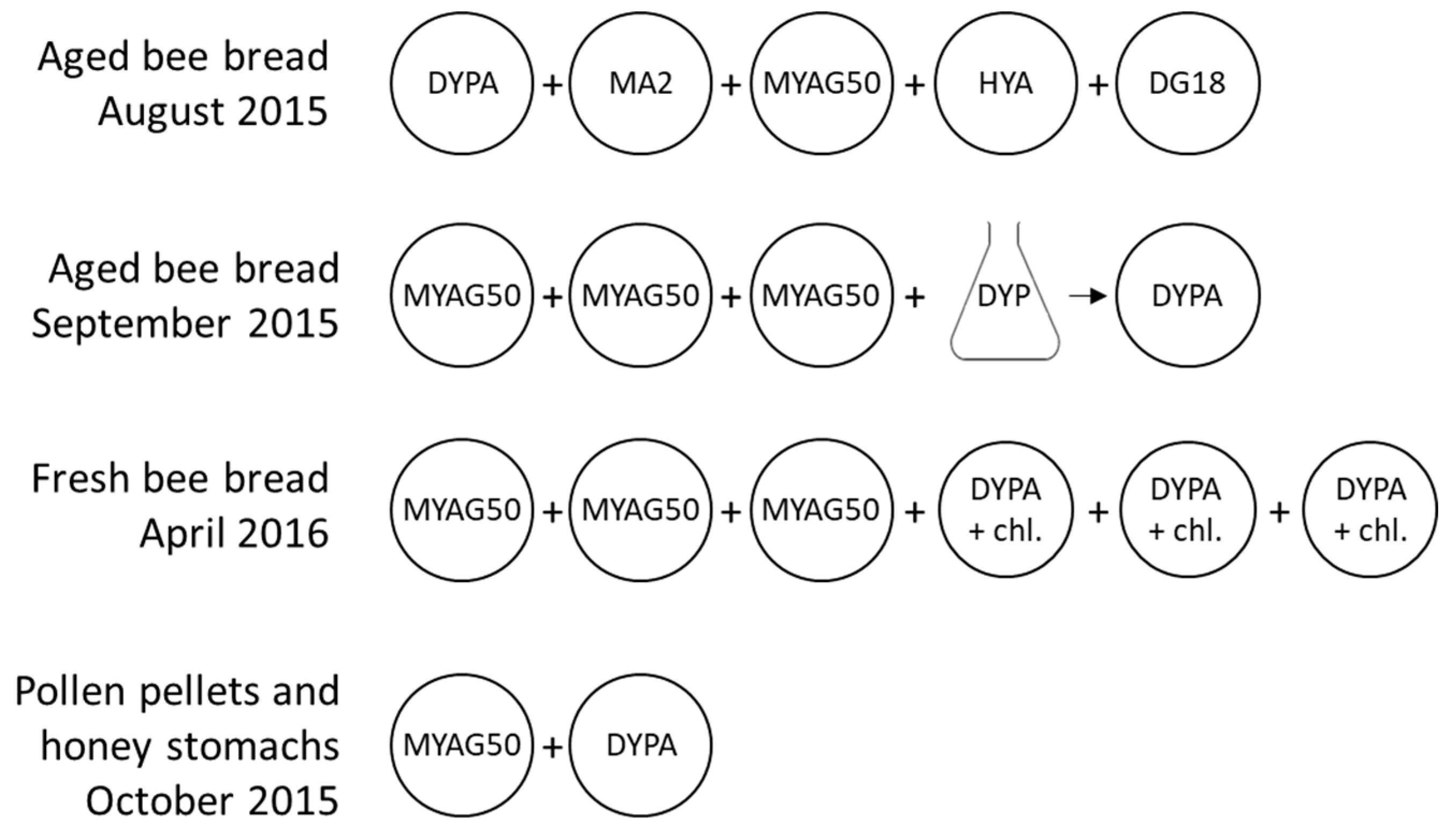
2.2. Cultures
2.3. Isolates and Identifications
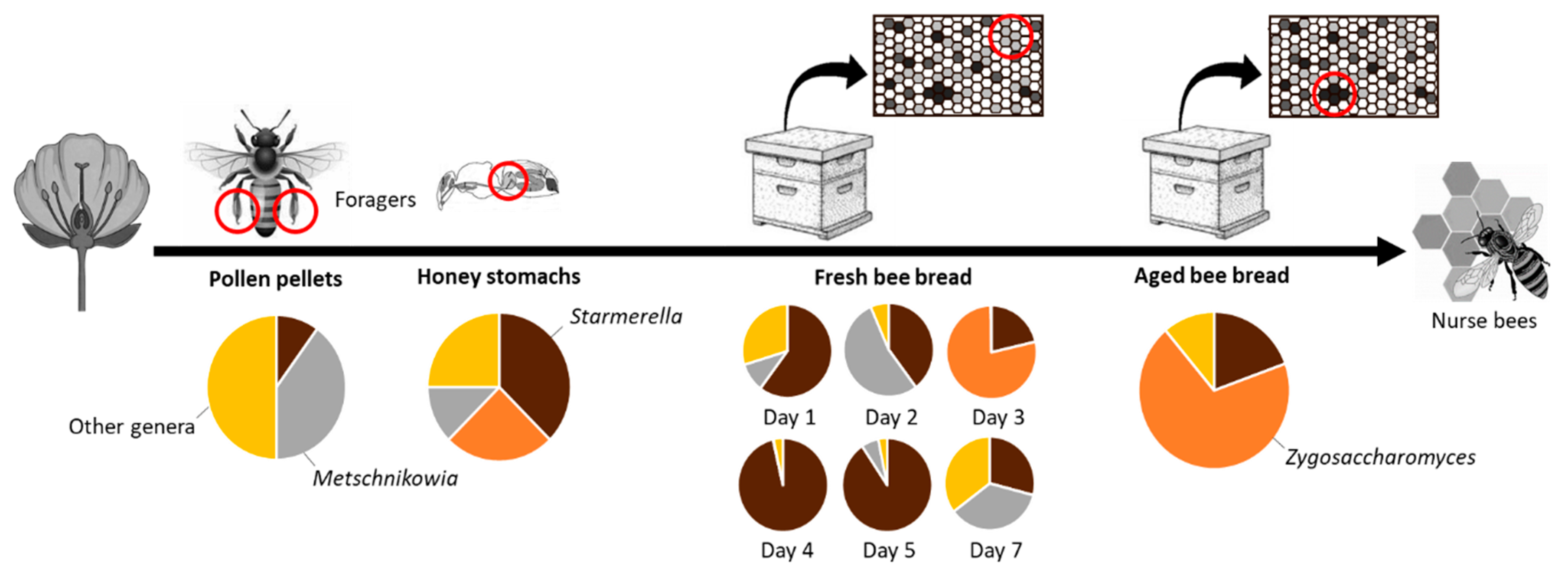
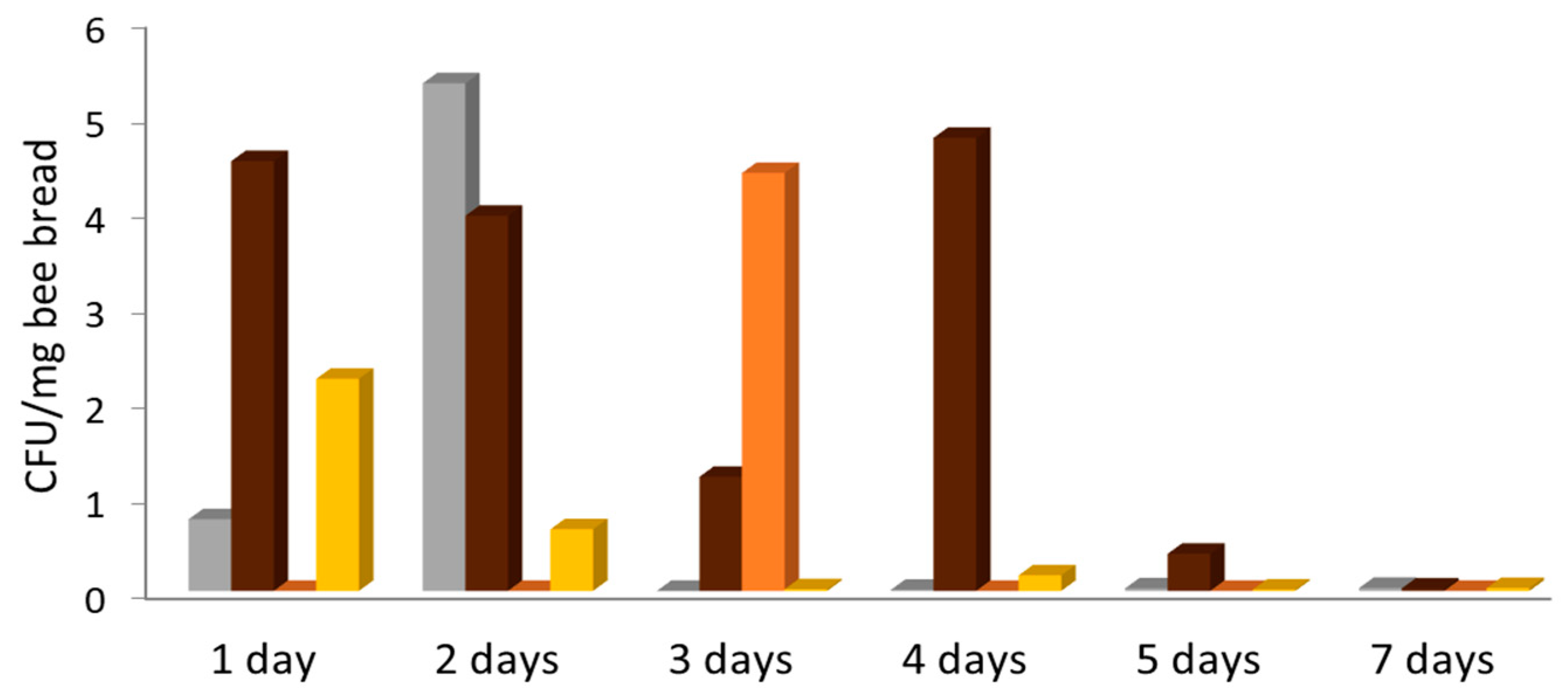
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Haydak, M.H. Honey bee nutrition. Annu. Rev. Entomol. 1970, 15, 143–156. [Google Scholar] [CrossRef]
- Brodschneider, R.; Crailsheim, K. Nutrition and health in honey bees. Apidologie 2010, 41, 278–294. [Google Scholar] [CrossRef]
- Parker, R.L. The Collection and Utilization of Pollen by the Honeybee. Ph.D. Thesis, Cornell University, Ithaca, NY, USA, September 1925. [Google Scholar]
- Podrižnik, B.; Božič, J. Maturation and stratification of antibacterial activity and total phenolic content of bee bread in honey comb cells. J. Apic. Res. 2015, 54, 81–92. [Google Scholar] [CrossRef]
- Vanengelsdorp, D.; Evans, J.D.; Donovall, L.; Mullin, C.; Frazier, M.; Frazier, J.; Tarpy, D.R.; Hayes, J.; Pettis, J.S. “Entombed Pollen”: A new condition in honey bee colonies associated with increased risk of colony mortality. J. Invertebr. Pathol. 2009, 101, 147–149. [Google Scholar] [CrossRef]
- Roulston, T.H.; Cane, J.H. Pollen nutritional content and digestibility for animals. Plant. Syst. Evol. 2000, 222, 187–209. [Google Scholar] [CrossRef]
- Roulston, T.H.; Cane, J.H.; Buchmann, S.L. What governs protein content of pollen: Pollinator preferences, pollen-pistil interactions, or phylogeny? Ecol. Monogr. 2000, 70, 617. [Google Scholar] [CrossRef]
- Pain, J.; Maugenet, J. Recherches biochimiques et physiologiques sur le pollen emmagasiné par les abeilles. Apidologie 1966, 9, 209–236. [Google Scholar] [CrossRef][Green Version]
- Herbert, E.W., Jr.; Shimanuki, H. Chemical composition and nutritive value of bee-collected and bee-stored pollen. Apidologie 1978, 9, 33–40. [Google Scholar] [CrossRef]
- DeGrandi-Hoffman, G.; Eckholm, B.J.; Huang, M.H. A comparison of bee bread made by Africanized and European honey bees (Apis mellifera) and its effects on hemolymph protein titers. Apidologie 2012, 44, 52–63. [Google Scholar] [CrossRef]
- Beutler, R.; Opfinger, E. Pollenernährung und nosemabefall der honigbiene (Apis Mellifica). J. Comp. Physiol. A 1950, 32, 383–421. [Google Scholar] [CrossRef]
- Cremonez, T.M.; De Jong, D.; Bitondi, M.M.G. Quantification of hemolyph proteins as a fast method for testing protein diets for honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 1998, 91, 1284–1289. [Google Scholar] [CrossRef]
- Anderson, K.E.; Carroll, M.J.; Sheehan, T.; Mott, B.M.; Maes, P.; Corby-Harris, V. Hive-stored pollen of honey bees: Many lines of evidence are consistent with pollen preservation, not nutrient conversion. Mol. Ecol. 2014, 23, 5904–5917. [Google Scholar] [CrossRef]
- Carroll, M.J.; Brown, N.; Goodall, C.; Downs, A.M.; Sheenan, T.H.; Anderson, K.E. Honey bees preferentially consume freshly-stored pollen. PLoS ONE 2017, 12, e0175933. [Google Scholar] [CrossRef]
- Maes, P.; Rodrigues, P.A.P.; Oliver, R.; Mott, B.M.; Anderson, K.E. Diet-related gut bacterial dysbiosis correlates with impaired development, increased mortality and Nosema disease in the honeybee (Apis mellifera). Mol. Ecol. 2016, 25, 5439–5450. [Google Scholar] [CrossRef]
- Vásquez, A.; Olofsson, T.C. The lactic acid bacteria involved in the production of bee pollen and bee bread. J. Apic. Res. 2009, 48, 189–195. [Google Scholar] [CrossRef]
- De Vuyst, L.; Lefeber, T.; Papalexandratou, Z.; Camu, N. The functional role of lactic acid bacteria in cocoa bean fermentation. In Biotechnology of Lactic Acid Bacteria; Wiley: Hoboken, NJ, USA, 2010; pp. 301–325. [Google Scholar]
- Chevtchik, V. Mikrobiologie pylového kvašení. Publ. Fac. Sci. Univ. Masaryk 1950, 323, 103–130. [Google Scholar]
- Gilliam, M. Microbiology of pollen and bee bread: The yeasts. Apidologie 1979, 10, 43–53. [Google Scholar] [CrossRef]
- Moran, N.A.; Hansen, A.K.; Powell, J.E.; Sabree, Z.L. Distinctive gut microbiota of honey bees assessed using deep sampling from individual worker bees. PLoS ONE 2012, 7, e36393. [Google Scholar] [CrossRef]
- Anderson, K.E.; Sheehan, T.H.; Mott, B.M.; Maes, P.; Snyder, L.; Schwan, M.R.; Walton, A.; Jones, B.M.; Corby-Harris, V. Microbial ecology of the hive and pollination landscape: Bacterial associates from floral nectar, the alimentary tract and stored food of honey bees (Apis mellifera). PLoS ONE 2013, 8, e83125. [Google Scholar] [CrossRef]
- Saraiva, M.A.; Zemolin, A.P.P.; Franco, J.L.; Boldo, J.T.; Stefenon, V.M.; Triplett, E.W.; Camargo, F.A.D.O.; Roesch, L.F.W. Relationship between honeybee nutrition and their microbial communities. Antonie van Leeuwenhoek 2015, 107, 921–933. [Google Scholar] [CrossRef]
- Khan, K.A.; Al-Ghamdi, A.A.; Ghramh, H.A.; Ansari, M.J.; Ali, H.; Alamri, S.A.; Kahtani, S.N.A.; Adgaba, N.; Qasim, M.; Hafeez, M. Structural diversity and functional variability of gut microbial communities associated with honey bees. Microb. Pathog. 2020, 138, 103793. [Google Scholar] [CrossRef]
- Cornman, R.S.; Tarpy, D.R.; Chen, Y.; Jeffreys, L.; Lopez, D.; Pettis, J.S.; Vanengelsdorp, D.; Evans, J.D. Pathogen webs in collapsing honey bee colonies. PLoS ONE 2012, 7, e43562. [Google Scholar] [CrossRef]
- Yun, J.-H.; Jung, M.-J.; Kim, P.S.; Bae, J.-W. Social status shapes the bacterial and fungal gut communities of the honey bee. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef]
- Pozo, I.M.; Van Kemenade, G.; Van Oystaeyen, A.; Aledón-Catalá, T.; Benavente, A.; Ende, W.V.D.; Wäckers, F.; Jacquemyn, H. The impact of yeast presence in nectar on bumble bee behavior and fitness. Ecol. Monogr. 2019, 90, 01393. [Google Scholar] [CrossRef]
- Carreck, N.L.; Andree, M.; Brent, C.S.; Cox-Foster, D.; Dade, A.H.; Ellis, J.D.; Hatjina, F.; Van Englesdorp, D. Standard methods forApis melliferaanatomy and dissection. J. Apic. Res. 2013, 52, 1–40. [Google Scholar] [CrossRef]
- Hennebert, G. (Université catholique de Louvain, Louvain-la-Neuve, Belgium). Personal communication/Internal quality document I013N, 2019.
- Hocking, A.D.; Pitt, I.J. Dichloran-glycerol medium for enumeration of xerophilic fungi from low-moisture foods. Appl. Environ. Microbiol. 1980, 39, 488–492. [Google Scholar] [CrossRef]
- Moncalvo, J.-M.; Lutzoni, F.M.; Rehner, S.A.; Johnson, J.; Vilgalys, R. Phylogenetic relationships of agaric fungi based on nuclear large subunit ribosomal DNA sequences. Syst. Biol. 2000, 49, 278–305. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.; Taylor, J. 38-Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Daniel, H. Evaluation of ribosomal RNA and actin gene sequences for the identification of ascomycetous yeasts. Int. J. Food Microbiol. 2003, 86, 61–78. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: molecular evolutionary genetics analysis version 6. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Sipiczki, M. Metschnikowia pulcherrima and related pulcherrimin-producing yeasts: Fuzzy species boundaries and complex antimicrobial antagonism. Microorganisms 2020, 8, 1029. [Google Scholar] [CrossRef]
- Lachance, M.-A. Metschnikowia: Half tetrads, a regicide and the fountain of youth. Yeast 2016, 33, 563–574. [Google Scholar] [CrossRef]
- Lachance, M.-A. Starmerella Rosa & Lachance (1998). In The Yeasts: A Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; Volume 2, pp. 811–815. [Google Scholar]
- Lachance, M.-A.; Dobson, J.; Wijayanayaka, D.N.; Smith, A.M.E. The use of parsimony network analysis for the formal delineation of phylogenetic species of yeasts: Candida apicola, Candida azyma, and Candida parazyma sp. nov., cosmopolitan yeasts associated with floricolous insects. Antonie van Leeuwenhoek 2010, 97, 155–170. [Google Scholar] [CrossRef]
- Haydak, M.H. Value of pollen substitutes for brood rearing of honeybees. J. Econ. Entomol. 1945, 38, 484–487. [Google Scholar] [CrossRef]
- Rosa, C.A.; Lachance, M.-A.; Silva, J.O.; Teixeira, A.C.P.; Marini, M.M.; Antonini, Y.; Martins, R.P. Yeast communities associated with stingless bees. FEMS Yeast Res. 2003, 4, 271–275. [Google Scholar] [CrossRef]
- Lachance, M.-A.; Starmer, W.T.; Rosa, C.A.; Bowles, J.M.; Barker, J.F.; Janzen, D.H. Biogeography of the yeasts of ephemeral flowers and their insects. FEMS Yeast Res. 2001, 1, 1–8. [Google Scholar] [CrossRef]
- Inglis, G.; Sigler, L.; Goette, M. Aerobic microorganisms associated with alfalfa leafcutter bees (Megachile rotundata). Microb. Ecol. 1993, 26, 125–143. [Google Scholar] [CrossRef]
- Spencer, J.F.T.; Gorin, P.A.J.; Hobbs, G.A.; Cooke, D.A. Yeasts isolated from bumblebee honey from Western Canada: Identification with the aid of proton magnetic resonance spectra of their mannose-containing polysaccharides. Can. J. Microbiol. 1970, 16, 117–119. [Google Scholar] [CrossRef]
- Egorova, A.I.; Bab’eva, I.P. Yeast flora of the honey bee (Apis mellifica L.). Izvestija Sibirskogo otdelenija Akademii nauk SSSR. Ser. Biol. Med. Nauk 1967, 2, 127–132. [Google Scholar]
- Teixeira, A.C.P.; Marini, M.M.; Nicoli, J.R.; Antonini, Y.; Martins, R.P.; Lachance, M.-A.; Rosa, C.A. Starmerella meliponinorum sp. nov., a novel ascomycetous yeast species associated with stingless bees. Int. J. Syst. Evol. Microbiol. 2003, 53, 339–343. [Google Scholar] [CrossRef]
- Pimentel, M.R.; Antonini, Y.; Martins, R.P.; Lachance, M.-A.; Rosa, C.A. Candida riodocensis and Candida cellae, two new yeast species from the clade associated with solitary bees in the Atlantic rain forest of Brazil. FEMS Yeast Res. 2005, 5, 875–879. [Google Scholar] [CrossRef] [PubMed]
- Daniel, H.-M.; Rosa, C.A.; Thiago-Calaça, P.S.S.; Antonini, Y.; Bastos, E.M.A.F.; Evrard, P.; Huret, S.; Fidalgo-Jiménez, A.; Lachance, M.-A. Starmerella neotropicalis f. a., sp. nov., a yeast species found in bees and pollen. Int. J. Syst. Evol. Microbiol. 2013, 63, 3896–3903. [Google Scholar] [CrossRef] [PubMed]
- Rosa, C.A.; Viana, E.M.; Martins, R.P.; Antonini, Y.; Lachance, M.-A. Candida batistae, a new yeast species associated with solitary digger nesting bees in Brazil. Mycologia 1999, 91, 428. [Google Scholar] [CrossRef]
- Monrocher, R. Quelques nouvelles espèces et variétés du genre Candida (levures asporogènes). Rev. Mycol. 1967, 32, 69–92. [Google Scholar]
- Brysch-Herzberg, M. Ecology of yeasts in plant–bumblebee mutualism in Central Europe. FEMS Microbiol. Ecol. 2004, 50, 87–100. [Google Scholar] [CrossRef]
- Dharampal, P.S.; Diaz-Garcia, L.; Haase, M.A.B.; Zalapa, J.E.; Currie, C.R.; Hittinger, C.T.; Steffan, S.A. Microbial diversity associated with the pollen stores of captive-bred bumble bee colonies. Insects 2020, 11, 250. [Google Scholar] [CrossRef]
- Grilione, P.; Federici, F.; Miller, M. Yeasts from honey bees (Apis Mellifera L.). In Advances in Biotechnology, Proceedings of the Fifth International Yeast Symposium, London, Canada, July 20–25 1980; Elsevier BV: Amsterdam, The Netherlands, 1981; pp. 599–605. [Google Scholar] [CrossRef]
- Lavie, P. Essais de lutte biologique contre l’Acarapis woodi, agent de l’acariose de l’abeille. C. R. Hebd. Acad. Sci. 1954, 238, 947–948. [Google Scholar]
- Neto, D.J.D.C.; De Morais, P.B. The vectoring of Starmerella species and other yeasts by stingless bees in a Neotropical savanna. Fungal Ecol. 2020, 47, 100973. [Google Scholar] [CrossRef]
- Parle, J.N.; Di Menna, M.E. The source of yeasts in New Zealand wines. N. Z. J. Agric. Res. 1966, 9, 98–107. [Google Scholar] [CrossRef]
- Rosa, C.A.; Lachance, M.-A. The yeast genus Starmerella gen. nov. and Starmerella bombicola sp. nov., the teleomorph of Candida bombicola (Spencer, Gorin & Tullock) Meyer & Yarrow. Int. J. Syst. Bacteriol. 1998, 48, 1413–1417. [Google Scholar] [CrossRef]
- Jones, D.F. Novel macrocyclic glycolipids from Torulopsis gropengiesseri. J. Chem. Soc. C 1967, 6, 479–484. [Google Scholar] [CrossRef]
- Konishi, M.; Fukuoka, T.; Morita, T.; Imura, T.; Kitamoto, D. Production of new types of sophorolipids by Candida batistae. J. Oleo Sci. 2008, 57, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P.; Price, N.P.; Ray, K.J.; Kuo, T.-M. Production of sophorolipid biosurfactants by multiple species of the Starmerella (Candida) bombicola yeast clade. FEMS Microbiol. Lett. 2010, 311, 140–146. [Google Scholar] [CrossRef] [PubMed]
- Konishi, M.; Morita, T.; Fukuoka, T.; Imura, T.; Uemura, S.; Iwabuchi, H.; Kitamoto, D. efficient production of acid-form sophorolipids from waste glycerol and fatty acid methyl esters by Candida floricola. J. Oleo Sci. 2018, 67, 489–496. [Google Scholar] [CrossRef]
- Hommel, R.; Weber, L.; Weiss, A.; Himmelreich, U.; Rilke, O.; Kleber, H.-P. Production of sophorose lipid by Candida (Torulopsis) apicola grown on glucose. J. Biotechnol. 1994, 33, 147–155. [Google Scholar] [CrossRef]
- Van Bogaert, I.N.A.; Saerens, K.; De Muynck, C.; Develter, D.; Soetaert, W.; Vandamme, E.J. Microbial production and application of sophorolipids. Appl. Microbiol. Biotechnol. 2007, 76, 23–34. [Google Scholar] [CrossRef]
- Marchant, R.; Banat, I.M. Microbial biosurfactants: Challenges and opportunities for future exploitation. Trends Biotechnol. 2012, 30, 558–565. [Google Scholar] [CrossRef]
- Pacini, E.; Hesse, M. Pollenkitt—its composition, forms and functions. Flora Morphol. Distrib. Funct. Ecol. Plants 2005, 200, 399–415. [Google Scholar] [CrossRef]
- James, S.A.; Stratford, M. Zygosaccharomyces Barker (1901). In The Yeasts: A Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: Amsterdam, The Netherlands, 1998; Volume 2, pp. 937–947. [Google Scholar]
- Solieri, L.; Dakal, T.C.; Giudici, P. Zygosaccharomyces sapae sp. nov., isolated from Italian traditional balsamic vinegar. Int. J. Syst. Evol. Microbiol. 2013, 63, 364–371. [Google Scholar] [CrossRef][Green Version]
- Sinpoo, C.; Williams, G.R.; Chantawannakul, P. Dynamics of fungal communities in corbicular pollen and bee bread. Chiang Mai J. Sci. 2017, 44, 1244–1256. [Google Scholar]
- Čadež, N.; Fülöp, L.; Dlauchy, D.; Péter, G. Zygosaccharomyces favi sp. nov., an obligate osmophilic yeast species from bee bread and honey. Antonie van Leeuwenhoek 2014, 107, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Péter, G.; National Collection of Agricultural and Industrial Microorganisms (NCAIM), Szent István University, Budapest, Hungary. Personal communication, 2017.
- James, S.A.; Stratford, M. Spoilage yeasts with emphasis on the genus Zygosaccharomyces. In Yeasts in Food; Elsevier BV: Amsterdam, The Netherlands, 2003; pp. 171–191. [Google Scholar]
- Grüẞ, J. Die Anpassung eines Pilzes (Anthomyces Reukaufii) an den Blütenbau und den Bienenrüssel. Ber. Deut. Bot. Ges. 1918, 35, 746–762. [Google Scholar]
- Pozo, M.I.; Lachance, M.-A.; Herrera, C.M. Nectar yeasts of two southern Spanish plants: The roles of immigration and physiological traits in community assembly. FEMS Microbiol. Ecol. 2012, 80, 281–293. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, B.; Lachance, M.-A.; Herrera, C.M. Phylogenetic analysis of the angiosperm-floricolous insect–yeast association: Have yeast and angiosperm lineages co-diversified? Mol. Phylogenetics Evol. 2013, 68, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Kakumanu, M.L.; Reeves, A.M.; Anderson, T.D.; Rodrigues, R.R.; Williams, M.A. Honey bee gut microbiome is altered by in-hive pesticide exposures. Front. Microbiol. 2016, 7, 1255. [Google Scholar] [CrossRef]
- Stefanini, I.; Dapporto, L.; Legras, J.-L.; Calabretta, A.; Di Paola, M.; De Filippo, C.; Viola, R.; Capretti, P.; Polsinelli, M.; Turillazzi, S.; et al. Role of social wasps in Saccharomyces cerevisiae ecology and evolution. Proc. Natl. Acad. Sci. USA 2012, 109, 13398–13403. [Google Scholar] [CrossRef]
- Hajsig, M. Contribution to the knowledge of yeasts from the intestinal content of bees. Vet. Arhiv 1959, 29, 145–156. [Google Scholar]
- Sandhu, D.K.; Waraich, M.K. Yeasts associated with pollinating bees and flower nectar. Microb. Ecol. 1985, 11, 51–58. [Google Scholar] [CrossRef]
- Batra, L.R.; Batra, S.W.T.; Bohart, G.E. The mycoflora of domesticated and wild bees (Apoidea). Mycopathol. 1973, 49, 13–44. [Google Scholar] [CrossRef]
- Burnside, C.E. Fungous Disease of the Honeybee; Technical Report for United States Department of Agriculture: Washington, DC, USA, 1930; Volume 149, pp. 1–41. [Google Scholar]
- Gilliam, M.; Wickerham, L.; Morton, H.L.; Martin, R.D. Yeasts isolated from honey bees, Apis mellifera, fed 2,4-D and antibiotics. J. Invertebr. Pathol. 1974, 24, 349–356. [Google Scholar] [CrossRef]
- Gilliam, M.; Morton, H.L.; Prest, D.B.; Martin, R.D.; Wickerham, L. The mycoflora of adult worker honeybees, Apis mellifera: Effects of 2,4,5-T and caging of bee colonies. J. Invertebr. Pathol. 1977, 30, 50–54. [Google Scholar] [CrossRef]
- Borsuk, G.; Ptaszyńska, A.A.; Olszewski, K.; Paleolog, J. Impact of nosemosis on the intestinal yeast flora of honey bees. Med. Weter. 2013, 69, 726–729. [Google Scholar]
- Torto, B.; Boucias, I.G.; Arbogast, R.T.; Tumlinson, J.H.; Teal, P.E.A. Multitrophic interaction facilitates parasite-host relationship between an invasive beetle and the honey bee. Proc. Natl. Acad. Sci. USA 2007, 104, 8374–8378. [Google Scholar] [CrossRef]
- Benda, N.D. Detection and characterization of kodamaea ohmeri associated with small hive beetle Aethina tumida infesting honey bee hives. J. Apic. Res. 2008, 47, 194–201. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Composition | Application |
|---|---|---|
| DYPA | Glucose 2% (Rocc, Sart-Eustache, Belgium) | General |
| Dextrose Yeast Extract Peptone Agar | Peptone 1% (Duchefa, Haarlem, Netherlands) | |
| Yeast extract 0.5% (Oxoid, Basingstoke, United Kingdom) | ||
| Agar 2% (Rocc, Sart-Eustache, Belgium) | ||
| MA2 | Malt extract 2% (Duchefa, Haarlem, Netherlands) | General, rich in complex nutrients |
| Malt Agar 2% | Agar 1.5% | |
| MYAG50 | Glucose 50% | Osmophiles |
| Malt Yeast Agar Glucose 50% | Peptone 0.5% | |
| Yeast extract 0.3% | ||
| Malt extract 0.3% | ||
| Agar 2% | ||
| HYA | Honey without preservatives 1 3% | Basidiomycetous bee yeasts |
| Honey Yeast Agar [28] | Yeast extract 0.5% | |
| Agar 2% | ||
| DG18 | Glucose 1% | Xerophilic fungi and yeasts |
| Dichloran-Glycerol Agar [29] | Peptone 0.5% | |
| Dipotassium phosphate 0.1% (VWR, Radnor, USA) | ||
| Magnesium sulfate 0.05% (VWR, Radnor, USA) | ||
| Chloramphenicol 0.01% (AppliChem, Darmstadt, Germany) | ||
| Glycerol 22% (VWR, Radnor, USA) | ||
| Agar 1.5% |
| Fresh Bee Bread 1 n = 28 | Aged Bee Bread 2 n = 107 | Pollen Pellets n = 12 | Honey Stomach n = 14 | |||||
|---|---|---|---|---|---|---|---|---|
| Samples | Colonies 3 | Samples | Colonies 3 | Samples | Colonies | Samples | Colonies | |
| Metschnikowia | ||||||||
| Metschnikowia (Candida) rancensis | 3 | 100 | 1 | 50 | ||||
| Metschnikowia cf. pulcherrima | 9 | 74.4 | 1 | 10 | ||||
| Starmerella | ||||||||
| Starmerella (Candida) apicola | 10 | 36.5 | 1 | 0.7 | ||||
| Starmerella (Candida) apis | 18 | 229.8 | ||||||
| Starmerella (Candida) bombi | 2 | 4.7 | 2 | 2 | 1 | 1 | ||
| Starmerella (Candida) magnoliae | 10 | 62.4 | 1 | 2 | 1 | 2 | 2 | 5 |
| Starmerella bombicola | 5 | 26.5 | 3 | >200 | ||||
| Zygosaccharomyces | ||||||||
| Zygosaccharomyces favi | 6 | 3.3 | ||||||
| Zygosaccharomyces mellis | 2 | 40.3 | 3 | 121 | ||||
| Zygosaccharomyces rouxii | 2 | 46 | 6 | 24.3 | 1 | 1 | ||
| Zygosaccharomyces sapae | 5 | 102 | ||||||
| Zygosaccharomyces siamensis | 1 | 0.3 | 5 | 14.3 | 1 | 2 | ||
| Other | ||||||||
| Aureobasidium pullulans | 1 | 0.7 | ||||||
| Candida parapsilosis | 1 4 | |||||||
| Debaryomyces hansenii | 9 | 5 | ||||||
| Debaryomyces sp. | 2 | 0.7 | ||||||
| Debaryomyces maramus | 2 | 4 | ||||||
| Dothiora prunorum | 2 | 0.6 | ||||||
| Hanseniaspora uvarum | 2 | 4 | 1 | 5 | ||||
| Kluyveromyces dobzhanskii | 1 | 3 | 1 | 3 | ||||
| Kodamaea ohmeri | 8 | 35.4 | ||||||
| Meyerozyma guilliermondii | 1 | 0.7 | ||||||
| Naganishia (Cryptococcus) diffluens | 1 | 1 | ||||||
| Rhodotorula mucilaginosa | 2 4 | |||||||
| Saccharomyces cerevisiae | 1 | >200 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Detry, R.; Simon-Delso, N.; Bruneau, E.; Daniel, H.-M. Specialisation of Yeast Genera in Different Phases of Bee Bread Maturation. Microorganisms 2020, 8, 1789. https://doi.org/10.3390/microorganisms8111789
Detry R, Simon-Delso N, Bruneau E, Daniel H-M. Specialisation of Yeast Genera in Different Phases of Bee Bread Maturation. Microorganisms. 2020; 8(11):1789. https://doi.org/10.3390/microorganisms8111789
Chicago/Turabian StyleDetry, Roxane, Noa Simon-Delso, Etienne Bruneau, and Heide-Marie Daniel. 2020. "Specialisation of Yeast Genera in Different Phases of Bee Bread Maturation" Microorganisms 8, no. 11: 1789. https://doi.org/10.3390/microorganisms8111789
APA StyleDetry, R., Simon-Delso, N., Bruneau, E., & Daniel, H.-M. (2020). Specialisation of Yeast Genera in Different Phases of Bee Bread Maturation. Microorganisms, 8(11), 1789. https://doi.org/10.3390/microorganisms8111789