Ciprofloxacin-Resistant Salmonella enterica Serovar Kentucky ST198 in Broiler Chicken Supply Chain and Patients, China, 2010–2016

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection Isolate Identification
2.2. Polymerase Chain Reaction (PCR) Amplification and Multilocus Sequence Typing (MLST)
2.3. Antimicrobial Susceptibility Testing
2.4. Detection of Quinolone Resistance Genes, QRDR Mutations, and SGI1 via PCR
2.5. Pulsed-Field Gel Electrophoresis
3. Results
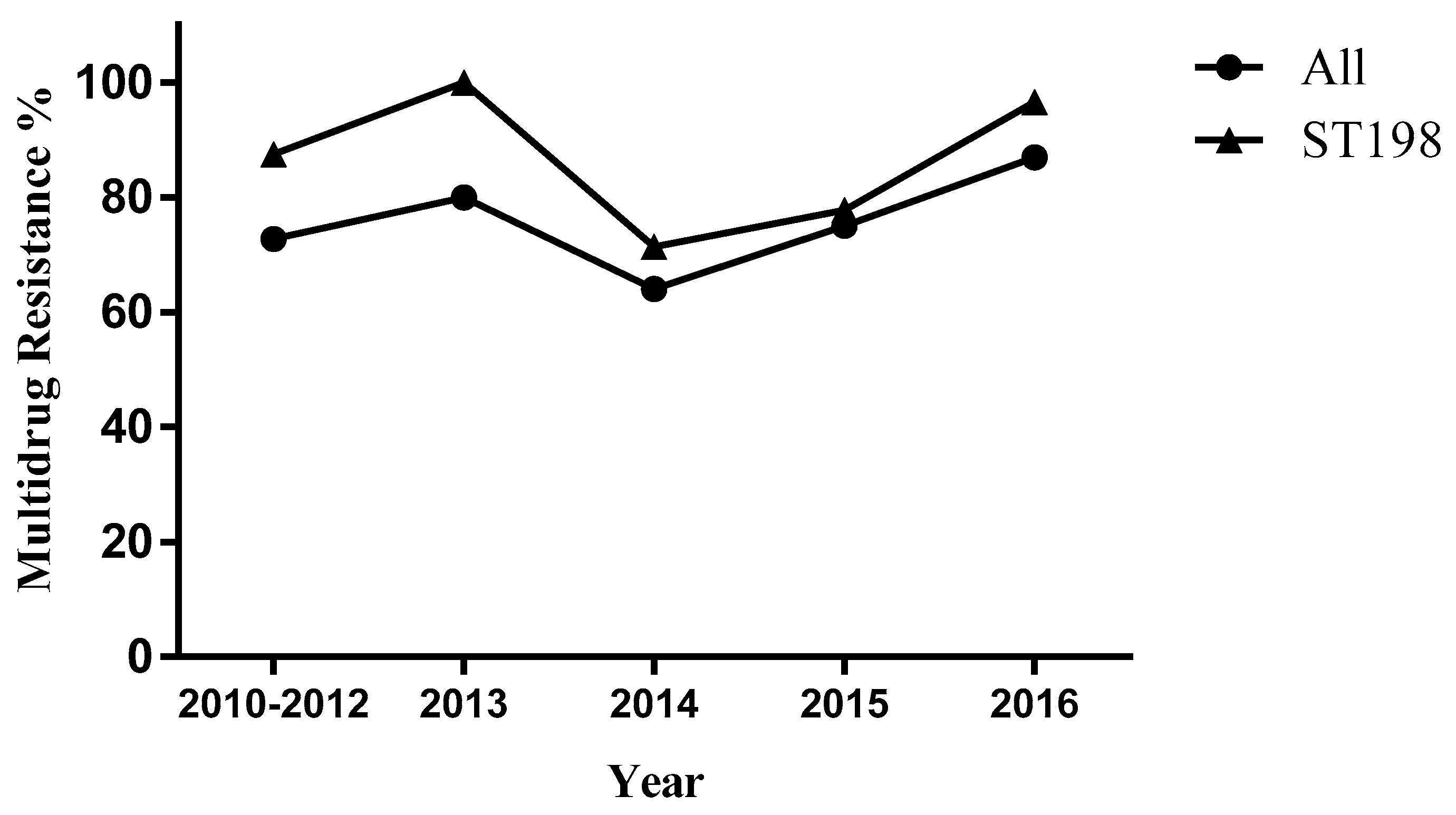
3.1. Prevalence of Salmonella enterica Kentucky
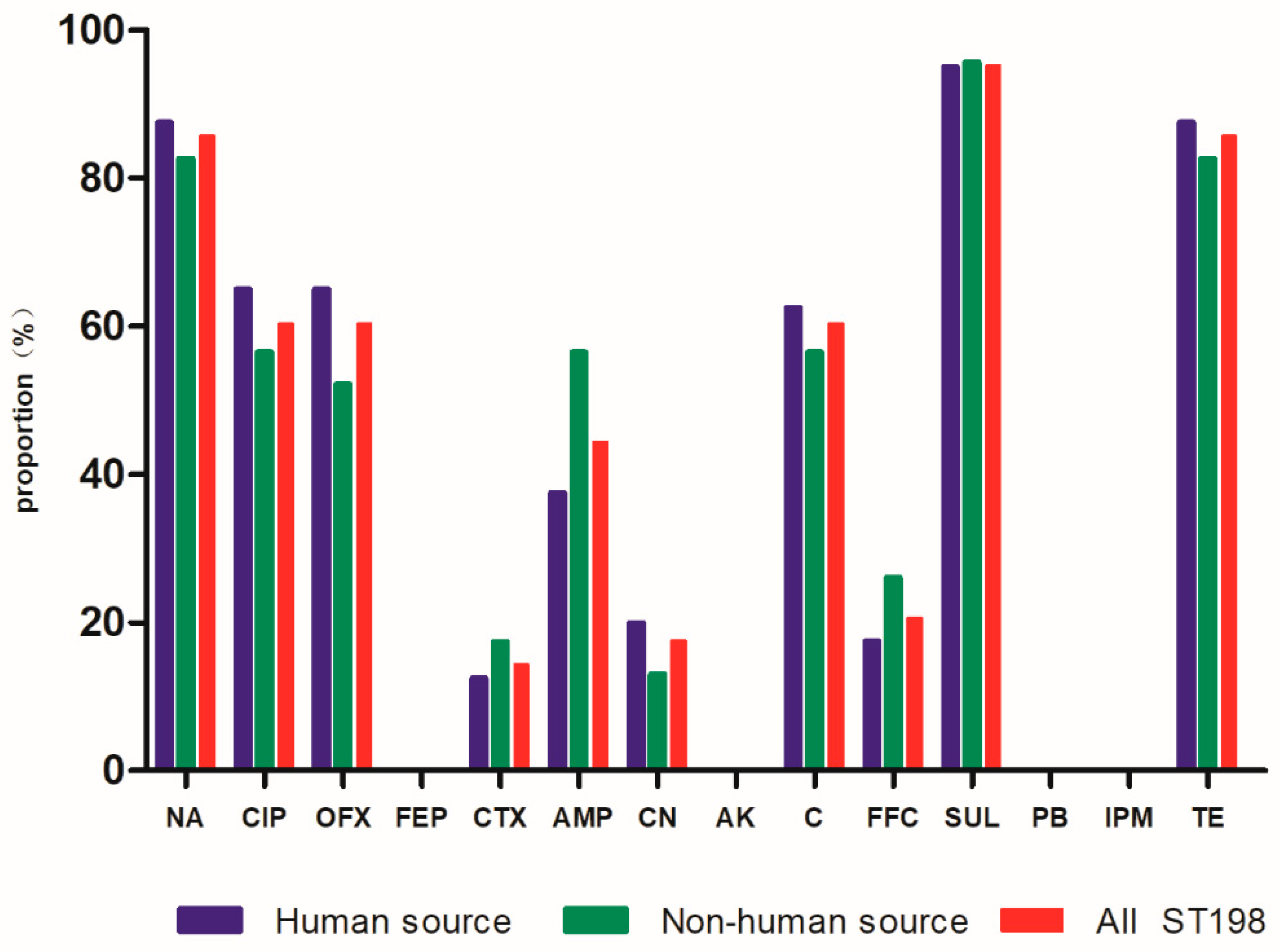
3.2. Antimicrobial Susceptibility Testing
3.3. Detection of Quinolone Resistance Genes
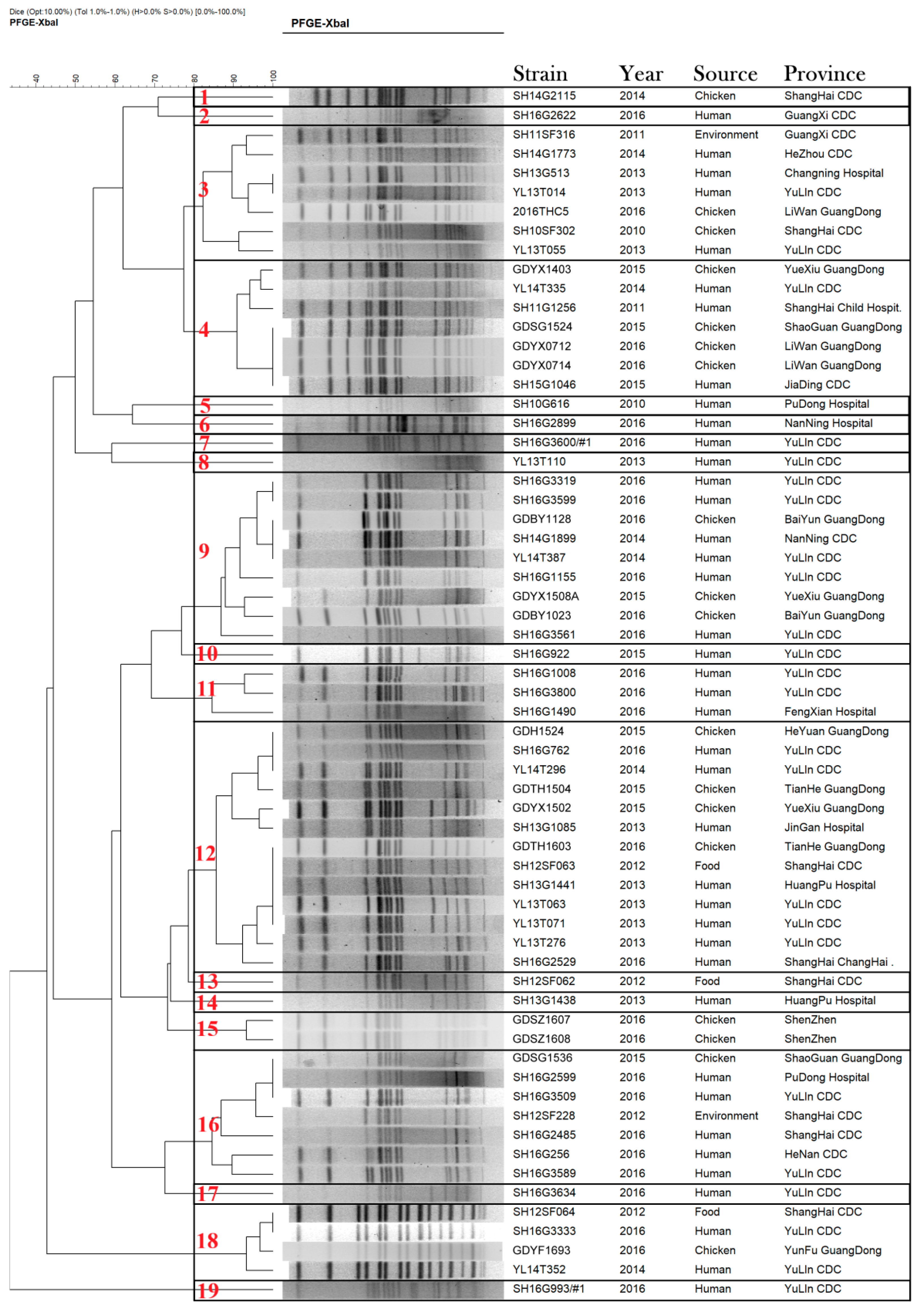
3.4. Pulsed-Field Gel Electrophoresis (PFGE)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Scallan, E.; Griffin, P.M.; Angulo, F.J.; Tauxe, R.V.; Hoekstra, R.M. Foodborne illness acquired in the United States--unspecified agents. Emerg. Infect. Dis. 2011, 17, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Guibourdenche, M.; Roggentin, P.; Mikoleit, M.; Fields, P.I.; Bockemühl, J.; Grimont, P.A.D.; Weill, F.-X. Supplement 2003–2007 (No. 47) to the White-Kauffmann-Le Minor scheme. Res. Microbiol. 2010, 161, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Chiu, L.-H.; Chiu, C.-H.; Horn, Y.-M.; Chiou, C.-S.; Lee, C.-Y.; Yeh, C.-M.; Yu, C.-Y.; Wu, C.-P.; Chang, C.-C.; Chu, C. Characterization of 13 multi-drug resistant Salmonella serovars from different broiler chickens associated with those of human isolates. BMC Microbiol. 2010, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Antimicrobial Susceptibility and Plasmid Replicon Typing of Salmonella enterica Serovar Kentucky Isolates Recovered from Broilers. Foodborne Pathog. Dis. 2016, 13, 309–315. [CrossRef]
- Zhang, L.; Fu, Y.; Xiong, Z.; Ma, Y.; Wei, Y.; Qu, X.; Zhang, H.; Zhang, J.; Liao, M. Highly Prevalent Multidrug-Resistant Salmonella from Chicken and Pork Meat at Retail Markets in Guangdong, China. Front. Microbiol. 2018, 9, 2104. [Google Scholar] [CrossRef]
- Jackson, B.R.; Griffin, P.M.; Cole, D.; Walsh, K.A.; Chai, S.J. Outbreak-associated Salmonella enterica serotypes and food Commodities, United States, 1998-2008. Emerg. Infect. Dis. 2013, 19, 1239–1244. [Google Scholar] [CrossRef]
- CRISPR Typing and Antibiotic Resistance Correlates with Polyphyletic Distribution in Human Isolates of Salmonella Kentucky. Foodborne Pathog. Dis. 2018, 15, 101–108. [CrossRef]
- Le Hello, S.; Bekhit, A.; Granier, S.A.; Barua, H.; Beutlich, J.; Zajac, M.; Munch, S.; Sintchenko, V.; Bouchrif, B.; Fashae, K.; et al. The global establishment of a highly-fluoroquinolone resistant Salmonella enterica serotype Kentucky ST198 strain. Front. Microbiol. 2013, 4, 395. [Google Scholar] [CrossRef]
- Le Hello, S.; Hendriksen, R.S.; Doublet, B.; Fisher, I.; Nielsen, E.M.; Whichard, J.M.; Bouchrif, B.; Fashae, K.; Granier, S.A.; Jourdan-Da Silva, N.; et al. International spread of an epidemic population of Salmonella enterica serotype Kentucky ST198 resistant to ciprofloxacin. J. Infect. Dis. 2011, 204, 675–684. [Google Scholar] [CrossRef]
- Westrell, T.; Monnet, D.L.; Gossner, C.; Heuer, O.; Takkinen, J. Drug-resistant Salmonella enterica serotype Kentucky in Europe. Lancet. Infect. Dis. 2014, 14, 270–271. [Google Scholar] [CrossRef]
- Ren, X.; Li, M.; Xu, C.; Cui, K.; Feng, Z.; Fu, Y.; Zhang, J.; Liao, M. Prevalence and molecular characterization of Salmonella enterica isolates throughout an integrated broiler supply chain in China. Epidemiol. Infect. 2016, 144, 2989–2999. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Qu, D.; Zhang, X.; Shen, J.; Cui, S.; Shi, Y.; Xi, M.; Sheng, M.; Zhi, S.; Meng, J. Prevalence and characterization of Salmonella serovars in retail meats of marketplace in Shaanxi, China. Int. J. Food Microbiol. 2010, 141, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Checkley, S.; Avery, B.; Chalmers, G.; Bohaychuk, V.; Gensler, G.; Reid-Smith, R.; Boerlin, P. Phenotypic and genetic characterization of antimicrobial resistance in Salmonella serovars isolated from retail meats in Alberta, Canada. Food Microbiol. 2012, 32, 110–117. [Google Scholar] [CrossRef]
- Yap, K.P.; Ho, W.S.; Gan, H.M.; Chai, L.C.; Thong, K.L. Global MLST of Salmonella Typhi Revisited in Post-genomic Era: Genetic Conservation, Population Structure, and Comparative Genomics of Rare Sequence Types. Front. Microbiol. 2016, 7, 270. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, M.; Xu, X.; Fu, Y.; Xiong, Z.; Zhang, L.; Qu, X.; Zhang, H.; Wei, Y.; Zhan, Z.; et al. High-levels of resistance to quinolone and cephalosporin antibiotics in MDR-ACSSuT Salmonella enterica serovar Enteritidis mainly isolated from patients and foods in Shanghai, China. Int. J. Food Microbiol. 2018, 286, 190–196. [Google Scholar] [CrossRef] [PubMed]
- Levings, R.S.; Partridge, S.R.; Djordjevic, S.P.; Hall, R.M. SGI1-K, a variant of the SGI1 genomic island carrying a mercury resistance region, in Salmonella enterica serovar Kentucky. Antimicrob. Agents. Chemother. 2007, 51, 317–323. [Google Scholar] [CrossRef]
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Barrett, T.J. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef]
- Yang, X.; Wu, Q.; Zhang, J.; Huang, J.; Chen, L.; Wu, S.; Zeng, H.; Wang, J.; Chen, M.; Wu, H.; et al. Prevalence, Bacterial Load, and Antimicrobial Resistance of Salmonella Serovars Isolated From Retail Meat and Meat Products in China. Front. Microbiol. 2019, 10, 2121. [Google Scholar] [CrossRef]
- Edwards, P.R. A new Salmonella type: Salmonella kentucky. J. Hyg. 1938, 38, 306–308. [Google Scholar] [CrossRef]
- Hawkey, J.; Le Hello, S.; Doublet, B.; Granier, S.A.; Hendriksen, R.S.; Fricke, W.F.; Ceyssens, P.-J.; Gomart, C.; Billman-Jacobe, H.; Holt, K.E.; et al. Global phylogenomics of multidrug-resistant Salmonella enterica serotype Kentucky ST198. Microb. Genom. 2019, 5, e000269. [Google Scholar] [CrossRef]
- Haley, B.; Kim, S.; Haendiges, J.; Keller, E.; Torpey, D.; Kim, A.; Crocker, K.; Myers, R.; Kessel, J. Salmonella enterica serovar Kentucky recovered from human clinical cases in Maryland, USA (2011–2015). Zoonoses Public Health 2019, 66, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Tay, M.; Pathirage, S.; Chandrasekaran, L.; Wickramasuriya, U.; Sadeepanie, N.; Waidyarathna, K.; Seow, K.; Hendriksen, R.; Takeuchi, M.; Schlundt, J. Whole Genome Sequencing Analysis of Nontyphoidal Salmonella enterica of Chicken Meat and Human Origin Under Surveillance in Sri Lanka. Foodborne Pathog. Dis. 2019, 16, 531–537. [Google Scholar] [CrossRef] [PubMed]
- Breurec, S.; Reynaud, Y.; Frank, T.; Farra, A.; Costilhes, G.; Weill, F.-X.; Le Hello, S. Serotype distribution and antimicrobial resistance of human Salmonella enterica in Bangui, Central African Republic, from 2004 to 2013. PLoS Negl. Trop. Dis. 2019, 13, e0007917. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Salazar, E.; Gudiño, M.E.; Sevillano, G.; Zurita, J.; Guerrero-López, R.; Jaramillo, K.; Calero-Cáceres, W. Antibiotic resistance of Salmonella strains from layer poultry farms in central Ecuador. J. Appl. Microbiol. 2019. [Google Scholar] [CrossRef]
- Li, Q.; Yin, J.; Li, Z.; Li, Z.; Du, Y.; Guo, W.; Bellefleur, M.; Wang, S.; Shi, H. Serotype distribution, antimicrobial susceptibility, antimicrobial resistance genes and virulence genes of Salmonella isolated from a pig slaughterhouse in Yangzhou, China. AMB Express 2019, 9, 210. [Google Scholar] [CrossRef]
- Ramadan, H.; Gupta, S.K.; Sharma, P.; Sallam, K.I.; Hiott, L.M.; Elsayed, H.; Barrett, J.B.; Frye, J.G.; Jackson, C.R. Draft genome sequences of two ciprofloxacin-resistant Salmonella enterica subsp. enterica serotype Kentucky ST198 isolated from retail chicken carcasses in Egypt. J. Glob. Antimicrob. Resist. 2018, 14, 101–103. [Google Scholar] [CrossRef]
- Baucheron, S.; Le Hello, S.; Doublet, B.; Giraud, E.; Weill, F.X.; Cloeckaert, A. ramR mutations affecting fluoroquinolone susceptibility in epidemic multidrug-resistant Salmonella enterica serovar Kentucky ST198. Front. Microbiol. 2013, 4, 213. [Google Scholar] [CrossRef]
- Frye, J.; Jackson, C. Genetic mechanisms of antimicrobial resistance identified in Salmonella enterica, Escherichia coli, and Enteroccocus spp. isolated from U.S. food animals. Front. Microbiol. 2013, 4, 135. [Google Scholar] [CrossRef]
- Doublet, B.; Praud, K.; Bertrand, S.; Collard, J.M.; Weill, F.X.; Cloeckaert, A. Novel insertion sequence- and transposon-mediated genetic rearrangements in genomic island SGI1 of Salmonella enterica serovar Kentucky. Antimicrob. Agents Chemother. 2008, 52, 3745–3754. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fluoroquinolone Resistance Classification | Gene about Fluoroquinolones | Proportion (%) | |
|---|---|---|---|
| PMQR (plasmid-mediated quinolone resistance) | oqxAB | 4.8%(3/63) | |
| qepA | 0.0%(0/63) | ||
| aac(6′)-Ib-cr | 30.2%(19/63) | ||
| qnrS | 7.9%(5/63) | ||
| qnrD | 0.0%(0/63) | ||
| qnrC | 0.0%(0/63) | ||
| qnrB | 6.3%(4/63) | ||
| qnrA | 0.0%(0/63) | ||
| QRDR (quinolone resistant determining region) | gyrA | His78Asn | 3.2%(2/63) |
| Ser83Phe | 87.3%(55/63) | ||
| Asp87Asn | 47.6%(30/63) | ||
| Asp87Gly | 12.7%(8/63) | ||
| Asp87Tyr | 1.6%(1/63) | ||
| parC | Ser85Ile | 65.1%(41/63) | |
| Salmonella Multidrug-Resistant Genomic Island 1 (SGI1) | Detection Rate (%) |
|---|---|
| SGI1-Ks | 28.6% (18/63) |
| SGI1-Ps | 1.6% (1/63) |
| SGI1-Ps-Qs | 0.0% (0/63) |
| SGI1-Qs | 0.0% (0/63) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, Z.; Wang, S.; Huang, Y.; Gao, Y.; Shen, H.; Chen, Z.; Bai, J.; Zhan, Z.; Wen, J.; Liao, M.; et al. Ciprofloxacin-Resistant Salmonella enterica Serovar Kentucky ST198 in Broiler Chicken Supply Chain and Patients, China, 2010–2016. Microorganisms 2020, 8, 140. https://doi.org/10.3390/microorganisms8010140
Xiong Z, Wang S, Huang Y, Gao Y, Shen H, Chen Z, Bai J, Zhan Z, Wen J, Liao M, et al. Ciprofloxacin-Resistant Salmonella enterica Serovar Kentucky ST198 in Broiler Chicken Supply Chain and Patients, China, 2010–2016. Microorganisms. 2020; 8(1):140. https://doi.org/10.3390/microorganisms8010140
Chicago/Turabian StyleXiong, Zhiying, Shaojun Wang, Yumei Huang, Yuan Gao, Haiyan Shen, Zhengquan Chen, Jie Bai, Zeqiang Zhan, Junping Wen, Ming Liao, and et al. 2020. "Ciprofloxacin-Resistant Salmonella enterica Serovar Kentucky ST198 in Broiler Chicken Supply Chain and Patients, China, 2010–2016" Microorganisms 8, no. 1: 140. https://doi.org/10.3390/microorganisms8010140
APA StyleXiong, Z., Wang, S., Huang, Y., Gao, Y., Shen, H., Chen, Z., Bai, J., Zhan, Z., Wen, J., Liao, M., & Zhang, J. (2020). Ciprofloxacin-Resistant Salmonella enterica Serovar Kentucky ST198 in Broiler Chicken Supply Chain and Patients, China, 2010–2016. Microorganisms, 8(1), 140. https://doi.org/10.3390/microorganisms8010140