Silencing of Phytopathogen Communication by the Halotolerant PGPR Staphylococcus Equorum Strain EN21

 ,
, 
Abstract
1. Introduction
2. Experimental Procedures
2.1. Bacterial Strains, Media, Compounds and Culture Conditions
2.2. Characterization of Strain EN21
2.3. Plant Growth-Promoting Assays
2.4. Quorum Quenching Activity Against Synthetic AHLs and Crude AHL Extracts From Plant Bacterial Pathogens
2.5. Competitive Assay
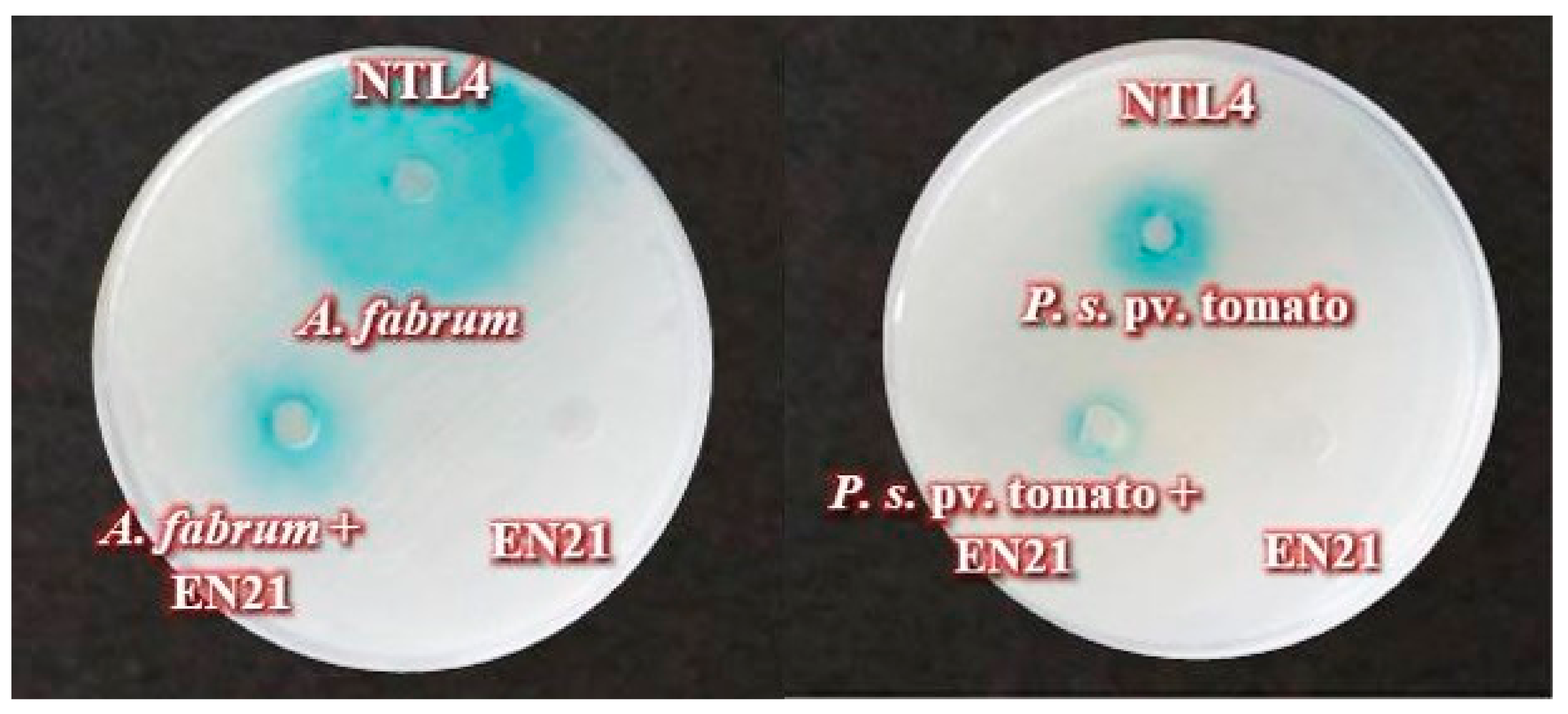
2.6. Phytopathogens QS System Interference by Co-Culture Assays
2.7. Tuber and Fruit Virulence Assays
2.8. In Vivo Arabidopsis Plant Virulence Test
2.9. In Vivo Tomato Plant Virulence Test
2.10. Statistical Analysis
3. Results
3.1. Characterization of Strain EN21
3.2. Plant Growth Promotion Capacity of Strain EN21
3.3. QQ Activity of EN21 against Synthetic AHLs and Crude AHL Extracts From Pathogenic Bacteria
3.4. Attenuation of Phytopathogenic Virulence Factors in Co-Cultures with EN21
3.5. EN21 Enhances Disease Resistance to Phytopathogens in Tubers and Fruits
3.6. EN21 Reduces P. syringae pv. Tomato DC3000 Virulence in Model Plant Arabidopsis
3.7. EN21 Reduces P. syringae pv. Tomato Virulence in Tomato Plants
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ashraf, M.Y.; Awan, A.R.; Mahmood, K. Rehabilitation of saline ecosystems through cultivation of salt tolerant plants. Pak. J. Bot. 2012, 44, 69–75. [Google Scholar]
- Almasri, M.N.; Kaluarachchi, J.J. Implications of on-ground nitrogen loading and soil transformations on ground water quality management. J. Am. Water Resour. Assoc. 2004, 40, 165–186. [Google Scholar] [CrossRef]
- Reddy, P.P. Plant Growth Promoting Rhizobacteria for Horticultural Crop Protection, 1st ed.; Springer: New Delhi, India, 2014; pp. 4–15. [Google Scholar]
- Sharma, A.; Vaishnav, A.; Jamali, H.; Srivastava, A.K.; Saxena, A.K.; Srivastava, A.K. Halophilic bacteria: Potential bioinoculants for sustainable agriculture and environment management under salt stress. In Plant-Microbe Interaction: An Approach to Sustainable Agriculture, 1st ed.; Devendra, K.C., Ajit, V., Narenda, T., Eds.; Springer: Singapore, 2016; pp. 297–325. [Google Scholar]
- Gajdács, M.; Spengler, G. The role of drug repurposing in the development of novel antimicrobial drugs: Non-antibiotic pharmacological agents as quorum sensing-inhibitors. Antibiotics 2019, 8, 270. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Nair, S.K. Quorum sensing: How bacteria can coordinate activity and synchronize their response to external signals? Protein Sci. 2012, 21, 1403–1417. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal–response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef]
- Whiteley, M.; Diggle, S.P.; Greenberg, E.P. Progress in and promise of bacterial quorum sensing research. Nature 2017, 551, 313–320. [Google Scholar] [CrossRef]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef]
- Deziel, E.; Lepine, F.; Milot, S.; He, J.; Mindrinos, M.N.; Tompkins, R.G.; Rahme, L.G. Analysis of Pseudomonas aeruginosa 4-hydroxy-2-alkylquinolines (HAQs) reveals a role for 4-hydroxy-2-heptylquinoline in cell-to-cell communication. Proc. Natl. Acad. Sci. USA 2004, 101, 1339–1344. [Google Scholar] [CrossRef]
- Barber, C.E.; Tang, J.L.; Feng, J.X.; Pan, M.Q.; Wilson, T.J.; Slater, H.; Dow, J.M.; Williams, P.; Daniels, M.J. A novel regulatory system required for pathogenicity of Xanthomonas campestris is mediated by a small diffusible signal molecule. Mol. Microbiol. 1997, 24, 555–566. [Google Scholar] [CrossRef] [PubMed]
- Quinones, B.; Dulla, G.; Lindow, S.E. Quorum sensing regulates exopolysaccharide production, motility, and virulence in Pseudomonas syringae. Mol. Plant Microbe Interact. 2005, 18, 682–693. [Google Scholar] [CrossRef] [PubMed]
- Venturi, V.; Venuti, C.; Devescovi, G.; Lucchese, C.; Friscina, A.; Degrassi, G.; Aguilar, C.; Mazzucchi, U. The plant pathogen Erwinia amylovora produces acyl-homoserine lactone signal molecules in vitro and in planta. FEMS Microbiol. Lett. 2004, 241, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Von Bodman, S.B.; Bauer, W.D.; Coplin, D.L. Quorum sensing in plant-pathogenic bacteria. Annu. Rev. Phytopathol. 2003, 41, 455–482. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.; Dessaux, Y.; Llamas, I. Saline environments as a source of potential quorum sensing disruptors to control bacterial infections: A review. Mar. Drugs 2019, 17, 191. [Google Scholar] [CrossRef] [PubMed]
- Crépin, A.; Beury-Cirou, A.; Barbey, C.; Farmer, C.; Hélias, V.; Burini, J.-F.; Faure, D.; Latour, X. N-acyl homoserine lactones in diverse Pectobacterium and Dickeya plant pathogens: Diversity, abundance, and involvement in virulence. Sensors 2012, 12, 3484–3497. [Google Scholar] [CrossRef] [PubMed]
- Dessaux, Y.; Faure, D. Quorum sensing and quorum quenching in Agrobacterium: A “Go/No Go System”? Genes 2018, 9, 210. [Google Scholar] [CrossRef]
- Fetzner, S. Quorum quenching enzymes. J. Biotechnol. 2015, 201, 2–14. [Google Scholar] [CrossRef]
- Han, Y.; Chen, F.; Li, N.; Zhu, B.; Li, X. Bacillus marcorestinctum sp. nov., a novel soil acylhomoserine lactone quorum-sensing signal quenching bacterium. Int. J. Mol. Sci. 2010, 11, 507–520. [Google Scholar] [CrossRef]
- Uroz, S.; D’Angelo-Picard, C.; Carlier, A.; Elasri, M.; Sicot, C.; Petit, A.; Oger, P.; Faure, D.; Dessaux, Y. Novel bacteria degrading N-acylhomoserine lactones and their use as quenchers of quorum-sensing-regulated functions of plant-pathogenic bacteria. Microbiology 2003, 149, 1981–1989. [Google Scholar] [CrossRef]
- Uroz, S.; Chhabra, S.R.; Cámara, M.; Williams, P.; Oger, P.; Dessaux, Y. N-Acylhomoserine lactone quorum-sensing molecules are modified and degraded by Rhodococcus erythropolis W2 by both amidolytic and novel oxidoreductase activities. Microbiology 2005, 151, 3313–3322. [Google Scholar] [CrossRef]
- Faure, D.; Dessaux, Y. Quorum sensing as a target for developing control strategies for the plant pathogen Pectobacterium. Eur. J. Plant Pathol. 2007, 119, 353–365. [Google Scholar] [CrossRef]
- Helman, Y.; Chernin, L. Silencing the mob: Disrupting quorum sensing as a means to fight plant disease. Mol. Plant Pathol. 2015, 16, 316–329. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.; Uroz, S.; Salto, R.; Fauchery, L.; Quesada, E.; Llamas, I. HqiA, a novel quorum-quenching enzyme which expands the AHL lactonase family. Sci. Rep. 2017, 7, 943. [Google Scholar] [CrossRef] [PubMed]
- Reina, J.C.; Torres, M.; Llamas, I. Stenotrophomonas maltophilia AHL-degrading strains isolated from marine invertebrate microbiota attenuate the virulence of Pectobacterium carotovorum and Vibrio coralliilyticus. Mar. Biotechnol. 2019, 21, 276–290. [Google Scholar] [CrossRef] [PubMed]
- McClean, K.H.; Winson, M.K.; Fish, L.; Taylor, A.; Chhabra, S.R.; Cámara, M.; Daykin, M.; Lamb, J.H.; Swift, S.; Bycroft, B.W.; et al. Quorum sensing and Chromobacterium violaceum: Exploitation of violacein production and inhibition for the detection of N-acylhomoserine lactones. Microbiology 1997, 143, 3703–3711. [Google Scholar] [CrossRef] [PubMed]
- Morohoshi, T.; Kato, M.; Fukamachi, K.; Kato, N.; Ikeda, T. N-acylhomoserine lactone regulates violacein production in Chromobacterium violaceum type strain ATCC 12472. FEMS Microbiol. Lett. 2008, 279, 124–130. [Google Scholar] [CrossRef]
- Chilton, M.D.; Currier, T.C.; Farrand, S.K.; Bendich, A.J.; Gordon, M.P.; Nester, E.W. Agrobacterium tumefaciens DNA and PS8 bacteriophage DNA not detected in crown gall tumors. Proc. Natl. Acad. Sci. USA 1974, 71, 3672–3676. [Google Scholar] [CrossRef]
- Moraine, R.A.; Rogovin, P. Kinetics of polysaccharide B-1459 fermentation. Biotechnol. Bioeng. 1966, 8, 511–524. [Google Scholar] [CrossRef]
- Rodríguez-Valera, F.; Ruíz-Berraquero, F.; Ramos-Cormenzana, A. Characteristics of the heterotrophic bacterial populations in hypersaline environments of different salt concentrations. Microb. Ecol. 1981, 7, 235–243. [Google Scholar] [CrossRef]
- Pikovskaya, R.I. Mobilization of phosphorus in soil connection with the vital activity of some microbial species. Microbiology 1984, 17, 362–370. [Google Scholar]
- Baird-Parker, A.C. A classification of micrococci and staphylococci based on physiological and biochemical tests. J. Gen. Microbiol. 1963, 30, 409–427. [Google Scholar] [CrossRef]
- Uttley, A.; Collins, C. Theory and practice of bacterial identification. In Cowan and Steel’s Manual for the Identification of Medical Bacteria, 3rd ed.; Barrow, G.I., Feltham, R.K.A., Eds.; Cambridge University Press: Cambridge, UK, 1993; pp. 46–49. [Google Scholar]
- Villalba, L.S.; Mikan, J.; Sánchez, J. Actividades hidrolíticas y caracterización isoenzimática de poblaciones microbianas aisladas del patrimonio documental del Archivo General de Colombia. Nova 2004, 2, 50–58. [Google Scholar] [CrossRef]
- Castro, R.; Álvarez, A.; Machado, E.; Mendoza, M.; Gómez, R.; García, P. Characterization of a extracellular chitinase produced by Serratia sp. BIOMI-363706 using colloidal chitin as substrate. Rev. Soc. Quím. Perú 2011, 77, 101–108. [Google Scholar]
- Jeffries, C.D.; Holtman, D.F.; Guse, D.G. Rapid method for determining the activity of microorganisms on nucleic acids. J. Bacteriol. 1957, 73, 590–591. [Google Scholar] [PubMed]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Ajuzieogu, C.; Odokuma, L. Toxicity assessment of produced water using Microtox Rapid Bioassay. South Asian J. Res. Microbiol. 2018, 1, 1–9. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evolut. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Abdul-Baki, A.A.; Anderson, J.D. Vigor determination in soybean seed by multiple criteria. Crop Sci. 1973, 13, 630–633. [Google Scholar] [CrossRef]
- Romero, M.; Martín-Cuadrado, A.-B.; Roca-Rivada, A.; Cabello, A.M.; Otero, A. Quorum quenching in cultivable bacteria from dense marine coastal microbial communities. FEMS Microbiol. Ecol. 2011, 75, 205–217. [Google Scholar] [CrossRef]
- Torres, M.; Rubio-Portillo, E.; Antón, J.; Ramos-Espla, A.A.; Quesada, E.; Llamas, I. Selection of the N-Acylhomoserine lactone-degrading bacterium Alteromonas stellipolaris PQQ-42 and of its potential for biocontrol in aquaculture. Front. Microbiol. 2016, 7, 646. [Google Scholar] [CrossRef]
- Marketon, M.M.; Gonzalez, J.E. Identification of two quorum-sensing systems in Sinorhizobium meliloti. J. Bacteriol. 2002, 184, 3466–3475. [Google Scholar] [CrossRef] [PubMed]
- Llamas, I.; Quesada, E.; Martínez-Canovas, M.J.; Gronquist, M.; Eberhard, A.; Gonzalez, J.E. Quorum sensing in halophilic bacteria: Detection of N-acyl-homoserine lactones in the exopolysaccharide-producing species of Halomonas. Extremophiles 2005, 9, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Frikha-Gargouri, O.; Ben Abdallah, D.; Bhar, I.; Tounsi, S. Antibiosis and bmyB gene presence as prevalent traits for the selection of efficient Bacillus biocontrol agents against crown gall disease. Front. Plant Sci. 2017, 8, 1363. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Timmermann, T.; Armijo, G.; Donoso, R.; Seguel, A.; Holuigue, L.; González, B. Paraburkholderia phytofirmans PsJN protects Arabidopsis thaliana against a virulent strain of Pseudomonas syringae through the activation of induced resistance. Mol. Plant Microbe Interact. 2017, 30, 215–230. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. In Methods in Enzymology, 1st ed.; Paker, L., Douce, R., Eds.; Academic Press: New York, NY, USA, 1987; Volume 148, pp. 350–382. [Google Scholar]
- Liang, Y.; Urano, D.; Liao, K.-L.; Hedrick, T.L.; Gao, Y.; Jones, A.M. A nondestructive method to estimate the chlorophyll content of Arabidopsis seedlings. Plant Methods 2017, 13, 26. [Google Scholar] [CrossRef]
- Yan, S.; Liu, H.; Mohr, T.J.; Jenrette, J.; Chiodini, R.; Zaccardelli, M.; Setubal, J.C.; Vinatzer, B.A. Role of recombination in the evolution of the model plant pathogen Pseudomonas syringae pv. tomato DC3000, a very atypical tomato strain. Appl. Environ. Microbiol. 2008, 74, 3171–3181. [Google Scholar] [CrossRef]
- Jeong, D.W.; Kim, H.R.; Han, S.; Jeon, C.O.; Lee, J.H. A proposal to unify two subspecies of Staphylococcus equorum: Staphylococcus equorum subsp. equorum and Staphylococcus equorum subsp. linens. Antonie Van Leeuwenhoek 2013, 104, 1049–1062. [Google Scholar] [CrossRef]
- Jha, B.; Gontia, I.; Hartmann, A. The roots of the halophyte Salicornia brachiata are a source of new halotolerant diazotrophic bacteria with plant growth-promoting potential. Plant Soil 2012, 356, 265–277. [Google Scholar] [CrossRef]
- Dutta, J.; Thakur, D. Evaluation of multifarious plant growth promoting traits, antagonistic potential and phylogenetic affiliation of rhizobacteria associated with commercial tea plants grown in Darjeeling, India. PLoS ONE 2017, 12, e0182302. [Google Scholar] [CrossRef] [PubMed]
- Mora-Ruíz, R.; Font-Verdera, F.; Díaz-Gil, C.; Urdiaín, M.; Rodríguez-Valdecantos, G.; González, B.; Orfila, A.; Rosselló-Móra, R. Moderate halophilic bacteria colonizing the phylloplane of halophytes of the subfamily Salicornioideae (Amaranthaceae). Syst. Appl. Microbiol. 2015, 38, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Mora-Ruíz, M.D.R.; Alejandre-Colomo, C.; Ledger, T.; González, B.; Orfila, A.; Rosselló-Móra, R. Non-halophilic endophytes associated with the euhalophyte Arthrocnemum macrostachyum and their plant growth promoting activity potential. FEMS Microbiol. Lett. 2018, 365, fny208. [Google Scholar] [CrossRef] [PubMed]
- Jeong, D.W.; Heo, S.; Ryu, S.; Blom, J. Genomic insights into the virulence and salt tolerance of Staphylococcus equorum. Sci. Rep. 2017, 7, 5383. [Google Scholar] [CrossRef] [PubMed]
- Mahmood, A.; Turgay, O.C.; Farooq, M.; Hayat, R. Seed biopriming with plant growth promoting rhizobacteria: A review. FEMS Microbiol. Ecol. 2016, 92, fiw112. [Google Scholar] [CrossRef] [PubMed]
- Saber, Z.; Pirdashti, H.; Esmaeili, M.; Abbasian, A.; Heidarzadeh, A. Response of wheat growth parameters to co-inoculation of plant growth promoting rhizobacteria (PGPR) and different levels of inorganic nitrogen and phosphorus. World Appl. Sci. J. 2012, 16, 213–219. [Google Scholar]
- Sharifi, R. Study of nitrogen rates effects and seed biopriming with PGPR on quantitative and qualitative yield of safflower (Carthamus tinctorius L.). Tech. J. Eng. App. Sci. 2012, 2, 162–166. [Google Scholar]
- Newman, K.L.; Chatterjee, S.; Ho, K.A.; Lindow, S.E. Virulence of plant pathogenic bacteria attenuated by degradation of fatty acid cell-to-cell signaling factors. Mol. Plant Microbe Interact. 2008, 21, 326–334. [Google Scholar] [CrossRef]
- Chu, Y.Y.; Nega, M.; Wolfle, M.; Plener, L.; Grond, S.; Jung, K.; Gotz, F. A new class of quorum quenching molecules from Staphylococcus species affects communication and growth of gram-negative bacteria. PLoS Pathog. 2013, 9, e1003654. [Google Scholar] [CrossRef]
- Molina, L.; Constantinescu, F.; Michel, L.; Reimmann, C.; Duffy, B.; Defago, G. Degradation of pathogen quorum-sensing molecules by soil bacteria: a preventive and curative biological control mechanism. FEMS Microbiol. Ecol. 2003, 45, 71–81. [Google Scholar] [CrossRef]
- Garge, S.S.; Nerurkar, A.S. Evaluation of quorum quenching Bacillus spp. for their biocontrol traits against Pectobacterium carotovorum subsp. carotovorum causing soft rot. Biocatal. Agric. Biotechnol. 2017, 9, 48–57. [Google Scholar] [CrossRef]
- Ivanov, D.A.; Bernards, M.A. Chlorophyll fluorescence imaging as a tool to monitor the progress of a root pathogen in a perennial plant. Planta 2016, 243, 263–279. [Google Scholar] [CrossRef] [PubMed]
- Cen, H.; Weng, H.; Yao, J.; He, M.; Lv, J.; Hua, S.; Li, H.; He, Y. Chlorophyll fluorescence imaging uncovers photosynthetic fingerprint of Citrus Huanglongbing. Front. Plant Sci. 2017, 8, 1509. [Google Scholar] [CrossRef]
- Qian, G.-L.; Fan, J.; Chen, D.-F.; Kang, Y.-J.; Han, B.; Hu, B.; Liu, F.-Q. Reducing Pectobacterium virulence by expression of an N-acyl homoserine lactonase gene Plpp-aiiA in Lysobacter enzymogenes strain OH11. Biol. Control 2010, 52, 17–23. [Google Scholar] [CrossRef]
- Li, Q.; Ni, H.; Meng, S.; He, Y.; Yu, Z.; Li, L. Suppressing Erwinia carotovora pathogenicity by projecting N-acyl homoserine lactonase onto the surface of Pseudomonas putida cells. J. Microbiol. Biotechnol. 2011, 21, 1330–1335. [Google Scholar] [CrossRef]
- Brazelton, J.N.; Pfeufer, E.E.; Sweat, T.A.; Gardener, B.B.; Coenen, C. 2,4-diacetylphloroglucinol alters plant root development. Mol. Plant Microbe Interact. 2008, 21, 1349–1358. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | Biopriming with EN21 | |
|---|---|---|
| Root length (cm) | 2.7 ± 1.0 | 2.8 ± 0.9 |
| Root length increase (%) | - | 3.9 |
| Shoot length (cm) | 2.3 ± 0.7 | 2.6 ± 0.8 |
| Shoot length increase (%) | - | 11.6 |
| Total length (cm) | 5.0 ± 1.6 | 5.4 ± 1.6 |
| Total length increase (%) | - | 7.5 |
| Germination rate (%) | 77.8 | 90.0 |
| Germination rate increase (%) | - | 15.7 |
| Vigour index | 390.9 | 486.1 |
| Vigour index increase (%) | - | 24.4 |
| Inoculation Treatment | Biopriming + Inoculation Treatment | |||
|---|---|---|---|---|
| Control | EN21 | Control | EN21 | |
| Root length (cm) | 11.9 ± 1.2 | 11.8 ± 1.4 | 12.3 ± 2.2 | 12.1 ± 1.7 |
| Shoot length (cm) | 18.2 ± 2.8 | 20.0 ± 2.1 | 19.3 ± 1.6 | 21.3 ± 1.6 * |
| Total length (cm) | 30.1 ± 2.6 | 31.8 ± 1.7 | 31.6 ± 2.6 | 33.4 ± 2.7 |
| Root dry weight (g) | 0.028 ± 0.013 | 0.038 ± 0.012 | 0.017 ± 0.004 | 0.027 ± 0.011 * |
| Shoot dry weight (g) | 0.121 ± 0.084 | 0.173 ± 0.051 | 0.218 ± 0.098 | 0.466 ± 0.071 * |
| Total dry weight (g) | 0.149 ± 0.090 | 0.211 ± 0.062 | 0.234 ± 0.101 | 0.493 ± 0.080 * |
| Chlorophyll a (μg mL−1) | Chlorophyll b (μg mL−1) | Total Chlorophyll (μg mL−1) | ||||
|---|---|---|---|---|---|---|
| Arnon | Lichtenthaler | Arnon | Lichtenthaler | Arnon | Lichtenthaler | |
| Control | 1.39 | 1.33 | 0.55 | 0.38 | 1.95 | 1.72 |
| EN21 | 1.24 | 1.18 | 0.53 | 0.39 | 1.77 | 1.57 |
| P. s. pv. tomato | 0.14 | 0.13 | 0.08 | 0.06 | 0.22 | 0.19 |
| P. s. pv. tomato + EN21 | 0.34 | 0.32 | 0.23 | 0.18 | 0.57 | 0.51 |
| Control | EN21 | P. s. pv. Tomato | P. s. pv. Tomato + EN21 | |
|---|---|---|---|---|
| Root dry weight (g) | 0.015 ± 0.005 | 0.022 ± 0.006 | 0.009 ± 0.003 | 0.016 ± 0.003 |
| Shoot dry weight (g) | 0.032 ± 0.007 | 0.041 ± 0.010 | 0.016 ± 0.005 * | 0.032 ± 0.006 |
| Total dry weight (g) | 0.048 ± 0.010 | 0.063 ± 0.015 | 0.024 ± 0.008 * | 0.048 ± 0.008 |
| Chlorophyll a (mg g−1) | Chlorophyll b (mg g−1) | Total Chlorophyll (mg g−1) | ||||
|---|---|---|---|---|---|---|
| Arnon | Lichtenthaler | Arnon | Lichtenthaler | Arnon | Lichtenthaler | |
| Control | 0.62 | 0.29 | 0.26 | 0.09 | 0.87 | 0.37 |
| EN21 | 0.51 | 0.24 | 0.21 | 0.08 | 0.72 | 0.32 |
| P. s. pv. tomato | 0.57 | 0.27 | 0.24 | 0.09 | 0.81 | 0.36 |
| P. s. pv. tomato + EN21 | 0.76 | 0.37 | 0.33 | 0.12 | 1.10 | 0.48 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vega, C.; Rodríguez, M.; Llamas, I.; Béjar, V.; Sampedro, I. Silencing of Phytopathogen Communication by the Halotolerant PGPR Staphylococcus Equorum Strain EN21. Microorganisms 2020, 8, 42. https://doi.org/10.3390/microorganisms8010042
Vega C, Rodríguez M, Llamas I, Béjar V, Sampedro I. Silencing of Phytopathogen Communication by the Halotolerant PGPR Staphylococcus Equorum Strain EN21. Microorganisms. 2020; 8(1):42. https://doi.org/10.3390/microorganisms8010042
Chicago/Turabian StyleVega, Clara, Miguel Rodríguez, Inmaculada Llamas, Victoria Béjar, and Inmaculada Sampedro. 2020. "Silencing of Phytopathogen Communication by the Halotolerant PGPR Staphylococcus Equorum Strain EN21" Microorganisms 8, no. 1: 42. https://doi.org/10.3390/microorganisms8010042
APA StyleVega, C., Rodríguez, M., Llamas, I., Béjar, V., & Sampedro, I. (2020). Silencing of Phytopathogen Communication by the Halotolerant PGPR Staphylococcus Equorum Strain EN21. Microorganisms, 8(1), 42. https://doi.org/10.3390/microorganisms8010042