Multinucleated Giant Cell Formation as a Portal to Chronic Bacterial Infections


{kind=link}
Abstract
1. Introduction
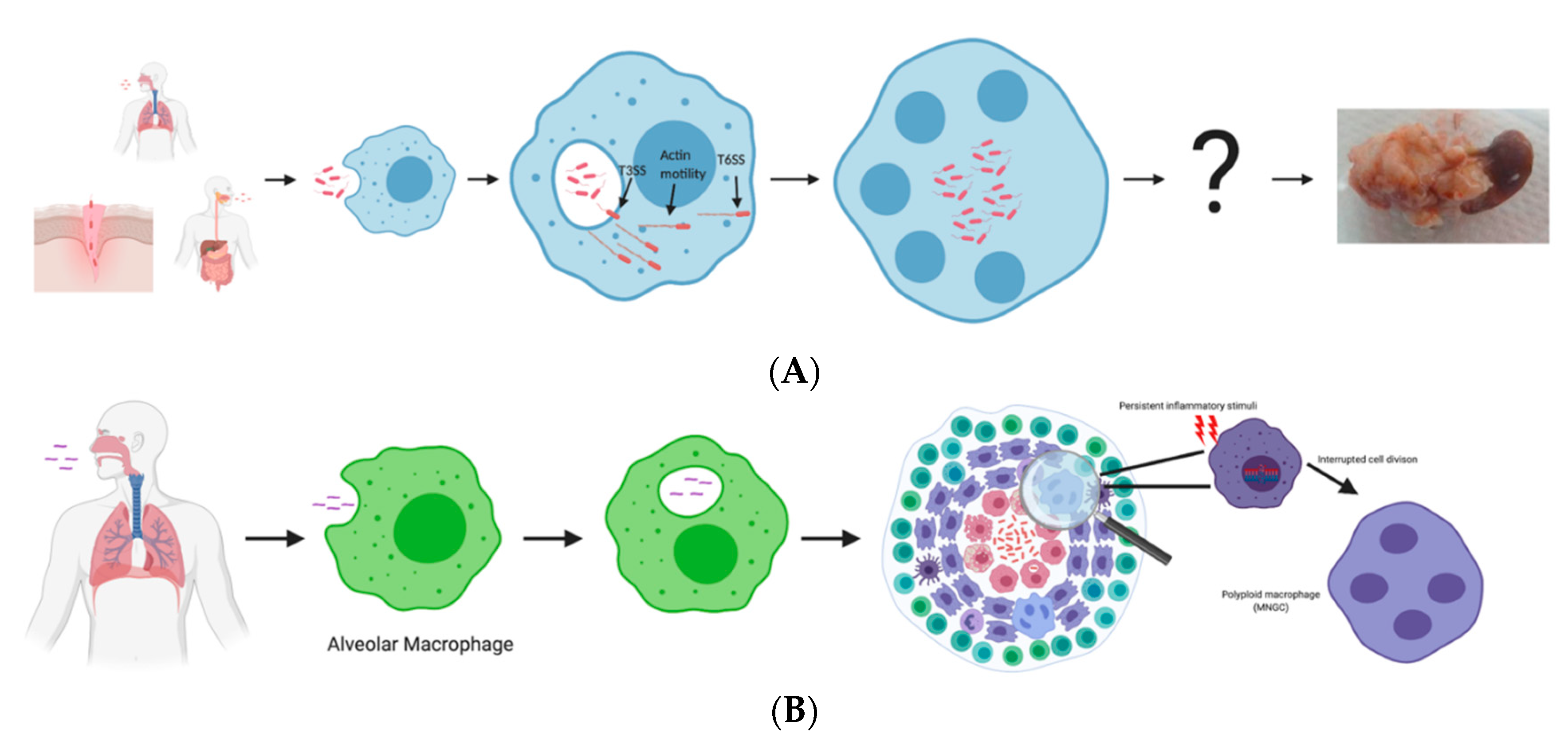
2. Bpm Pathogenesis Process Leading to MNGC Formation
3. Similarities and Differences between Melioidosis and Tuberculosis
4. Role of MNGCs in Disease
5. Future Directions and Models
6. Final Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Limmathurotsakul, D.; Golding, N.; Dance, D.A.B.; Messina, J.P.; Pigott, D.M.; Moyes, C.L.; Rolim, D.R.; Bertherat, E.; Day, N.P.J.; Peacock, S.J.; et al. Predicted global distribution of Burkholderia pseudomallei and burden of melioidosis. Nat. Microbiol. 2016, 1, 15008. [Google Scholar] [CrossRef] [PubMed]
- Cossaboom, C.M.; Marinova-Petkova, A.; Strysko, J.; Rodriguez, G.; Maness, T.; Ocampo, J.; Gee, J.E.; Elrod, M.G.; Gulvik, C.A.; Liu, L.; et al. Melioidosis in a Resident of Texas with No Recent Travel History, United States. Emerg. Infect. Dis. 2020, 26, 1295–1299. [Google Scholar] [CrossRef] [PubMed]
- Wiersinga, W.J.; Virk, H.S.; Torres, A.G.; Currie, B.J.; Peacock, S.J.; Dance, D.A.B.; Limmathurotsakul, D. Melioidosis. Nat. Rev. Dis. Primers 2018, 4, 17107. [Google Scholar] [CrossRef] [PubMed]
- Amemiya, K.; Dankmeyer, J.L.; Bearss, J.J.; Zeng, X.; Stonier, S.W.; Soffler, C.; Cote, C.K.; Welkos, S.L.; Fetterer, D.P.; Chance, T.B.; et al. Dysregulation of TNF-α and IFN-γ expression is a common host immune response in a chronically infected mouse model of melioidosis when comparing multiple human strains of Burkholderia pseudomallei. BMC Immunol. 2020, 21, 5. [Google Scholar] [CrossRef]
- Garg, R.; Shaw, T.; Vandana, K.E.; Magazine, R.; Mukhopadhyay, C. Melioidosis In Suspected Recurrent Tuberculosis: A disease in disguise. J. Infect. Dev. Ctries 2020, 14, 312–316. [Google Scholar] [CrossRef]
- Ninan, F.; Mishra, A.K.; John, A.O.; Iyadurai, R. Splenic Granuloma: Melioidosis or Tuberculosis? J. Family Med. Prim. Care 2018, 7, 271–273. [Google Scholar] [CrossRef]
- Wong, J.; Chen, Y.; Gan, Y.H. Host Cytosolic Glutathione Sensing by a Membrane Histidine Kinase Activates the Type Vi Secretion System in an Intracellular Bacterium. Cell Host Microbe 2015, 18, 38–48. [Google Scholar] [CrossRef]
- Kespichayawattana, W.; Rattanachetkul, S.; Wanun, T.; Utaisincharoen, P.; Sirisinha, S. Burkholderia pseudomallei induces cell fusion and actin-associated membrane protrusion: A possible mechanism for cell-to-cell spreading. Infect. Immun. 2000, 68, 5377–5384. [Google Scholar] [CrossRef]
- Whiteley, L.; Meffert, T.; Haug, M.; Weidenmaier, C.; Hopf, V.; Bitschar, K.; Schittek, B.; Kohler, C.; Steinmetz, I.; West, T.E.; et al. Entry, Intracellular Survival, and Multinucleated-Giant-Cell-Forming Activity of Burkholderia pseudomallei in Human Primary Phagocytic and Nonphagocytic Cells. Infect. Immun. 2017, 85, e00468. [Google Scholar] [CrossRef]
- Walkden, H.; Delbaz, A.; Nazareth, L.; Batzloff, M.; Shelper, T.; Beacham, I.R.; Chacko, A.; Shah, M.; Beagley, K.W.; Tello Velasquez, J.; et al. Burkholderia pseudomallei invades the olfactory nerve and bulb after epithelial injury in mice and causes the formation of multinucleated giant glial cells in vitro. PLoS Negl. Trop. Dis. 2020, 14, e0008017. [Google Scholar] [CrossRef]
- Stevens, M.P.; Wood, M.W.; Taylor, L.A.; Monaghan, P.; Hawes, P.; Jones, P.W.; Wallis, T.S.; Galyov, E.E. An Inv/Mxi-Spa-like type III protein secretion system in Burkholderia pseudomallei modulates intracellular behaviour of the pathogen. Mol. Microbiol. 2002, 46, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Chen, Y.; Ouyang, X.; Gan, Y.H. Identification of tomato plant as a novel host model for Burkholderia pseudomallei. BMC Microbiol. 2010, 10, 28. [Google Scholar] [CrossRef]
- Angus, A.A.; Agapakis, C.M.; Fong, S.; Yerrapragada, S.; Estrada-de los Santos, P.; Yang, P.; Song, N.; Kano, S.; Caballero-Mellado, J.; de Faria, S.M.; et al. Plant-associated symbiotic Burkholderia species lack hallmark strategies required in mammalian pathogenesis. PLoS ONE 2014, 9, e83779. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.P.; Haque, A.; Atkins, T.; Hill, J.; Wood, M.W.; Easton, A.; Nelson, M.; Underwood-Fowler, C.; Titball, R.W.; Bancroft, G.J.; et al. Attenuated virulence and protective efficacy of a Burkholderia pseudomallei bsa type III secretion mutant in murine models of melioidosis. Microbiology (Read.) 2004, 150, 2669–2676. [Google Scholar] [CrossRef]
- Burtnick, M.N.; Brett, P.J.; Nair, V.; Warawa, J.M.; Woods, D.E.; Gherardini, F.C. Burkholderia pseudomallei type III secretion system mutants exhibit delayed vacuolar escape phenotypes in RAW 264.7 murine macrophages. Infect. Immun. 2008, 76, 2991–3000. [Google Scholar] [CrossRef]
- Vander Broek, C.W.; Zainal Abidin, N.; Stevens, J.M. BipC, a Predicted Burkholderia pseudomallei Type 3 Secretion System Translocator Protein with Actin Binding Activity. Front. Cell Infect. Microbiol. 2017, 7, 333. [Google Scholar] [CrossRef]
- Jitprasutwit, S.; Jitprasutwit, N.; Hemsley, C.M.; Onlamoon, N.; Withatanung, P.; Muangsombut, V.; Vattanaviboon, P.; Stevens, J.M.; Ong, C.; Stevens, M.P.; et al. Identification of Burkholderia pseudomallei Genes Induced During Infection of Macrophages by Differential Fluorescence Induction. Front. Microbiol. 2020, 11, 72. [Google Scholar] [CrossRef]
- Stolt, C.; Schmidt, I.H.; Sayfart, Y.; Steinmetz, I.; Bast, A. Heme Oxygenase-1 and Carbon Monoxide Promote Burkholderia pseudomallei Infection. J. Immunol. 2016, 197, 834–846. [Google Scholar] [CrossRef]
- Stevens, M.P.; Stevens, J.M.; Jeng, R.L.; Taylor, L.A.; Wood, M.W.; Hawes, P.; Monaghan, P.; Welch, M.D.; Galyov, E.E. Identification of a bacterial factor required for actin-based motility of Burkholderia pseudomallei. Mol. Microbiol. 2005, 56, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Srinon, V.; Chaiwattanarungruengpaisan, S.; Korbsrisate, S.; Stevens, J.M. Burkholderia pseudomallei BimC Is Required for Actin-Based Motility, Intracellular Survival, and Virulence. Front. Cell Infect. Microbiol. 2019, 9, 63. [Google Scholar] [CrossRef]
- Kang, W.T.; Vellasamy, K.M.; Chua, E.G.; Vadivelu, J. Functional characterizations of effector protein BipC, a type III secretion system protein, in Burkholderia pseudomallei pathogenesis. J. Infect. Dis. 2015, 211, 827–834. [Google Scholar] [CrossRef]
- Shalom, G.; Shaw, J.G.; Thomas, M.S. In vivo expression technology identifies a type VI secretion system locus in Burkholderia pseudomallei that is induced upon invasion of macrophages. Microbiology (Read.) 2007, 153, 2689–2699. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, S.; West, T.E.; Boyer, F.; Chiang, W.C.; Carl, M.A.; Hood, R.D.; Rohmer, L.; Tolker-Nielsen, T.; Skerrett, S.J.; Mougous, J.D. Burkholderia type VI secretion systems have distinct roles in eukaryotic and bacterial cell interactions. PLoS Pathog. 2010, 6, e1001068. [Google Scholar] [CrossRef] [PubMed]
- Burtnick, M.N.; DeShazer, D.; Nair, V.; Gherardini, F.C.; Brett, P.J. Burkholderia mallei cluster 1 type VI secretion mutants exhibit growth and actin polymerization defects in RAW 264.7 murine macrophages. Infect. Immun. 2010, 78, 88–99. [Google Scholar] [CrossRef]
- Pilatz, S.; Breitbach, K.; Hein, N.; Fehlhaber, B.; Schulze, J.; Brenneke, B.; Eberl, L.; Steinmetz, I. Identification of Burkholderia pseudomallei genes required for the intracellular life cycle and in vivo virulence. Infect. Immun. 2006, 74, 3576–3586. [Google Scholar] [CrossRef]
- Hopf, V.; Gohler, A.; Eske-Pogodda, K.; Bast, A.; Steinmetz, I.; Breitbach, K. BPSS1504, a cluster 1 type VI secretion gene, is involved in intracellular survival and virulence of Burkholderia pseudomallei. Infect. Immun. 2014, 82, 2006–2015. [Google Scholar] [CrossRef]
- Chen, Y.; Wong, J.; Sun, G.W.; Liu, Y.; Tan, G.Y.; Gan, Y.H. Regulation of type VI secretion system during Burkholderia pseudomallei infection. Infect. Immun. 2011, 79, 3064–3073. [Google Scholar] [CrossRef] [PubMed]
- Gallique, M.; Bouteiller, M.; Merieau, A. The Type VI Secretion System: A Dynamic System for Bacterial Communication? Front. Microbiol. 2017, 8, 1454. [Google Scholar] [CrossRef]
- Toesca, I.J.; French, C.T.; Miller, J.F. The Type VI secretion system spike protein VgrG5 mediates membrane fusion during intercellular spread by pseudomallei group Burkholderia species. Infect. Immun. 2014, 82, 1436–1444. [Google Scholar] [CrossRef]
- Whiteley, L.; Haug, M.; Klein, K.; Willmann, M.; Bohn, E.; Chiantia, S.; Schwarz, S. Cholesterol and host cell surface proteins contribute to cell-cell fusion induced by the Burkholderia type VI secretion system 5. PLoS ONE 2017, 12, e0185715. [Google Scholar] [CrossRef]
- Suparak, S.; Muangsombut, V.; Riyapa, D.; Stevens, J.M.; Stevens, M.P.; Lertmemongkolchai, G.; Korbsrisate, S. Burkholderia pseudomallei-induced cell fusion in U937 macrophages can be inhibited by monoclonal antibodies against host cell surface molecules. Microbes Infect. 2011, 13, 1006–1011. [Google Scholar] [CrossRef] [PubMed]
- Pai, M.; Behr, M.A.; Dowdy, D.; Dheda, K.; Divangahi, M.; Boehme, C.C.; Ginsberg, A.; Swaminathan, S.; Spigelman, M.; Getahun, H.; et al. Tuberculosis. Nat. Rev. Dis. Primers 2016, 2, 16076. [Google Scholar] [CrossRef] [PubMed]
- Martinot, A.J. Microbial Offense vs Host Defense: Who Controls the TB Granuloma? Vet. Pathol. 2018, 55, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Tobin, D.M.; Vary Jr, J.C.; Ray, J.P.; Walsh, G.S.; Dunstan, S.J.; Bang, N.D.; Hagge, D.A.; Khadge, S.; King, M.C.; Hawn, T.R.; et al. The lta4h locus modulates susceptibility to mycobacterial infection in zebrafish and humans. Cell 2010, 140, 717–730. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.O. The structure of mononuclear phagocytes differentiating in vivo. I. Sequential fine and histologic studies of the effect of Bacillus Calmette-Guerin (BCG). Am. J. Pathol. 1974, 76, 17–48. [Google Scholar]
- Cronan, M.R.; Beerman, R.W.; Rosenberg, A.F.; Saelens, J.W.; Johnson, M.G.; Oehlers, S.H.; Sisk, D.M.; Jurcic Smith, K.L.; Medvitz, N.A.; Miller, S.E.; et al. Macrophage Epithelial Reprogramming Underlies Mycobacterial Granuloma Formation and Promotes Infection. Immunity 2016, 45, 861–876. [Google Scholar] [CrossRef]
- Lay, G.; Poquet, Y.; Salek-Peyron, P.; Puissegur, M.P.; Botanch, C.; Bon, H.; Levillain, F.; Duteyrat, J.L.; Emile, J.F.; Altare, F. Langhans giant cells from M. tuberculosis-induced human granulomas cannot mediate mycobacterial uptake. J. Pathol. 2007, 211, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Herrtwich, L.; Nanda, I.; Evangelou, K.; Nikolova, T.; Horn, V.; Sagar, E.D.; Stefanowski, J.; Rogell, L.; Klein, C.; Gharun, K.; et al. DNA Damage Signaling Instructs Polyploid Macrophage Fate in Granulomas. Cell 2016, 167, 1264–1280.e18. [Google Scholar] [CrossRef]
- Helming, L.; Gordon, S. Macrophage Fusion Induced by Il-4 Alternative Activation Is a Multistage Process Involving Multiple Target Molecules. Eur. J. Immunol. 2007, 37, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Chierakul, W.; Wuthiekanun, V.; Chaowagul, W.; Amornchai, P.; Cheng, A.C.; White, N.J.; Day, N.P.; Peacock, S.J. Short report: Disease severity and outcome of melioidosis in HIV coinfected individuals. Am. J. Trop. Med. Hyg. 2005, 73, 1165–1166. [Google Scholar] [CrossRef]
- Ross, B.N.; Micheva-Viteva, S.; Hong-Geller, E.; Torres, A.G. Evaluating the role of Burkholderia pseudomallei K96243 toxins BPSS0390, BPSS0395, and BPSS1584 in persistent infection. Cell Microbiol. 2019, 21, e13096. [Google Scholar] [CrossRef] [PubMed]
- Bast, A.; Krause, K.; Schmidt, H.E.; Pudla, M.; Brakopp, S.; Hopf, V.; Breitbach, K.; Steinmetz, I. Caspase-1-dependent and -independent cell death pathways in Burkholderia pseudomallei infection of macrophages. PLoS Pathog. 2014, 10, e1003986. [Google Scholar] [CrossRef] [PubMed]
- FitzGerald, E.S.; Luz, N.F.; Jamieson, A.M. Competitive Cell Death Interactions in Pulmonary Infection: Host Modulation Versus Pathogen Manipulation. Front. Immunol. 2020, 11, 814. [Google Scholar] [CrossRef] [PubMed]
- Chow, T.K.; Eu, L.C.; Chin, K.F.; Ong, K.C.; Pailoor, J.; Vadivelu, J.; Wong, K.T. Incidental Splenic Granuloma Due to Burkholderia pseudomallei: A Case of Asymptomatic Latent Melioidosis? Am. J. Trop. Med. Hyg. 2016, 94, 522–524. [Google Scholar] [CrossRef][Green Version]
- Mezouar, S.; Diarra, I.; Roudier, J.; Desnues, B.; Mege, J.L. Tumor Necrosis Factor-Alpha Antagonist Interferes with the Formation of Granulomatous Multinucleated Giant Cells: New Insights into Mycobacterium tuberculosis Infection. Front. Immunol. 2019, 10, 1947. [Google Scholar] [CrossRef]
- Conejero, L.; Patel, N.; de Reynal, M.; Oberdorf, S.; Prior, J.; Felgner, P.L.; Titball, R.W.; Salguero, F.J.; Bancroft, G.J. Low-dose exposure of C57BL/6 mice to Burkholderia pseudomallei mimics chronic human melioidosis. Am. J. Pathol. 2011, 179, 270–280. [Google Scholar] [CrossRef]
- Pampaloni, F.; Reynaud, E.G.; Stelzer, E.H.K. The Third Dimension Bridges the Gap between Cell Culture and Live Tissue. Nat. Rev. Mol. Cell Biol. 2007, 8, 839–845. [Google Scholar] [CrossRef]
- Duval, K.; Grover, H.; Han, L.H.; Mou, Y.; Pegoraro, A.F.; Fredberg, J.; Chen, Z. Modeling Physiological Events in 2d vs. 3d Cell Culture. Physiology (Bethesda) 2017, 32, 266–277. [Google Scholar] [CrossRef]
- Elkington, P.; Lerm, M.; Kapoor, N.; Mahon, R.; Pienaar, E.; Huh, D.; Kaushal, D.; Schlesinger, L.S. In Vitro Granuloma Models of Tuberculosis: Potential and Challenges. J. Infect. Dis. 2019, 219, 1858–1866. [Google Scholar] [CrossRef]
- Dedhia, P.H.; Bertaux-Skeirik, N.; Zavros, Y.; Spence, J.R. Organoid Models of Human Gastrointestinal Development and Disease. Gastroenterology 2016, 150, 1098–1112. [Google Scholar] [CrossRef]
- Qian, X.; Song, H.; Ming, G.L. Brain Organoids: Advances, Applications and Challenges. Development 2019, 146, dev166074. [Google Scholar] [CrossRef] [PubMed]
- Akbari, S.; Arslan, N.; Senturk, S.; Erdal, E. Next-Generation Liver Medicine Using Organoid Models. Front. Cell Dev. Biol. 2019, 7, 345. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.C.; Singh, D.; Kwan, G.; Deuble, M.; Aquilina, C.; Korah, I.; Norton, R. Neuromelioidosis: Craniospinal MRI Findings in Burkholderia pseudomallei Infection. J. Neuroimaging 2016, 26, 75–82. [Google Scholar] [CrossRef]
- St John, J.A.; Walkden, H.; Nazareth, L.; Beagley, K.W.; Ulett, G.C.; Batzloff, M.R.; Beacham, I.R.; Ekberg, J.A. Burkholderia pseudomallei Rapidly Infects the Brain Stem and Spinal Cord via the Trigeminal Nerve after Intranasal Inoculation. Infect. Immun. 2016, 84, 2681–2688. [Google Scholar] [CrossRef] [PubMed]
- van Leeuwen, L.M.; van der Sar, A.M.; Bitter, W. Animal Models of Tuberculosis: Zebrafish. Cold Spring Harb. Perspect. Med. 2014, 5, a018580. [Google Scholar] [CrossRef]
- Vergunst, A.C.; Meijer, A.H.; Renshaw, S.A.; O’Callaghan, D. Burkholderia cenocepacia Creates an Intramacrophage Replication Niche in Zebrafish Embryos, Followed by Bacterial Dissemination and Establishment of Systemic Infection. Infect. Immun. 2010, 78, 1495–1508. [Google Scholar] [CrossRef] [PubMed]
- Torraca, V.; Masud, S.; Spaink, H.P.; Meijer, A.H. Macrophage-Pathogen Interactions in Infectious Diseases: New Therapeutic Insights from the Zebrafish Host Model. Dis. Model. Mech. 2014, 7, 785–797. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stockton, J.L.; Torres, A.G. Multinucleated Giant Cell Formation as a Portal to Chronic Bacterial Infections. Microorganisms 2020, 8, 1637. https://doi.org/10.3390/microorganisms8111637
Stockton JL, Torres AG. Multinucleated Giant Cell Formation as a Portal to Chronic Bacterial Infections. Microorganisms. 2020; 8(11):1637. https://doi.org/10.3390/microorganisms8111637
Chicago/Turabian StyleStockton, Jacob L., and Alfredo G. Torres. 2020. "Multinucleated Giant Cell Formation as a Portal to Chronic Bacterial Infections" Microorganisms 8, no. 11: 1637. https://doi.org/10.3390/microorganisms8111637
APA StyleStockton, J. L., & Torres, A. G. (2020). Multinucleated Giant Cell Formation as a Portal to Chronic Bacterial Infections. Microorganisms, 8(11), 1637. https://doi.org/10.3390/microorganisms8111637