Genotypic and Phenotypic Characterization of Treponema phagedenis from Bovine Digital Dermatitis

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
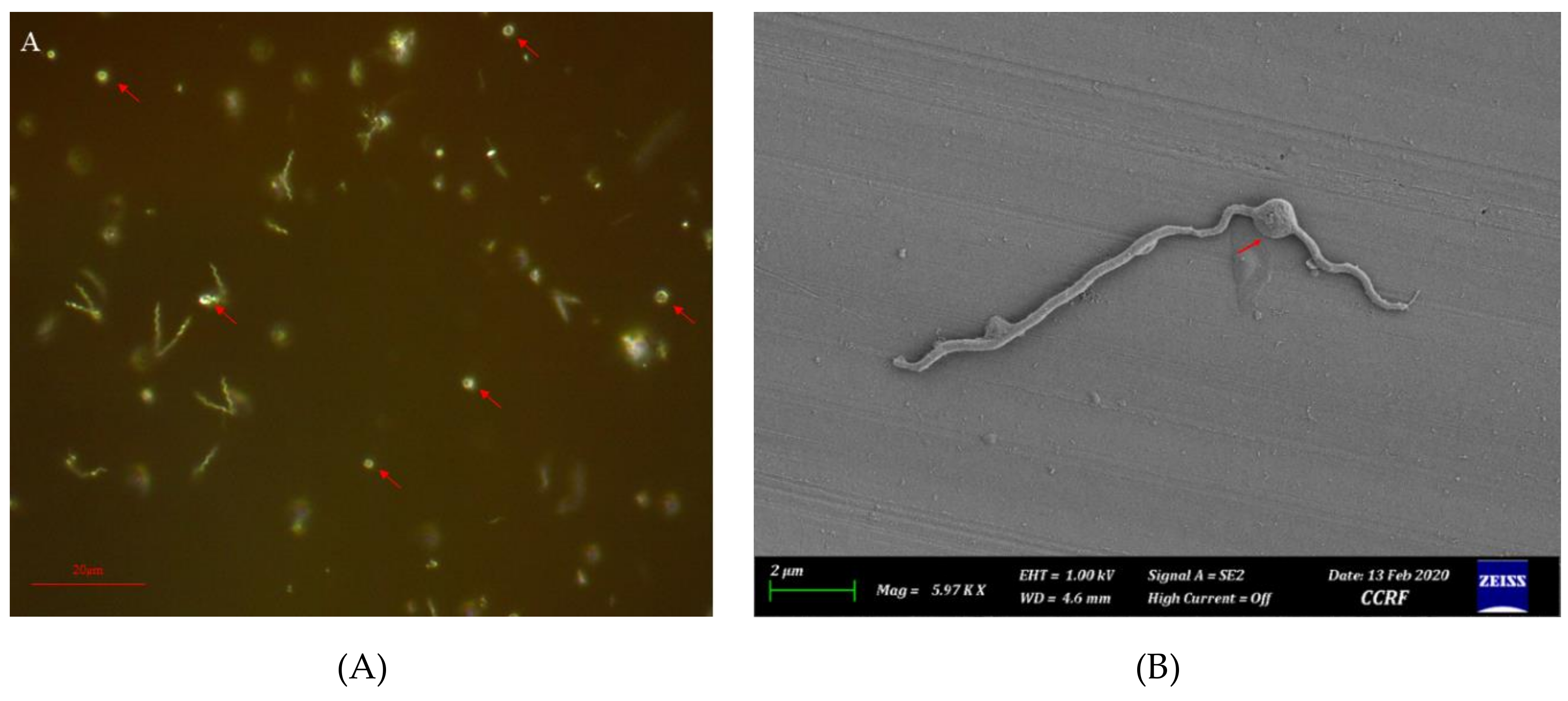
2.1. Collection, Isolation, Growth, and Microscopy
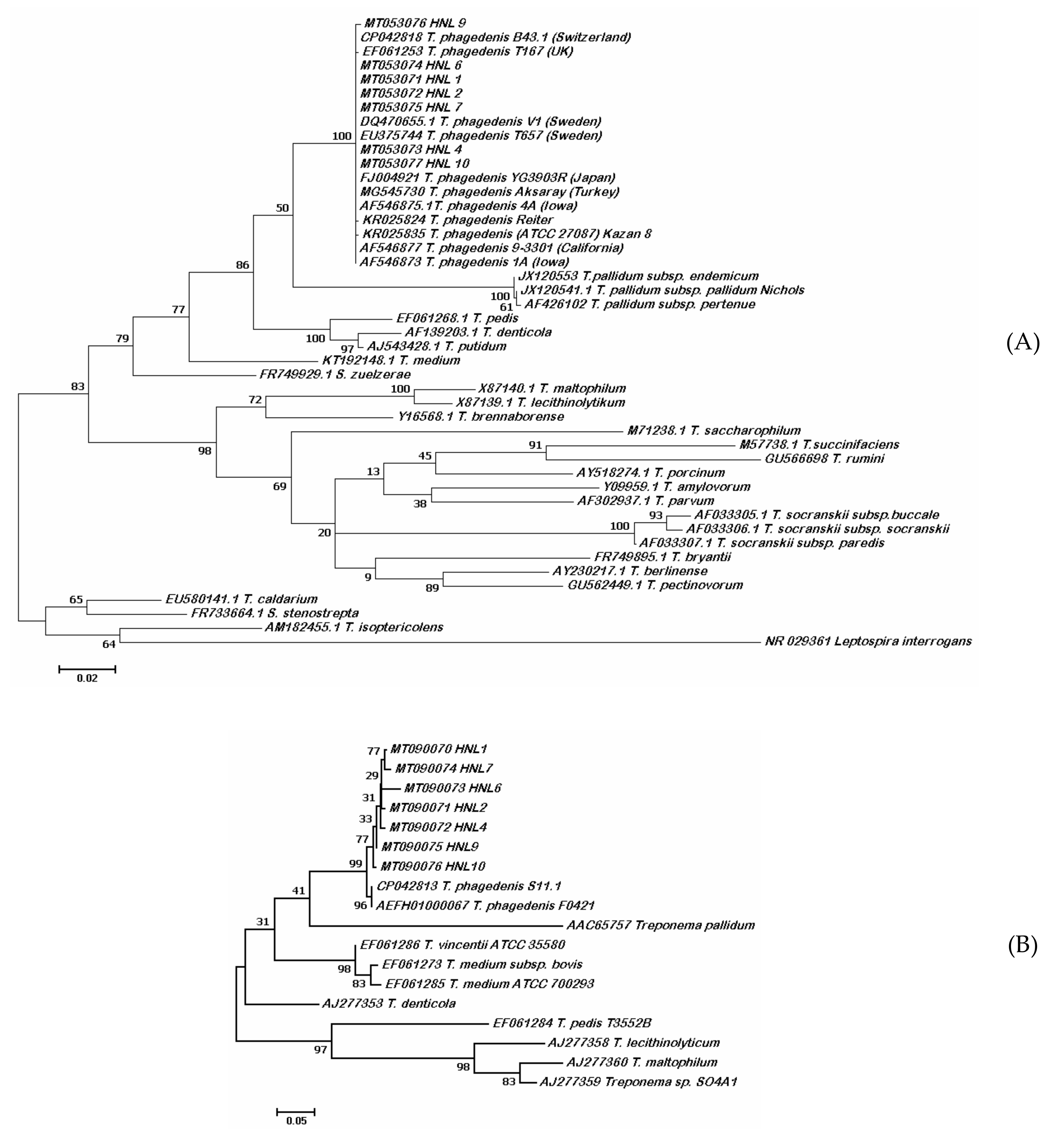
2.2. Molecular Characterization
2.3. API® ZYM, Minimum Inhibitory Concentration (MIC), and Short Chain Fatty Acids (SCFA)
3. Results
3.1. Isolation, Growth, and General Characteristics
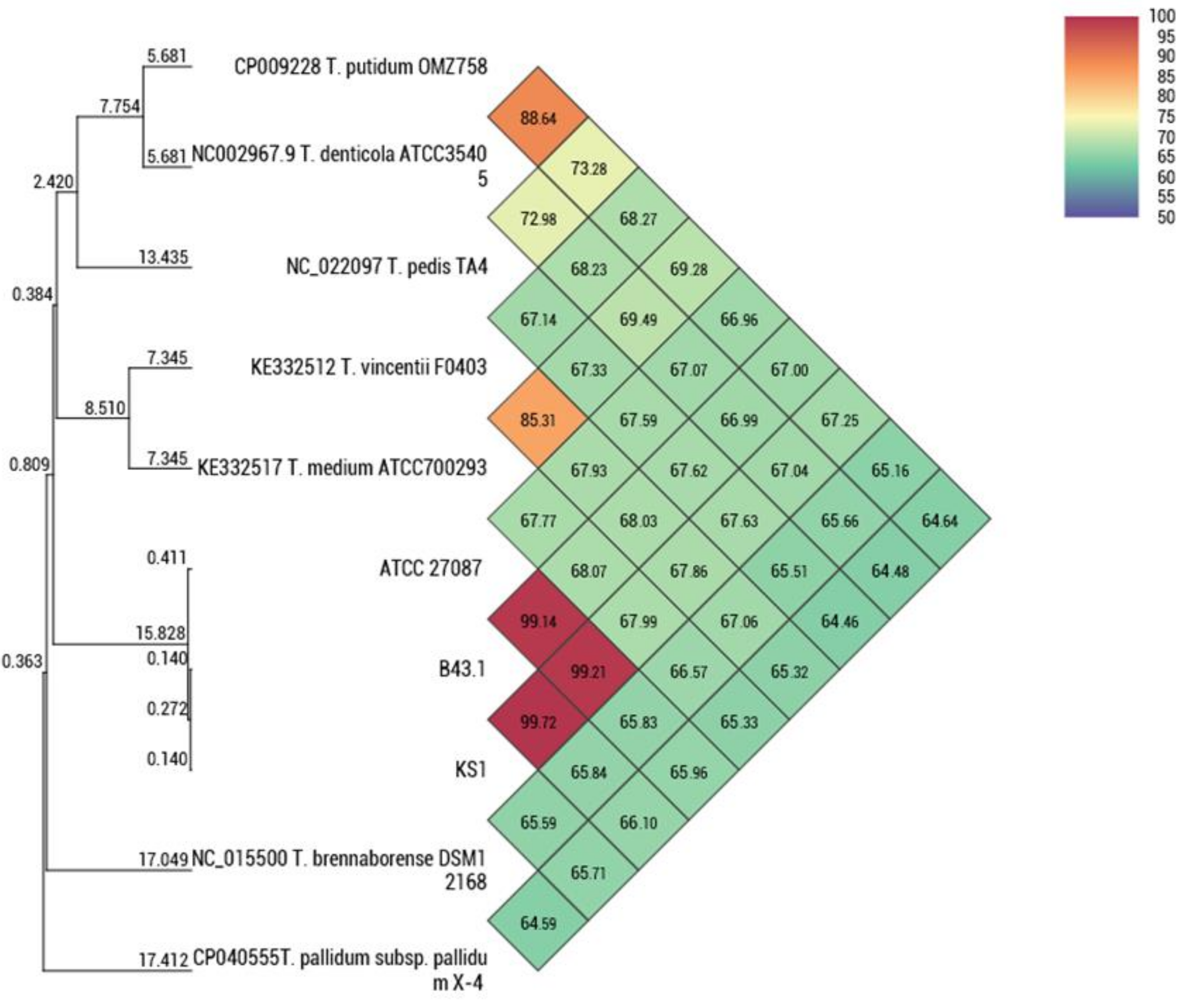
3.2. Molecular Characteristics
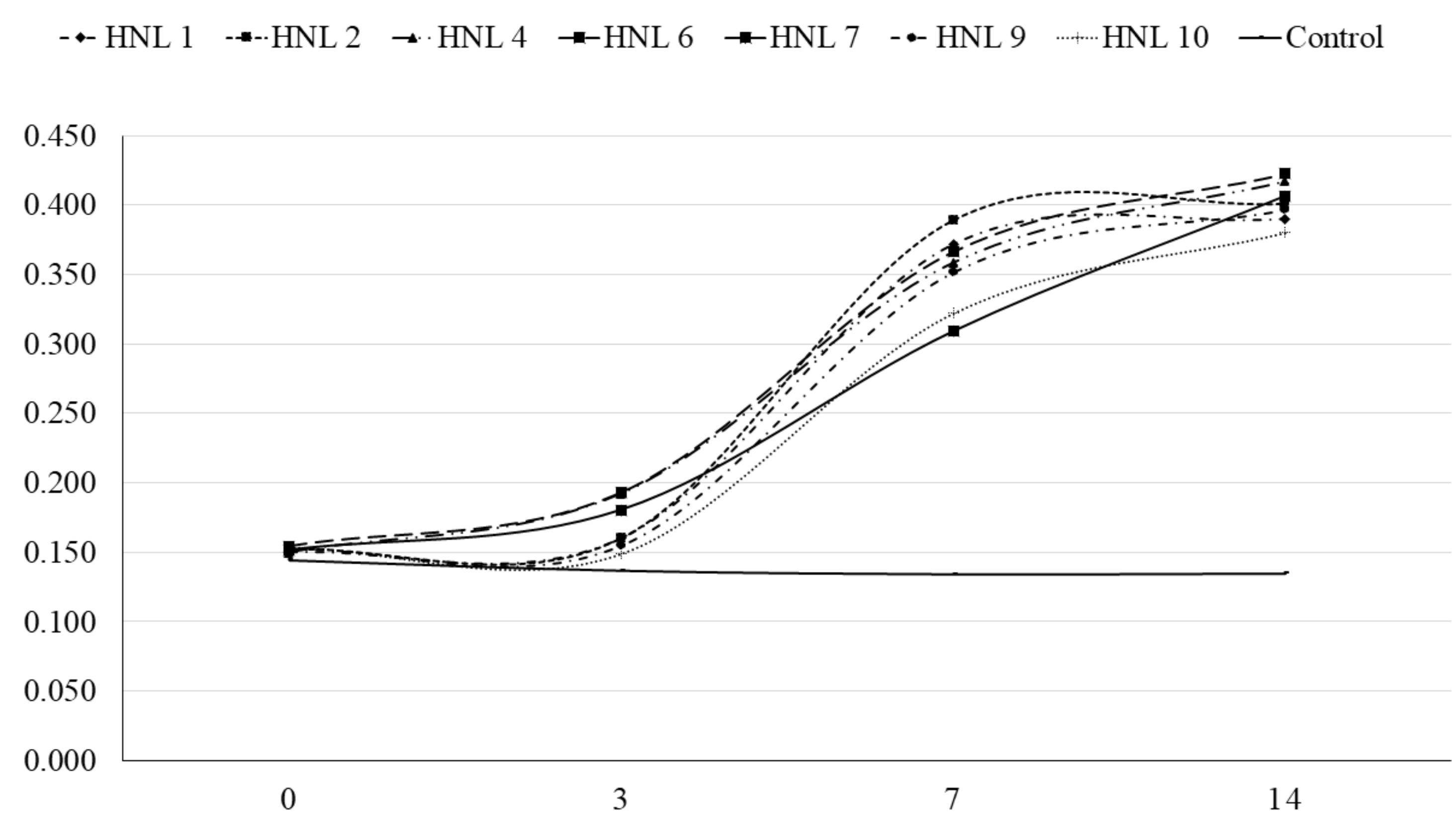
3.3. API® ZYM, Minimum Inhibitory Concentration (MIC), Short-Chain Fatty Acid (SCFA) Quantification
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Kuhnert, P.; Brodard, I.; Alsaaod, M.; Steiner, A.; Stoffel, M.H.; Jores, J. Treponema phagedenis (ex Noguchi 1912) Brumpt 1922 sp. nov., nom. rev., isolated from bovine digital dermatitis. Int. J. Syst. Evol. Microbiol. 2020, 70. [Google Scholar] [CrossRef] [PubMed]
- Yano, T.; Yamagami, R.; Misumi, K.; Kubota, C.; Kyaw, K.M.; Hayashi, T.; Yoshitani, K.; Ohtake, O.; Misawa, N. Genetic heterogeneity among strains of Treponema phagedenis-like spirochetes isolated from dairy cattle with papillomatous digital dermatitis in Japan. J. Clin. Microbiol. 2009, 47, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Demirkan, I.; Walker, R.L.; Murray, R.D.; Blowey, R.W.; Carter, S.D. Serological Evidence of Spirochaetal Infections Associated with Digital Dermatitis in Dairy Cattle. Vet. J. 1999, 157, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Evans, N.J.; Murray, R.D.; Carter, S.D. Bovine digital dermatitis: Current concepts from laboratory to farm. Vet. J. 2016, 211, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Klitgaard, K.; Nielsen, M.W.; Ingerslev, H.C.; Boye, M.; Jensen, T.K. Discovery of bovine digital dermatitis-associated Treponema spp. in the dairy herd environment by a targeted deep-sequencing approach. Appl. Environ. Microbiol. 2014, 80, 4427–4432. [Google Scholar] [CrossRef] [PubMed]
- Cheli, R.; Mortellaro, C. Digital dermatitis in cattle. In Proceedings of the 8th International Meeting on Diseases of Cattle, Milan, Italy, 9–13 September 1974; pp. 208–213. [Google Scholar]
- Moreira, T.F.; Facury Filho, E.J.; Carvalho, A.U.; Strube, M.L.; Nielsen, M.W.; Klitgaard, K.; Jensen, T.K. Pathology and bacteria related to digital dermatitis in dairy cattle in all year round grazing system in Brazil. PLoS ONE 2018, 13, 1. [Google Scholar] [CrossRef]
- Wilson-Welder, J.H.; Alt, D.P.; Nally, J.E. Digital dermatitis in cattle: Current bacterial and immunological findings. Animals 2015, 5, 1114. [Google Scholar] [CrossRef]
- Mamuad, L.L.; Joo, B.; Al, S.; Espiritu, H.M.; Jeong, S.; Kim, W.; Lee, S.; Cho, Y. Treponema spp., the dominant pathogen in the lesion of bovine digital dermatitis and its characterization in dairy cattle. Vet. Microbiol. 2020, 245, 108696. [Google Scholar] [CrossRef]
- Zinicola, M.; Lima, F.; Lima, S.; Machado, V.; Gomez, M. Altered Microbiomes in Bovine Digital Dermatitis Lesions, and the Gut as a Pathogen Reservoir. PLoS ONE 2015, 1–23. [Google Scholar] [CrossRef]
- Sullivan, L.E.; Clegg, S.R.; Angell, J.W.; Newbrook, K.; Blowey, R.W.; Carter, S.D.; Bell, J.; Duncan, J.S.; Grove-White, D.H.; Murray, R.D.; et al. High-level association of bovine digital dermatitis Treponema spp. with contagious ovine digital dermatitis lesions and presence of Fusobacterium necrophorum and Dichelobacter nodosus. J. Clin. Microbiol. 2015, 53, 1628–1638. [Google Scholar] [CrossRef]
- Nielsen, M.W.; Strube, M.L.; Isbrand, A.; Al-Medrasi, W.D.H.M.; Boye, M.; Jensen, T.K.; Klitgaard, K. Potential bacterial core species associated with digital dermatitis in cattle herds identified by molecular profiling of interdigital skin samples. Vet. Microbiol. 2016, 186, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Mushtaq, M.; Manzoor, S.; Pringle, M.; Rosander, A.; Bongcam-Rudloff, E. Draft genome sequence of “Treponema phagedenis” strain V1, isolated from bovine digital dermatitis. Stand. Genomic Sci. 2015, 10, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Demirkan, I.; Erdoğan, M.; Demirkan, A.Ç.; Bozkurt, F.; Altındiş, M.; Navruz, F.Z.; Köse, Z. Isolation and identification of Treponema pedis and Treponema phagedenis-like organisms from bovine digital dermatitis lesions found in dairy cattle in Turkey. J. Dairy Sci. 2018, 101, 10317–10326. [Google Scholar] [CrossRef] [PubMed]
- Nally, J.E.; Hornsby, R.L.; Alt, D.P.; Whitelegge, J.P. Phenotypic and proteomic characterization of treponemes associated with bovine digital dermatitis. Vet. Microbiol. 2019, 235, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Pringle, M.; Bergsten, C.; Fernström, L.L.; Höök, H.; Johansson, K.E. Isolation and characterization of Treponema phagedenis-like spirochetes from digital dermatitis lesions in Swedish dairy cattle. Acta Vet. Scand. 2008, 50, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Trott, D.J.; Moeller, M.R.; Zuerner, R.L.; Goff, J.P.; Waters, W.R.; Alt, D.P.; Walker, R.L.; Wannemuehler, M.J. Characterization of Treponema phagedenis-like spirochetes isolated from papillomatous digital dermatitis lesions in dairy cattle. J. Clin. Microbiol. 2003, 41, 2522–2529. [Google Scholar] [CrossRef] [PubMed]
- Wilson-Welder, J.H.; Elliott, M.K.; Zuerner, R.L.; Bayles, D.O.; Alt, D.P.; Stanton, T.B. Biochemical and molecular characterization of Treponema phagedenis-like spirochetes isolated from a bovine digital dermatitis lesion. BMC Microbiol. 2013, 13. [Google Scholar] [CrossRef]
- Demirkan, I.; Williams, H.F.; Dhawi, A.; Carter, S.D.; Winstanley, C.; Bruce, K.D.; Hart, C.A. Characterization of a spirochaete isolated from a case of bovine digital dermatitis. J. Appl. Microbiol. 2006, 101, 948–955. [Google Scholar] [CrossRef]
- Lee, I.; Kim, Y.O.; Park, S.C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef]
- Evans, N.J.; Brown, J.M.; Demirkan, I.; Birtles, R.; Hart, C.A.; Carter, S.D. In vitro susceptibility of bovine digital dermatitis associated spirochaetes to antimicrobial agents. Vet. Microbiol. 2008, 136, 115–120. [Google Scholar] [CrossRef][Green Version]
- Yano, T.; Moe, K.K.; Chuma, T.; Misawa, N. Antimicrobial susceptibility of Treponema phagedenis-like spirochetes isolated from dairy cattle with papillomatous digital dermatitis lesions in Japan. J. Vet. Med. Sci. 2010, 72, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Miguel, M.A.; Lee, S.S.; Mamuad, L.L.; Choi, Y.J.; Jeong, C.D.; Son, A.; Cho, K.K.; Kim, E.T.; Kim, S.B.; Lee, S.S. Enhancing Butyrate Production, Ruminal Fermentation and Microbial Population through Supplementation with Clostridium saccharobutylicum. J. Microbiol. Biotechnol. 2019, 29, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Dworkin, M.; Falkow, S.; Rosenberg, E.; Schleifer, K.-H.; Stackebrandt, E. Proteobacteria: Delta and Epsilon Subclasses, Deeply Rooting Bacteria. In The Prokaryotes, A Handbook on the Biology of Bacteria; Springer: New York, NY, USA, 2006; Volume 7, ISBN 978-0-387-25476-0. [Google Scholar]
- Matilla, M.A.; Krell, T. The effect of bacterial chemotaxis on host infection and pathogenicity. FEMS Microbiol. Rev. 2018, 42, 40–67. [Google Scholar] [CrossRef] [PubMed]
- Lux, R.; Miller, J.N.; Park, N.H.; Shi, W. Motility and chemotaxis in tissue penetration of oral epithelial cell layers by Treponema denticola. Infect. Immun. 2001, 69, 6276–6283. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.; Kennell, W.; Holt, S.C. Characterization of hemolysis and hemoxidation activities by Treponema denticola. Microb. Pathog. 1994, 16, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.; Chan, E.C.S.; Siboo, R. Iron acquisition by oral hemolytic spirochetes: Isolation of a hemin-binding protein and identification of iron reductase activity. Can. J. Microbiol. 1996, 42, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Margulis, L.; Maniotis, A.; MacAllister, J.; Scythes, J.; Brorson, O.; Hall, J.; Krumbein, W.E.; Chanman, M.J. Spirochete round bodies syphilis, lyme disease & AIDS: Resurgence of “the great imitator”? Symbiosis 2009, 47, 51–58. [Google Scholar] [CrossRef]
- Moore, L.J.; Woodward, M.J.; Grogono-Thomas, R. The occurrence of treponemes in contagious ovine digital dermatitis and the characterisation of associated Dichelobacter nodosus. Vet. Microbiol. 2005, 111, 199–209. [Google Scholar] [CrossRef]
- Demirkan, I.; Evans, N.J.; Singh, P.; Brown, J.M.; Getty, B.; Carter, S.D.; Timofte, D.; Hart, C.A.; Vink, W.D.; Birtles, R.J.; et al. Association of Unique, Isolated Treponemes with Bovine Digital Dermatitis Lesions. J. Clin. Microbiol. 2009, 47, 689–696. [Google Scholar] [CrossRef]
- Krieg, N.R.; Staley, J.T.; Brown, D.R.; Hedlund, B.P.; Paster, B.J.; Ward, N.L.; Ludwig, W.; Whitman, W.B.; Parte, A.C. The Bacteroidetes, Spirochaetes, Tenericutes (Mollicutes), Acidobacteria, Fibrobacteres, Fusobacteria, Dictyoglomi, Gemmatimonadetes, Lentisphaerae, Verrucomicrobia, Chlamydiae, and Planctomycetes. In Bergey’s Manual of Systematic Bacteriology; Springer: New York, NY, USA, 2011; Volume 4, ISBN 9780387950426. [Google Scholar]
- Van Horn, K.G.; Smibert, R.M. Fatty acid requirement of Treponema denticola and Treponema vincentii. Can. J. Microbiol. 1981, 28, 344–350. [Google Scholar] [CrossRef]
- O’Leary, W. Practical Handbook of Microbiology; CRC Press: New York, NY, USA; Washington, DC, USA, 1989; ISBN 9781466587403. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolate | Enzyme Activity | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | |
| Korean isolates * | S | W | W | - | - | - | - | - | - | S | W | - | S | W | - | - | S | - | - |
| 4a (Iowa, USA) c | S | W | W | - | - | - | - | - | - | S | W | - | S | W | - | - | S | - | - |
| V1 (Sweden) b | S | W | W | - | - | - | - | - | - | S | W | - | S | W | - | - | S | - | - |
| 1452 (Iowa, USA) c | S | W | W | - | - | - | - | - | - | S | W | - | S | S | W | - | W | - | W |
| Iowa isolates a | + | + | + | - | - | - | - | - | - | + | + | - | + | - | - | - | + | - | - |
| Japan isolates d | + | + | + | - | - | - | - | - | - | + | - | - | + | + | - | - | + | - | + |
| Turkey Group II e | + | + | + | - | + | - | - | - | - | + | + | - | + | + | - | - | + | - | - |
| B43.1 (Switzerland) f | + | + | - | - | - | - | - | - | - | + | + | - | + | - | - | - | + | - | - |
| Kazan a | + | + | + | - | + | - | - | - | - | + | + | - | + | - | - | - | + | - | - |
| Isolate | MIC (µg/mL) | |||||
|---|---|---|---|---|---|---|
| Ampicillin | Enrofloxacin | Erythromycin | Kanamycin | Rifampicin | Tetracycline | |
| HNL 1 | <0.5 | 128 | 1 | 128 | 128 | 2 |
| HNL 2 | <0.5 | 128 | 1 | 128 | 128 | 2 |
| HNL 4 | <0.5 | 128 | 1 | 128 | 128 | 2 |
| HNL 6 | <0.5 | 128 | 1 | 128 | 128 | 2 |
| HNL 7 | <0.5 | 128 | 1 | 128 | 128 | 2 |
| HNL 9 | <0.5 | 128 | 1 | 128 | 128 | 2 |
| HNL 10 | <0.5 | 128 | 1 | 128 | 128 | 2 |
| Short-chain fatty acid (mM) | Time (Day) | Isolates (HNL) | SEM | p-Value | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 4 | 6 | 7 | 9 | 10 | Control | ||||
| Formic acid | 3 | 0.00 | 0.00 x | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.000 | - |
| 7 | 4.85 b | 7.33 ab,y | 7.59 ab | 9.44 a | 8.43 ab | 8.83 ab | 6.15 ab | 0.00 c | 1.026 | <0.001 | |
| 14 | 6.98 b | 10.54 a,z | 10.34 a | 10.81 a | 8.05 ab | 7.17 b | 7.12 b | 0.00 c | 0.774 | <0.001 | |
| SEM | 1.236 | 0.463 | 0.664 | 0.439 | 0.973 | 0.367 | 0.658 | 0.000 | |||
| Acetic acid | 3 | 20.63 c,z | 22.26 c,z | 39.02 a,y | 38.55 a,y | 38.02 a,x | 26.18 b,z | 24.80 b,y | 22.4 c | 0.391 | <0.001 |
| 7 | 27.00 d,y | 35.40 c,y | 38.07 abc,y | 40.13 ab,y | 41.81 a,y | 36.06 c,y | 28.43 d,y | 21.88 e | 0.991 | <0.001 | |
| 14 | 31.73 c,x | 41.55 b,x | 50.30 a,x | 50.99 a,x | 48.34 a,x | 40.84 b,x | 40.01 b,z | 24.59 d | 0.523 | <0.001 | |
| SEM | 0.175 | 0.757 | 0.591 | 1.225 | 1.014 | 0.269 | 1.026 | 0.023 | |||
| Propionic acid | 3 | 5.35 c,x | 6.13 c,x | 11.86 a,x | 10.87 b,x | 10.46 a,x | 5.72 c,x | 5.62 c,x | 5.4 c | 0.185 | <0.001 |
| 7 | 4.27 b,y | 4.67 ab,y | 4.14 b,y | 4.59 ab,y | 4.07 b,y | 4.19 b,y | 4.40 ab,y | 5.10 a | 0.160 | 0.044 | |
| 14 | 0.00 c,z | 0.00 c,z | 0.00 c,z | 0.00 c,z | 6.59 a,z | 0.00 c,z | 0.00 c,z | 4.6 b | 0.011 | <0.001 | |
| SEM | 0.136 | 0.285 | 0.078 | 0.128 | 0.134 | 0.057 | 0.076 | 0.057 | |||
| Butyric acid | 3 | 50.51 ab | 59.37 a | 74.56 a,z | 77.97 a | 77.70 a | 49.63 ab | 49.99 ab | 18.79 b | 6.327 | 0.025 |
| 7 | 121.03 a | 121.44 a | 118.74 a,y | 120.09 a | 118.07 a | 117.74 a | 116.49 a | 15.1 8 b | 3.656 | <0.001 | |
| 14 | 117.99 a | 120.84 a | 124.31 a,x | 123.21 a | 124.04 a | 119.75 a | 119.61 a | 14.5 7 b | 4.323 | 0.002 | |
| SEM | 0.796 | 3.247 | 0.920 | 1.866 | 4.691 | 1.000 | 1.146 | 0.459 | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Espiritu, H.M.; Mamuad, L.L.; Jin, S.-j.; Kim, S.-h.; Kwon, S.-w.; Lee, S.-s.; Lee, S.-m.; Cho, Y.-i. Genotypic and Phenotypic Characterization of Treponema phagedenis from Bovine Digital Dermatitis. Microorganisms 2020, 8, 1520. https://doi.org/10.3390/microorganisms8101520
Espiritu HM, Mamuad LL, Jin S-j, Kim S-h, Kwon S-w, Lee S-s, Lee S-m, Cho Y-i. Genotypic and Phenotypic Characterization of Treponema phagedenis from Bovine Digital Dermatitis. Microorganisms. 2020; 8(10):1520. https://doi.org/10.3390/microorganisms8101520
Chicago/Turabian StyleEspiritu, Hector M., Lovelia L. Mamuad, Su-jeong Jin, Seon-ho Kim, Seok-won Kwon, Sang-suk Lee, Sang-myeong Lee, and Yong-il Cho. 2020. "Genotypic and Phenotypic Characterization of Treponema phagedenis from Bovine Digital Dermatitis" Microorganisms 8, no. 10: 1520. https://doi.org/10.3390/microorganisms8101520
APA StyleEspiritu, H. M., Mamuad, L. L., Jin, S.-j., Kim, S.-h., Kwon, S.-w., Lee, S.-s., Lee, S.-m., & Cho, Y.-i. (2020). Genotypic and Phenotypic Characterization of Treponema phagedenis from Bovine Digital Dermatitis. Microorganisms, 8(10), 1520. https://doi.org/10.3390/microorganisms8101520