1. Introduction
Commercial poultry is one of the most efficient sources of animal-protein available today [
1]. With a growing worldwide human population and a shift within the population to a diet higher in animal protein, the production of safe and efficiently produced food-animals has never been more important [
2]. At the same time, there has been an increase in the restrictions placed on available disease treatments for chicken and the management of chicken production [
3]. The federal government has begun enforcing the Food and Drug Administration Veterinary Feed Directive in 2017 that is significantly reducing the availability and usage of antibiotics as growth promoters in farm animals [
4]. Antibiotics are useful tools that prevent/treat disease, improve feed conversion, improve animal health/welfare and reduce carbon footprint [
5]. Viable alternatives to antibiotics are needed. As a result, finding alternatives to previous growth promoting and health enhancing practices is a major research enterprise. The challenge for poultry producers is to feed the world while following the regulatory mandates of the local jurisdictions where the birds are raised. One promising avenue for the enhancement of bird growth and health are “biotic” feed additives, probiotics, prebiotics and postbiotics [
6]. Probiotics, more commonly referred to in animal production as direct fed microbials, are live bacterial cultures that take up residence in the animal gut and, ideally, provide a beneficial function. Prebiotics are feed components not digestible by the host and are specifically selected to foster the growth of beneficial gut bacteria. Finally, postbiotics are fed products that are generally produced by beneficial gut microbes and have a positive impact on host health.
In the intestine, the number of genomes and biochemical reactions within the microbiota are greater than those of the host and significantly impact many aspects of host health, metabolism, immunity, development and behavior, while microbial imbalance, referred to as dysbiosis, is associated with disease [
7]. A critical and well understood function of the gut microbiota is the metabolism of host-indigestible feed ingredients. This ability of the microbiome significantly enhances the energy utilization potential of feed by the host. The microbiota and the metabolites they produce also have a profound effect on the signal transduction of the host and are often regulators of host response [
8,
9]. Postbiotics are substances produced by these metabolic processes in bacteria. They are effectively probiotic metabolic byproducts and non-viable bacterial products. The various metabolic byproducts produced as postbiotics include: Short chain fatty acids, bacteriocins, functional peptides and proteins [
6,
10]. The immune system–microbiota cross talk is necessary for the proper development and functioning of immunity. Recent studies have uncovered a major role for microbial metabolites in the regulation of the immune system [
9].
A major challenge when studying feed additives is determining the mode of action and efficacy. This is especially true for those products that may enhance growth under certain production conditions or that impact the immune system upon challenge or stress. Often, statistically significant data are difficult to generate in studies performed in well-controlled conditions such as a university research farm. These studies do not mimic the wide variety of conditions and challenges of a normal production situation. Our approach has been to determine the molecular mechanism of action of feed interventions by studying the relevant host tissue itself from a proteomic perspective [
11]. This molecular proteomic approach has two major advantages. One advantage is that in considering the change in protein function, we do not have to make assumptions generally made in genomic or transcriptomic data analysis, namely that the gene expression changes result in true phenotypic alterations. The second advantage is that an effective feed intervention will elicit a proteomic change based on its interaction with either the gut microbiome or the host tissue. Even if this change is not detectable in gross measurements, such as feed-conversion ratio or daily weight gain, it is detectible at the level of changes to the function of proteins, cells, and tissue.
The technique our group uses to measure these proteomic changes is the species-specific kinome peptide array [
12,
13]. This technology is designed for the species of interest—in this case chicken—and measures the activity of kinase enzymes within a tissue sample. Kinases are enzymes which phosphorylate protein, altering their function. Phosphorylation is the major post-translational modification carried out by host cells, and it has an impact on nearly every physiological function [
14]. The array is specifically designed to measure changes in innate and adaptive immunity, carbohydrate, protein and fatty acid metabolism, and stress responses [
15].
The suspension of antimicrobial growth promoters and ionophore coccidiostats has resulted in a reemergence of necrotic enteritis (NE), a severe
Clostridium perfringens (
C. perfringens)-induced disease that results in devastating clinical disease in chickens [
16]. Historically, antibiotics have been used to control
C. perfringens-induced necrotic enteritis in poultry. With growing concern about antibiotic resistance, poultry producers are looking for alternatives to combat this disease that is estimated to cost the world poultry industry up to
$6 billion annually [
17].
The purpose of this project was to evaluate postbiotic water additives from a proprietary pure probiotic culture of three lactic acid bacteria that may improve the health and welfare of the bird. We used coccidiosis vaccine and C. perfringens infection model as the challenge to bird health. Results from this study show that the postbiotic product resulted in detectable weight gain and reduced lesion scores compared to challenged groups. Changes in the kinome profile of the gut tissue of broiler birds, with and without pathogenic challenge, were detected. These changes were predominantly in the jejunum and centered on immune-related activities. Based on the comparison with the challenge groups, the nature of the immunological change was to modulate the immune response and dampen pathological signaling induced by the C. perfringens challenge, keeping the broiler gut in a state of homeostasis. The postbiotic appears to predominantly impact the innate immune system—the first line of defense and the branch of the immune system most quickly induced by a pathogen. The mechanism of this impact on innate immunity is centered on the phosphoinositide 3-kinase—protein kinase B (PI3K-Akt) signaling pathway and the decreased phosphorylation of key signaling intermediates. By understanding the mechanism of action, strategic feed interventions can be designed to better protect poultry health and reduce disease pathogenesis.
2. Materials and Methods
2.1. Bird Trial
The study was conducted at the Southern Plains Agricultural Research Center, Agricultural Research Service, United States Department of Agriculture and was approved by Animal Care and Use Committee at the Southern Plains Agricultural Research Center. All experiments were conducted according to guidelines established by the United States Department of Agriculture (USDA) Animal Care and Use Committee, which operates in accordance with established principles (National Research Council Guide for the Care and Use of Laboratory Animals 2011). The protocol was approved by the acting USDA Plains Area Animal Care and Use Committee that operates at the location where the experiments were carried out, Institutional Animal Care and Use Committee (IACUC) No. 2018-006.
A basal industry type broiler starter diet was prepared to meet or exceed the 1994 National Research Council’s Nutrient Requirements of Poultry. These diets were fed as crumbled pellets. Feed was kept in closed plastic/rubber containers and visually inspected every 6–9 days for pest contamination. The water was observed daily to assure the proper function of the automatic delivery system.
The study consisted of two replicates for a total of 400 broiler chicks. For a single replicate, a total of 200 broiler chicks were distributed among 8 floor pens lined with pine wood shavings (litter). A total of 8 treatments were randomly assigned to pens, and the birds were allowed free access to a commercial formulated broiler feed. Water (untreated or containing postbiotic product as appropriate) was available at all times throughout the trial. For the second replicate, two hundred and fifty fertile eggs (18-days-old) were transported from a commercial hatchery to the USDA facility for hatching. Once eggs hatched, 200 birds were moved to the rearing house on USDA facility. Group body weights were taken on days 1, 14, and 21. On day 14, all birds were orally gavaged with a 10 X dose of the coccidia vaccine (Coccivac B52, Kenilworth, NJ, USA). On days 17–19, three birds from the negative control, postbiotic negative control, Coccivac control, and Coccivac plus postbiotic were necropsied and had gut tissue samples taken, while the remaining birds were orally gavaged with 1–3 mLs of a liquid thioglycollate medium (Becton Dickinson Co., Sparks, MD, USA) broth culture of C. perfringens at 107–109 colony-forming units (cfu)/mL daily for three successive days (days 17–19). On day 21, birds were euthanized, weighed and examined for the degree of presence of necrotic enteritis lesions.
2.2. Postbiotic Product From a Proprietary Pure Probiotic Culture
Starting on day 1, postbiotic product was added to water at manufacturer’s recommendations of 1 ounce/gallon of fresh water using plastic watering jugs or 5 gallon buckets hooked up to nipple drinkers until the end of the study except for T1, T3, T4, and T5, which were given water with no metabolite products added (
Table 1). Every 4–6 days, water systems were cleaned, and fresh product water mixture was made and provided to the T2, T6, T7, and T8 treatment groups. Pure Cultures, Inc. produced a fermented product containing organic acids produced from a consortium or cocktail containing the following strains:
Pediococcus acidilactici* (Agricultural Research Service Culture Collection (NRRL)# B-67717),
Lactobacillus reuteri* (NRRL# B-67718),
Enterococcus faecium* NRRL# B-67720), and
Lactobacillus acidophilus** (NRRL# B-67701) (*proprietary strain to Pure Cultures, Inc.; **non-proprietary strain). Product is trademarked Flock Vitality
TM.
2.3. Necrotic Enteritis Intestinal Lesion Scoring and Tissue Sampling:
On day 21, birds were humanly euthanized; all euthanasia procedures followed the guidelines set down in the American Veterinary Medical Association (AVMA) Guidelines for the Euthanasia of Animals. Birds were examined for the degree of presence of necrotic enteritis lesions. All birds were lesion scored by the same person throughout the study, and 12 birds had tissue samples (liver, duodenum, jejunum and ileum) taken for further immunological/microbial testing. To quantitatively measure C. perfringens, a section of the small intestine about 15–20 cm in length, cranial to Meckel’s diverticulum, was removed. The sample was placed in 10 mL of anaerobic thioglycollate and stomached for 30 s. A 0.5 mL sample was removed from the stomached material, placed into an anaerobic vial containing 4.5 mL of anaerobic thioglycollate, and placed into an anaerobic chamber. The stomached material was incubated for 24 h at 37 °C and streaked on blood agar. The following day, the plates were examined for the presence of C. perfringens colonies. The scoring was based on a 0–4 lesion scoring, with 0 being normal and 4 being the most severe. In trial 2, on days 17–19, three birds from T1, T2, T3, and T7 were euthanized following the AVMA Guidelines for the Euthanasia of Animals. Tissue samples (liver, duodenum, jejunum and ileum) taken on days 17–19 were collected for further immunological/microbial testing. Tissue samples from the duodenum and the jejunum of five birds per experimental group were collected at day 21, flash frozen in liquid nitrogen, and sent to the University of Delaware to conduct kinome analysis using chicken-specific peptide arrays.
2.4. Chicken-Specific Immunometabolic Kinome Peptide Array
Biological replicate tissues from 5 birds per group were analyzed by a kinome peptide array. Peptide array protocol was carried out as previously described and summarized below [
18]. Briefly, 40 mg of tissue samples were used for the kinome peptide array protocol. Samples were homogenized by a Bead Ruptor homogenizer in 100 uL of a lysis buffer containing protease inhibitors. Homogenized samples were then mixed with an activation mix containing ATP and applied to peptide arrays. Arrays were incubated in a humidity chamber at 37 °C with 5% CO
2, thus allowing kinases to phosphorylate their target sites. Samples were then washed off the arrays, and a florescent phosphostain was applied. Stains not bound to phosphorylated sites were removed by a destaining process. Arrays were then imaged using a Tecan PowerScanner microarray scanner (Tecan Systems, San Jose, CA, USA) at 532–560 nm with a 580 nm filter to detect dye fluorescence.
Array images were then gridded using GenePix Pro software (Molecular Devices, San Jose, CA, USA), and the spot intensity signal was collected, thus ensuring peptide spots were correctly associated with their phosphorylation sites. Greater intensity fluorescence correlates to greater phosphorylation at the target site.
Fluorescent intensities for treatments were then compared with controls using a data normalization program—Platform for Intelligent, Integrated Kinome Analysis, version 2 (PIIKA2) [
19]. The resulting data output was then used in downstream applications such as Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) [
20] and Kyoto Encyclopedia of Genes and Genomes (KEGG) [
21] databases used to pinpoint changes in protein–protein interactions and signal transduction pathways.
4. Discussion
The goal of this study was to determine the mode of action of an in-water postbiotic product and its effects on gastrointestinal challenge in broiler chickens. Besides feeding the postbiotic itself, various challenges were tested, including C. perfringens inoculation, 10 X Coccivac inoculation, and C. perfringens plus Coccivac inoculation as a model for NE. We also considered two segments of the gut, the duodenum and the jejunum, to determine the main location of activity of the postbiotic.
Between two replicate trials compared to challenge groups, the addition of the postbiotic showed an improved weight gain, decreased
C. perfringens colony counts, decreased lesions scores and decreased mortality (
Table 2 and
Table 3). The postbiotic alone did not improve weight gain compared to control, non-challenged, birds. This may be due to the immune modulation we observed with the postbiotic, engaging certain aspects of the immune system, potentially affecting growth. However, this immune modulation also helped to maintain growth and health during an infectious challenge.
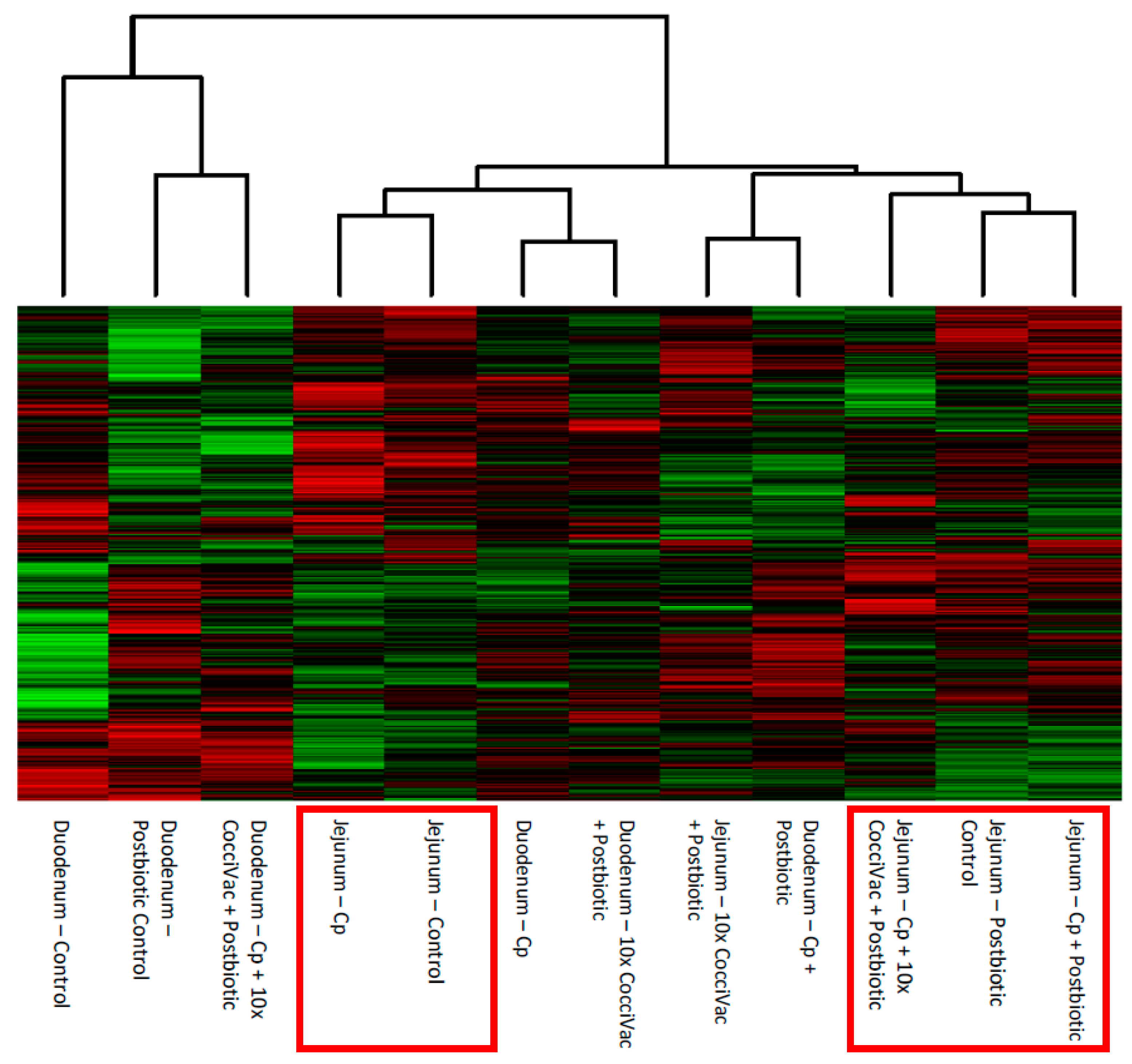
The comparison of experimental groups, including control groups separately, showed that the jejunum had the most consistent response to the postbiotic compared to the duodenum (
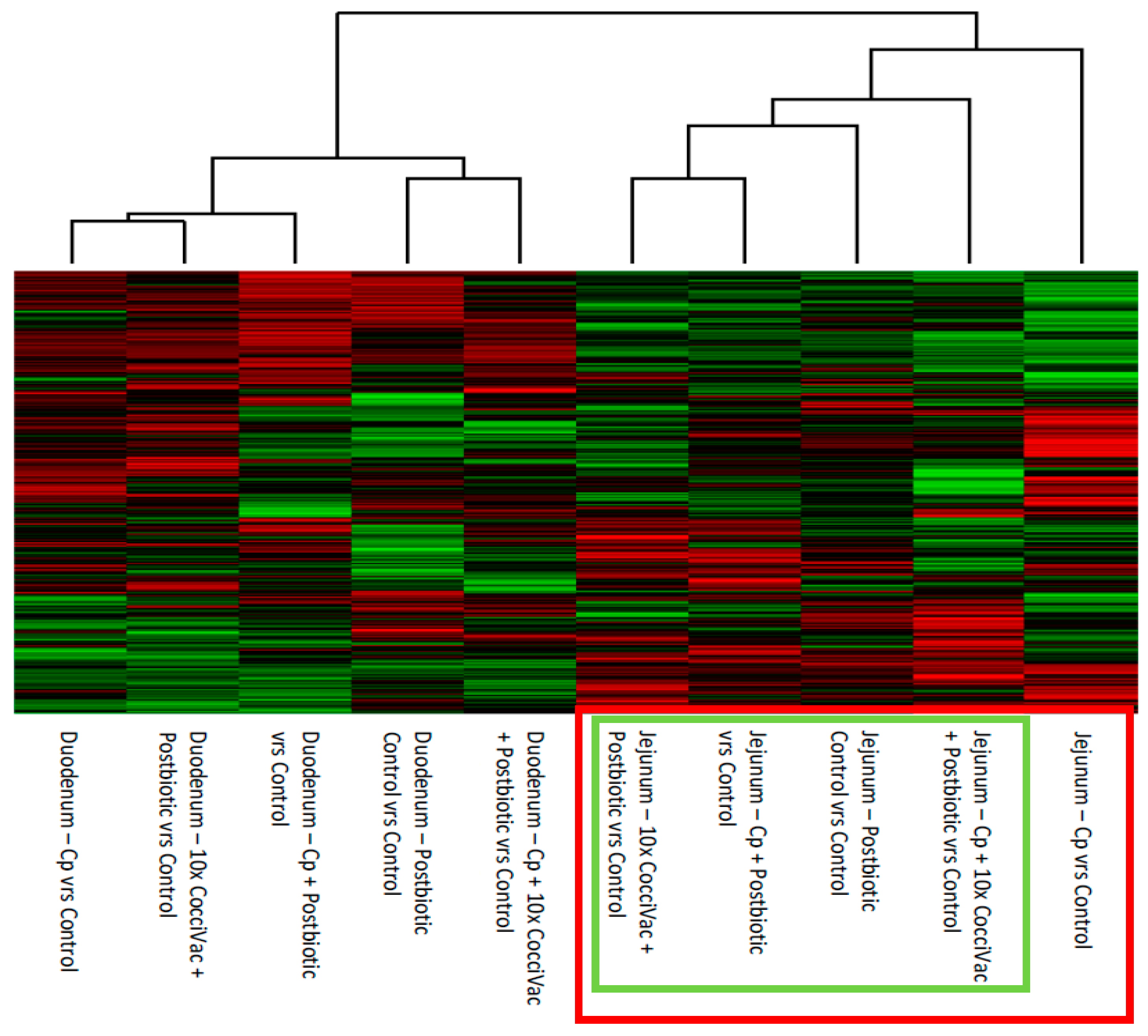
Figure 1). This jejunum response was further supported when we removed the control kinase activity signal from the data by comparing all kinome profiles to their respective non-treated, tissue matched controls (
Figure 2). The jejunum had a consistent step-wise clustering of samples with the group not treated with postbiotic clustering as the most distinct. The group without the postbiotic (jejunum inoculated with
C. perfringens) did not display responses as similar to the other groups and thus was clustered furthest out (to the right side of the figure). The duodenum, however, showed a clustering of challenge and metabolite without a clear pattern. Our interpretation is that the duodenum is less responsive to the postbiotic. Thus, the predominate kinome profile signal comes from the response to challenge.
To verify further this differential response to the postbiotic between the jejunum and the duodenum, we compared the peptides that were statistically differentially phosphorylated between postbiotic treatment,
C. perfringens challenge and the combination of postbiotic plus
C. perfringens challenge in the duodenum and the jejunum, separately (
Figure 3). These results confirmed what we had observed from the heatmap and cluster data; there was a significant amount of overlap between groups in the duodenum. Fully 20% of the differentially phosphorylated peptides were common between all three experimental groups. In addition, 28.1% were common between the
C. perfringens challenge and the
C. perfringens challenge plus postbiotic group. Combined, this represented nearly half of all differentially phosphorylated peptides in the data set. In contrast, the jejunum displayed distinct responses between each experimental group. There was relatively little overlap between the different groups in the jejunum; 61.2% of all differentially phosphorylated peptides were unique to one of the three groups. These results show that the jejunum was highly responsive to each of the experimental interventions. Based on the above results, subsequent analysis was conducted on the jejunum and three of the experimental groups, postbiotic,
C. perfringens challenge and postbiotic plus
C. perfringens challenge.
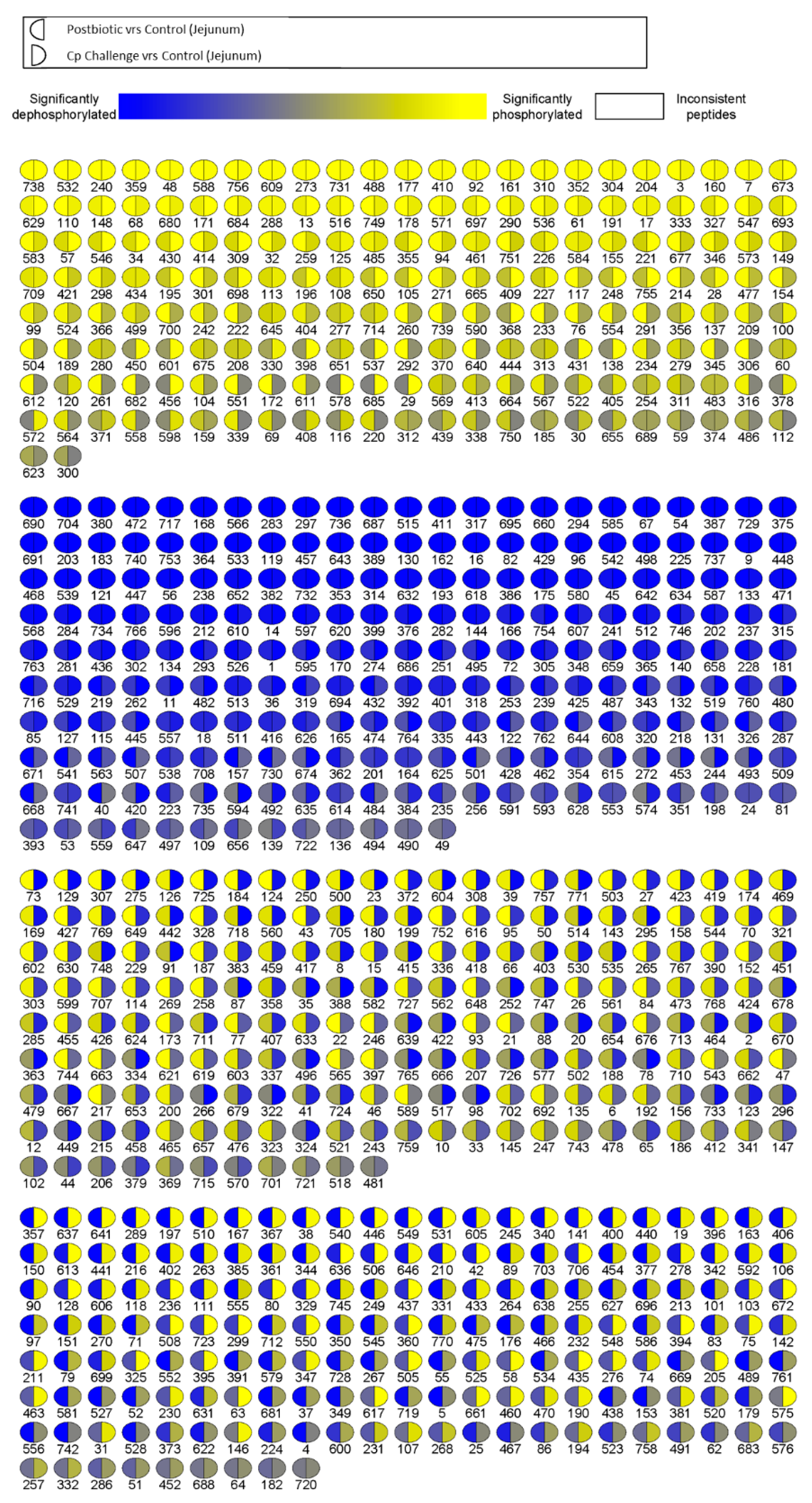
In order to visualize the differences between postbiotic-treated and
C. perfringens-challenged kinome profiles, we generated a visualization map of the peptide phosphorylation (
Figure 4). This map includes all peptides on the array, and each spot is split between postbiotic-treated (left) and
C. perfringens-challenged (right). The map shows that roughly half of the peptides were differentially phosphorylated. In fact, they were phosphorylated in opposite directions between the two groups (i.e., are yellow on one half and blue on the other half), while half were similarly phosphorylated (i.e. are all yellow or all blue). It is important to note that not all of these events are statistically significant; the colors show the significance and direction of phosphorylation (increased or decreased relative to control). The obvious difference between these two groups agrees with the heatmap, where the
C. perfringens challenge is the furthest clustered group.
Since the majority of the differentially phosphorylated peptides were unique to each of the three groups, we used these unique peptide lists to conduct biological function analysis using the online STRING protein–protein interaction database [
20]. The postbiotic peptide list generated a number of immune GO biological processes (
Table 4, bold type). This indicated that the postbiotic had an impact on immune responses within the jejunal tissue relative to control. Conducting the same analysis with the
C. perfringens challenge unique peptide list generated a larger number of immune related GO biological process top hits (
Table 5, bold type). Ten of the top 20 most significant biological processes were immune related. As
C. perfringens is a pathogenic bacterium that can elicit significant gastrointestinal pathology in broiler chickens, it is consistent with previous data that
C. perfringens would generate a significant immune response [
26,
27,
28]. When the same analysis was done considering the unique peptides from the
C. perfringens plus postbiotic group, none of the top 20 biological processes were immune response related (
Table 6). The combination of
C. perfringens and the postbiotic results in unique phosphorylation events that were not related to immunity. Given
Figure 4, where we see that many of the phosphorylation events elicited by
C. perfringens and the postbiotic were in opposite directions, the combination appears to cancel out the others responses, thus resulting in a lack of unique immune signaling. This provides intriguing evidence that, coupled with an immune inducing pathogen, the postbiotic results in a homeostatic immune modulation.
A more detailed analysis of the significant peptide phosphorylation involves incorporating them into specific signal transduction pathways. The STRING database organized the peptides into KEGG pathways and generated a significance value for each pathway.
Table 7 displays the top 20 KEGG pathways generated from the unique peptide list in the postbiotic-treated jejunum. In
Table 7, there are a number of immune pathways highlighted in bold type, several of which including the PI3K-Akt signaling pathway, the toll-like receptor signaling pathway, and the tumor necrosis factor (TNF) signaling pathway can be classified as innate immune pathways. Likewise in
Table 8 are the KEGG pathways generated from the unique peptides from the
C. perfringens challenge jejunum. Here, the immune pathways were predominately-adaptive immune pathways including the T cell receptor signaling pathway, natural killer cell mediated cytotoxicity, and the Fc epsilon receptor I (Fc epsilon RI) signaling pathway. Given that the postbiotic generated responses within innate immune pathways and
C. perfringens generated responses within adaptive immune pathways, this explains why we see GO biological processes related to immune response but with different peptide members eliciting those responses. One set was innate, and one set was adaptive. The combined postbiotic and
C. perfringens challenge group unique peptides generated substantially fewer KEGG pathways—14 total pathways. While there was some overlap in pathways from the previous KEGG lists, including insulin signaling and PI3K-AKT signaling pathways, there were also unique pathways such as the chemokine signaling pathway. Given the smaller number of pathways and the fewer peptides within those pathways, it appears that the combination muted the response of the two individual stimulating experimental treatments. Perhaps putting the birds in a more hemostatic but certainly less immunologically active state.
Finally, it was of interest that the top KEGG pathway in the postbiotic dataset was the PI3K-Akt signaling pathway, which did not show up as a top pathway in the
C. perfringens challenge data, while the insulin signaling pathway was the top pathway in the
C. perfringens challenge data and did not show up in the postbiotic dataset. We have presented the individual peptides and their phosphorylation states in these pathways for each experimental group in
Table 10 and
Table 11. It can be observed from these tables that PI3K-Akt members were predominately dephosphorylated in the postbiotic treatment group, and these corresponded to deactivating events in critical proteins including NFκB [
22], Akt [
24], PRKCA [
25], and likely TSC2 [
23] (though the function of this site is not well characterized) (
Table 10). The insulin signaling pathway as described in the KEGG database is a pathway that incorporates a variety of immune and metabolic responses, including mitogen-activated protein kinase (MAPK) signaling, glucose metabolism, apoptosis and phosphatidyl inositol signaling [
21]. When the statistically significant proteins affected by
C. perfringens (
Table 11) were put into the STRING database, their function was mapped to immune related responses including “response to cytokine” (GO biological processes) [
29] and “signaling by interleukins” (reactome pathways) [
30]. The postbiotic-affected proteins mapped to generic cellular signal transduction responses. This result indicates that
C. perfringens had an impact on innate immune responses not observed in the postbiotic treatment group.







{kind=link}
{kind=link}
{kind=link}
{kind=link}