Changes of Root Endophytic Bacterial Community Along a Chronosequence of Intensively Managed Lei Bamboo (Phyllostachys praecox) Forests in Subtropical China

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Experimental Design and Sampling
2.3. Analysis of Soil Attributes
2.4. DNA Extraction and High-Throughput Sequencing
2.5. 16S rRNA Gene Sequencing Data Processing
3. Results
3.1. Soil Attributes
3.2. Overall Taxonomic Distribution
3.3. Shifts in Bacterial Community Structure
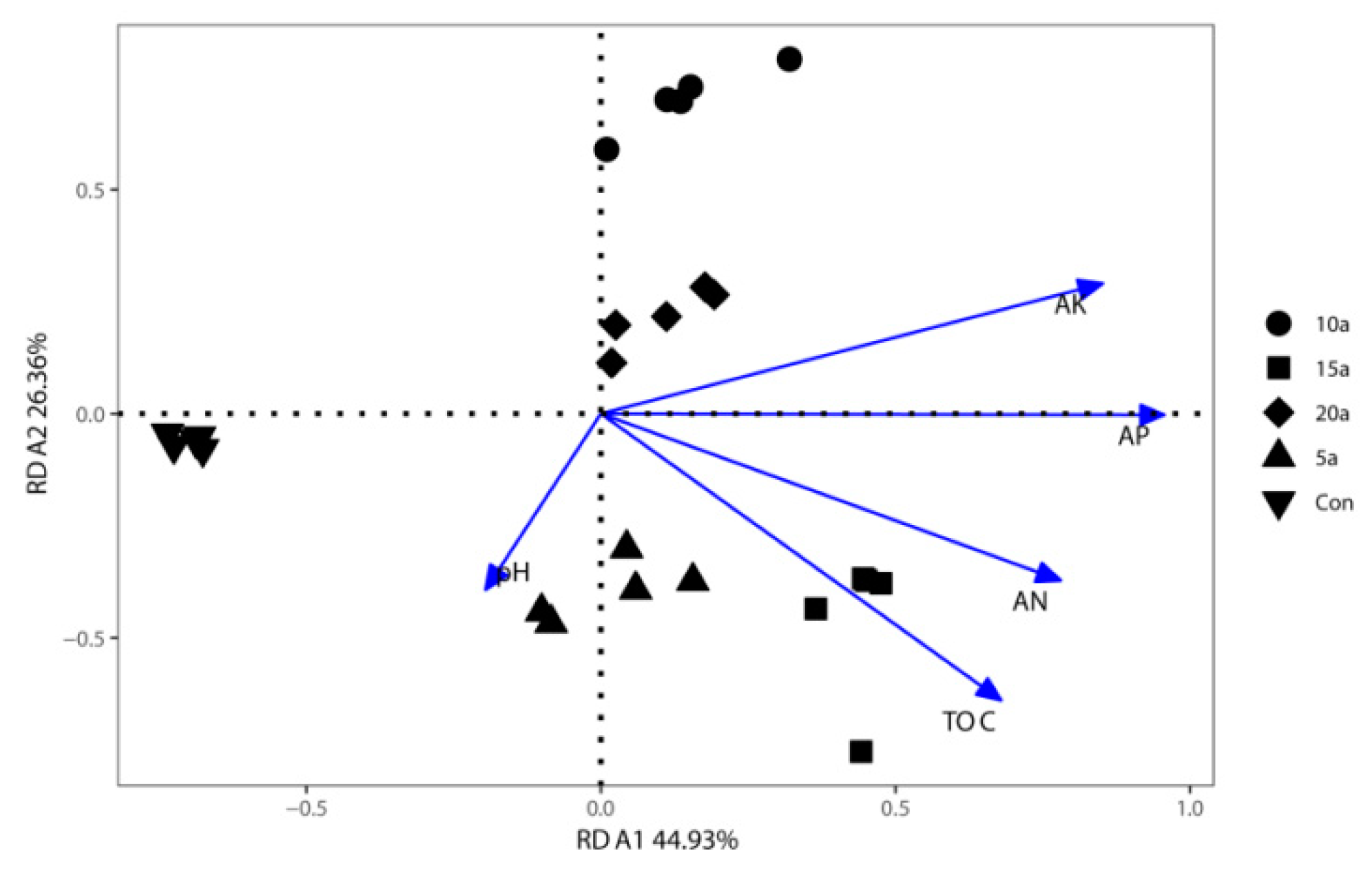
3.4. Contribution of Soil Attributes to Bacterial Community Structure
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Turner, T.R.; James, E.K.; Poole, P.S. The plant microbiome. Genome Biol. 2013, 14, 209. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Wu, Z.; Liu, T.; Wang, Y. Biodiversity, Phylogeny, and Antifungal Functions of Endophytic Fungi Associated with Zanthoxylum bungeanum. Int. J. Mol. Sci. 2016, 17, 1541. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-López, A.S.; Pintelon, I.; Stevens, V.; Imperato, V.; Timmermans, J.-P.; González-Chávez, C.; Carrillo-González, R.; Van Hamme, J.; Vangronsveld, J.; Thijs, S. Seed Endophyte Microbiome of Crotalaria pumila Unpeeled: Identification of Plant-Beneficial Methylobacteria. Int. J. Mol. Sci. 2018, 19, 291. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhong, Z.; Gai, X.; Ying, J.; Li, W.; Du, X.; Bian, F.; Yang, C. Leaf-Associated Shifts in Bacterial and Fungal Communities in Response to Chicken Rearing Under Moso Bamboo Forests in Subtropical China. Forests 2019, 10, 216. [Google Scholar] [CrossRef]
- Schulz, B. What are endophytes? In Microbial Root Endophytes; Schulz, B.J.E., Boyle, C.I.C., Sieber, T.N., Eds.; Springer: Berlin, Germany, 2006; pp. 1–13. [Google Scholar]
- Reinhold-Hurek, B.; Hurek, T. Living inside plants: Bacterial endophytes. Curr. Opin. Plant Biol. 2011, 14, 435–443. [Google Scholar] [CrossRef]
- Giauque, H.; Connor, E.W.; Hawkes, C.V. Endophyte traits relevant to stress tolerance, resource use and habitat of origin predict effects on host plants. New Phytol. 2019, 221, 2239–2249. [Google Scholar] [CrossRef]
- Li, Q.; Guo, R.; Li, Y.; Hartman, W.H.; Li, S.; Zhang, Z.; Tringe, S.G.; Wang, H. Insight into the Bacterial Endophytic Communities of Peach Cultivars Related to Crown Gall Disease Resistance. Appl. Environ. Microbiol. 2019, 85, e02931-18. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, X.; Zhang, W.; Lang, D.; Zhang, X.; Cui, G.; Zhang, X. Interactions between Endophytes and Plants: Beneficial Effect of Endophytes to Ameliorate Biotic and Abiotic Stresses in Plants. J. Plant Biol. 2019, 62, 1–13. [Google Scholar] [CrossRef]
- Baoune, H.; Ould El Hadj-Khelil, A.; Pucci, G.; Sineli, P.; Loucif, L.; Polti, M.A. Petroleum degradation by endophytic Streptomyces spp. isolated from plants grown in contaminated soil of southern Algeria. Ecotoxicol. Environ. Saf. 2018, 147, 602–609. [Google Scholar]
- Corrêa, R.C.G.; Iark, D.; de Sousa Idelfonso, A.; Uber, T.M.; Bracht, A.; Peralta, R.M. Endophytes as Pollutant-Degrading Agents: Current Trends and Perspectives. Endophytes Second. Metab. In Endophytes and Secondary Metabolites. Reference Series in Phytochemistry; Jha, S., Ed.; Springer: Cham, Switzerland, 2019; pp. 609–630. [Google Scholar]
- Li, H.-Y.; Wei, D.-Q.; Shen, M.; Zhou, Z.-P. Endophytes and their role in phytoremediation. Fungal Divers. 2012, 54, 11–18. [Google Scholar] [CrossRef]
- Singh, D.; Singh, V.K.; Singh, A.K. Endophytic Microbes: Prospects and Their Application in Abiotic Stress Management and Phytoremediation. In Seed Endophytes: Biology and Biotechnology; Verma, S.K., White, J.J.F., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 313–333. [Google Scholar]
- Suman, A.; Yadav, A.N.; Verma, P. Endophytic Microbes in Crops: Diversity and Beneficial Impact for Sustainable Agriculture. In Microbial Inoculants in Sustainable Agricultural Productivity; Research Perspectives; Singh, D.P., Singh, H.B., Prabha, R., Eds.; Springer: New Delhi, India, 2016; Volume 1, pp. 117–143. [Google Scholar]
- Kauppinen, M.; Saikkonen, K.; Helander, M.; Pirttilä, A.M.; Wäli, P.R. Epichloë grass endophytes in sustainable agriculture. Nat. Plants 2016, 2, 15224. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Verma, H.; Singh, P.P.; Singh, P.; Singh, M.; Mishra, V.; Kumar, A. Rhizome Endophytes: Roles and Applications in Sustainable Agriculture. In Seed Endophytes: Biology and Biotechnology; Verma, S.K., White, J.J.F., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 405–421. [Google Scholar]
- Robinson, R.J.; Fraaije, B.A.; Clark, I.M.; Jackson, R.W.; Hirsch, P.R.; Mauchline, T.H. Endophytic bacterial community composition in wheat (Triticum aestivum) is determined by plant tissue type, developmental stage and soil nutrient availability. Plant Soil 2016, 405, 381–396. [Google Scholar] [CrossRef]
- Estendorfer, J.; Stempfhuber, B.; Haury, P.; Vestergaard, G.; Rillig, M.C.; Joshi, J.; Schröder, P.; Schloter, M. The Influence of Land Use Intensity on the Plant-Associated Microbiome of Dactylis glomerata L. Front. Plant Sci. 2017, 8, 930. [Google Scholar] [CrossRef] [PubMed]
- Jha, P.N.; Gomaa, A.-B.; Yanni, Y.G.; El-Saadany, A.-E.Y.; Stedtfeld, T.M.; Stedtfeld, R.D.; Gantner, S.; Chai, B.; Cole, J.; Hashsham, S.A.; et al. Alterations in the Endophyte-Enriched Root-Associated Microbiome of Rice Receiving Growth-Promoting Treatments of Urea Fertilizer and Rhizobium Biofertilizer. Microb. Ecol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Bernard, S.M.; Andersen, G.L.; Chen, W. Developing microbe–plant interactions for applications in plant-growth promotion and disease control, production of useful compounds, remediation and carbon sequestration. Microb. Biotechnol. 2009, 2, 428–440. [Google Scholar] [CrossRef]
- Liu, M.; Liu, J.; Jiang, C.; Wu, M.; Song, R.; Gui, R.; Jia, J.; Li, Z. Improved nutrient status affects soil microbial biomass, respiration, and functional diversity in a Lei bamboo plantation under intensive management. J. Soils Sediments 2017, 17, 917–926. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, Z.; Bian, F.; Yang, C. Effects of composted bamboo residue amendments on soil microbial communities in an intensively managed bamboo (Phyllostachys praecox) plantation. Appl. Soil Ecol. 2019, 136, 178–183. [Google Scholar] [CrossRef]
- Fang, W.; He, J.; Lu, X.; Chen, J. Cultivation techniques of early shooting and high yielding for Lei bamboo sprout. J. Zhejing For. Coll. (China) 1994, 11, 121–128. [Google Scholar]
- Liu, Y.; Zhou, G.; Du, H.; Berninger, F.; Mao, F.; Li, X.; Chen, L.; Cui, L.; Li, Y.; Zhu, D.E.; et al. Response of carbon uptake to abiotic and biotic drivers in an intensively managed Lei bamboo forest. J. Environ. Manag. 2018, 223, 713–722. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, P.; Chang, S.X.; Wu, J.; Lin, L. Organic mulch and fertilization affect soil carbon pools and forms under intensively managed bamboo (Phyllostachys praecox) forests in southeast China. J. Soils Sediments 2010, 10, 739–747. [Google Scholar] [CrossRef]
- Zhuang, S.; Sun, X.; Liu, G.; Wong, M.; Cao, Z. Carbon Sequestration in Bamboo Plantation Soil with Heavy Winter Organic Mulching Management. Bot. Rev. 2011, 77, 252–261. [Google Scholar] [CrossRef]
- Zhai, W.; Zhong, Z.; Gao, G.; Yang, H. Influence of Mulching Management on Soil Bacterial Structure and Diversity in Phyllostachys praecox Stands. Sci. Silvae Sin. 2017, 53, 133–142. [Google Scholar]
- Chaparro, J.M.; Sheflin, A.M.; Manter, D.K.; Vivanco, J.M. Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fertil. Soils 2012, 48, 489–499. [Google Scholar] [CrossRef]
- Dubey, A.; Malla, M.A.; Khan, F.; Chowdhary, K.; Yadav, S.; Kumar, A.; Sharma, S.; Khare, P.K.; Khan, M.L. Soil microbiome: A key player for conservation of soil health under changing climate. Biodivers. Conserv. 2019, 28, 2405–2429. [Google Scholar] [CrossRef]
- Jansson, J.K.; Hofmockel, K.S. The soil microbiome—From metagenomics to metaphenomics. Curr. Opin. Microbiol. 2018, 43, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, Y.; Chang, S.X.; Liang, X.; Qin, H.; Chen, J.; Xu, Q. Linking soil fungal community structure and function to soil organic carbon chemical composition in intensively managed subtropical bamboo forests. Soil Biol. Biochem. 2017, 107, 19–31. [Google Scholar] [CrossRef]
- Hu, C.Z.; Zhang, J.M.; Hu, M.Q. Study on biological characteristics of lei bamboo. J. Zhejiang For. Coll. 1992, 9, 133–143. [Google Scholar]
- Lu, R. Soil and Agro-Chemical Analytical Methods; China Agricultural Science and Technology Press: Beijing, China, 1999; Volume 107, pp. 147–168. [Google Scholar]
- Olson, S.R. Estimation of Available Phosphorus in Soils by Extraction with Sodium Bicarbonate; US Government Printing Office: Washington, DC, USA, 1954.
- Xiao, S.; Zhang, Q.; Chen, X.; Dong, F.; Chen, H.; Liu, M.; Ali, I. Speciation Distribution of Heavy Metals in Uranium Mining Impacted Soils and Impact on Bacterial Community Revealed by High-Throughput Sequencing. Front. Microbiol. 2019, 10, 1867. [Google Scholar] [CrossRef] [PubMed]
- Tong, J.; Sun, X.; Li, S.; Qu, B.; Wan, L. Reutilization of Green Waste as Compost for Soil Improvement in the Afforested Land of the Beijing Plain. Sustainability 2018, 10, 2376. [Google Scholar] [CrossRef]
- Li, Y.; Sun, L.-L.; Sun, M.-L.; Su, H.-N.; Zhang, X.-Y.; Xie, B.-B.; Chen, X.-L.; Zhang, Y.-Z.; Qin, Q.-L. Vertical and horizontal biogeographic patterns and major factors affecting bacterial communities in the open South China Sea. Sci. Rep. 2018, 8, 8800. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.; O’Hara, R.; Simpson, G.; Solymos, P. Vegan: Community Ecology Package. R Package Version 2.5-2. 2018. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin, Germany, 2016. [Google Scholar]
- Gaston, K.J. Species Richness: Measure and Measurement; Blackwell: Cambridge, UK, 1996. [Google Scholar]
- Gomes, E.A.; Lana, U.G.; Quensen, J.F.; de Sousa, S.M.; Oliveira, C.A.; Guo, J.; Guimarães, L.J.; Tiedje, J.M. Root-associated microbiome of maize genotypes with contrasting phosphorus use efficiency. Phytobiomes 2018, 2, 129–137. [Google Scholar] [CrossRef]
- Han, J.; Xia, D.; Li, L.; Sun, L.; Yang, K.; Zhang, L. Diversity of Culturable Bacteria Isolated from Root Domains of Moso Bamboo (Phyllostachys edulis). Microb. Ecol. 2009, 58, 363–373. [Google Scholar] [CrossRef]
- Lundberg, D.S.; Lebeis, S.L.; Paredes, S.H.; Yourstone, S.; Gehring, J.; Malfatti, S.; Tremblay, J.; Engelbrektson, A.; Kunin, V.; Rio, T.G.D.; et al. Defining the core Arabidopsis thaliana root microbiome. Nature 2012, 488, 86–90. [Google Scholar] [CrossRef]
- Peng, G.; Zhang, W.; Luo, H.; Xie, H.; Lai, W.; Tan, Z. Enterobacter oryzae sp. nov., a nitrogen-fixing bacterium isolated from the wild rice species Oryza latifolia. Int. J. Syst. Evol. Microbiol. 2009, 59, 1650–1655. [Google Scholar] [CrossRef]
- Shehata, H.R.; Dumigan, C.; Watts, S.; Raizada, M.N. An endophytic microbe from an unusual volcanic swamp corn seeks and inhabits root hair cells to extract rock phosphate. Sci. Rep. 2017, 7, 13479. [Google Scholar] [CrossRef]
- Chen, S. Ecological Adaptation of Senescence Mechanism of the Major Organs of Phyllostachys Violascens to Mulching Management in; Chinese Academy of Forestry: Beijing, China, 2014. [Google Scholar]
- Gu, Y.; Ding, Y.; Ren, C.; Sun, Z.; Rodionov, D.A.; Zhang, W.; Yang, S.; Yang, C.; Jiang, W. Reconstruction of xylose utilization pathway and regulons in Firmicutes. BMC Genom. 2010, 11, 255. [Google Scholar] [CrossRef] [PubMed]
- Juretschko, S.; Loy, A.; Lehner, A.; Wagner, M. The Microbial Community Composition of a Nitrifying-Denitrifying Activated Sludge from an Industrial Sewage Treatment Plant Analyzed by the Full-Cycle rRNA Approach. Syst. Appl. Microbiol. 2002, 25, 84–99. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R. The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 1995, 41, 109–117. [Google Scholar] [CrossRef]
- Tokala, R.K.; Strap, J.L.; Jung, C.M.; Crawford, D.L.; Salove, M.H.; Deobald, L.A.; Bailey, J.F.; Morra, M.J. Novel Plant-Microbe Rhizosphere Interaction Involving Streptomyces lydicus WYEC108 and the Pea Plant (Pisum sativum). Appl. Environ. Microbiol. 2002, 68, 2161–2171. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of Plant Growth-Promoting Bacteria for Biocontrol of Plant Diseases: Principles, Mechanisms of Action, and Future Prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef]
- Zhao, J.; Zhang, R.; Xue, C.; Xun, W.; Sun, L.; Xu, Y.; Shen, Q. Pyrosequencing Reveals Contrasting Soil Bacterial Diversity and Community Structure of Two Main Winter Wheat Cropping Systems in China. Microb. Ecol. 2014, 67, 443–453. [Google Scholar] [CrossRef]
- Zhao, J.; Ni, T.; Li, Y.; Xiong, W.; Ran, W.; Shen, B.; Shen, Q.; Zhang, R. Responses of Bacterial Communities in Arable Soils in a Rice-Wheat Cropping System to Different Fertilizer Regimes and Sampling Times. PLoS ONE 2014, 9, e85301. [Google Scholar] [CrossRef]
- Cui, E.; Fan, X.; Li, Z.; Liu, Y.; Neal, A.L.; Hu, C.; Gao, F. Variations in soil and plant-microbiome composition with different quality irrigation waters and biochar supplementation. Appl. Soil Ecol. 2019, 142, 99–109. [Google Scholar] [CrossRef]
- Rodríguez-Blanco, A.; Sicardi, M.; Frioni, L. Plant genotype and nitrogen fertilization effects on abundance and diversity of diazotrophic bacteria associated with maize (Zea mays L.). Biol. Fertil. Soils 2015, 51, 391–402. [Google Scholar] [CrossRef]
- Wemheuer, F.; Wemheuer, B.; Kretzschmar, D.; Pfeiffer, B.; Herzog, S.; Daniel, R.; Vidal, S. Impact of grassland management regimes on bacterial endophyte diversity differs with grass species. Lett. Appl. Microbiol. 2016, 62, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Ren, F.; Dong, W.; Yan, D.-H. Endophytic bacterial communities of Jingbai Pear trees in north China analyzed with Illumina sequencing of 16S rDNA. Arch. Microbiol. 2019, 201, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Greissworth, E.; Mucci, C.; Williams, M.A.; De Bolt, S. Characterization of culturable bacterial endophytes of switchgrass (Panicum virgatum L.) and their capacity to influence plant growth. GCB Bioenergy 2013, 5, 674–682. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| pH | TOC (g/kg) | AN (g/kg) | AP (g/kg) | AK (mg/kg) | |
|---|---|---|---|---|---|
| Con | 4.55 ± 0.25 b | 25.66 ± 0.92 c | 159.12 ± 25.25 d | 45.81 ± 12.06 c | 114.32 ± 16.64 b |
| 5a | 5.35 ± 0.59 a | 44.51 ± 4.47 b | 409.49 ± 80.07 b | 159.90 ± 14.31 ab | 240.60 ± 100.31 a |
| 10a | 4.23 ± 0.32 b | 26.59 ± 3.18 c | 241.56 ± 58.52 c | 145.52 ± 16.62 b | 303.20 ± 87.01 a |
| 15a | 4.21 ± 0.19 b | 59.04 ± 12.80 a | 417.50 ± 42.97 ab | 163.41 ± 13.94 a | 278.99 ± 41.66 a |
| 20a | 4.56 ± 0.41 b | 54.37 ± 4.22 a | 480.82 ± 26.43 a | 154.94 ± 5.15 ab | 311.98 ± 77.16 a |
| pH | TOC | AN | AP | AK | |
|---|---|---|---|---|---|
| Proteobacteria | 0.012 | −0.266 | −0.472 * | −0.583 ** | −0.477 * |
| Actinobacteria | 0.341 | −0.118 | 0.178 | 0.295 | 0.095 |
| Firmicutes | −0.064 | 0.308 | 0.497 * | 0.507 ** | 0.544 ** |
| Bacteroidetes | −0.318 | 0.624 ** | 0.474 * | 0.613 ** | 0.439 * |
| Acidobacteria | −0.288 | −0.347 | −0.261 | −0.208 | −0.212 |
| Chloroflexi | −0.182 | 0.148 | −0.048 | −0.043 | −0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Zhong, Z.; Gai, X.; Du, X.; Bian, F.; Yang, C.; Gao, G.; Wen, X. Changes of Root Endophytic Bacterial Community Along a Chronosequence of Intensively Managed Lei Bamboo (Phyllostachys praecox) Forests in Subtropical China. Microorganisms 2019, 7, 616. https://doi.org/10.3390/microorganisms7120616
Zhang X, Zhong Z, Gai X, Du X, Bian F, Yang C, Gao G, Wen X. Changes of Root Endophytic Bacterial Community Along a Chronosequence of Intensively Managed Lei Bamboo (Phyllostachys praecox) Forests in Subtropical China. Microorganisms. 2019; 7(12):616. https://doi.org/10.3390/microorganisms7120616
Chicago/Turabian StyleZhang, Xiaoping, Zheke Zhong, Xu Gai, Xuhua Du, Fangyuan Bian, Chuanbao Yang, Guibin Gao, and Xing Wen. 2019. "Changes of Root Endophytic Bacterial Community Along a Chronosequence of Intensively Managed Lei Bamboo (Phyllostachys praecox) Forests in Subtropical China" Microorganisms 7, no. 12: 616. https://doi.org/10.3390/microorganisms7120616
APA StyleZhang, X., Zhong, Z., Gai, X., Du, X., Bian, F., Yang, C., Gao, G., & Wen, X. (2019). Changes of Root Endophytic Bacterial Community Along a Chronosequence of Intensively Managed Lei Bamboo (Phyllostachys praecox) Forests in Subtropical China. Microorganisms, 7(12), 616. https://doi.org/10.3390/microorganisms7120616