Characterization of Non-O157 STEC Infecting Bacteriophages Isolated from Cattle Faeces in North-West South Africa

 ,
,  ,
,  , and
, and
Abstract

1. Introduction
2. Materials and Methods
2.1. Bacteriophage Isolation, Host Range Determination, Propagation and Titration
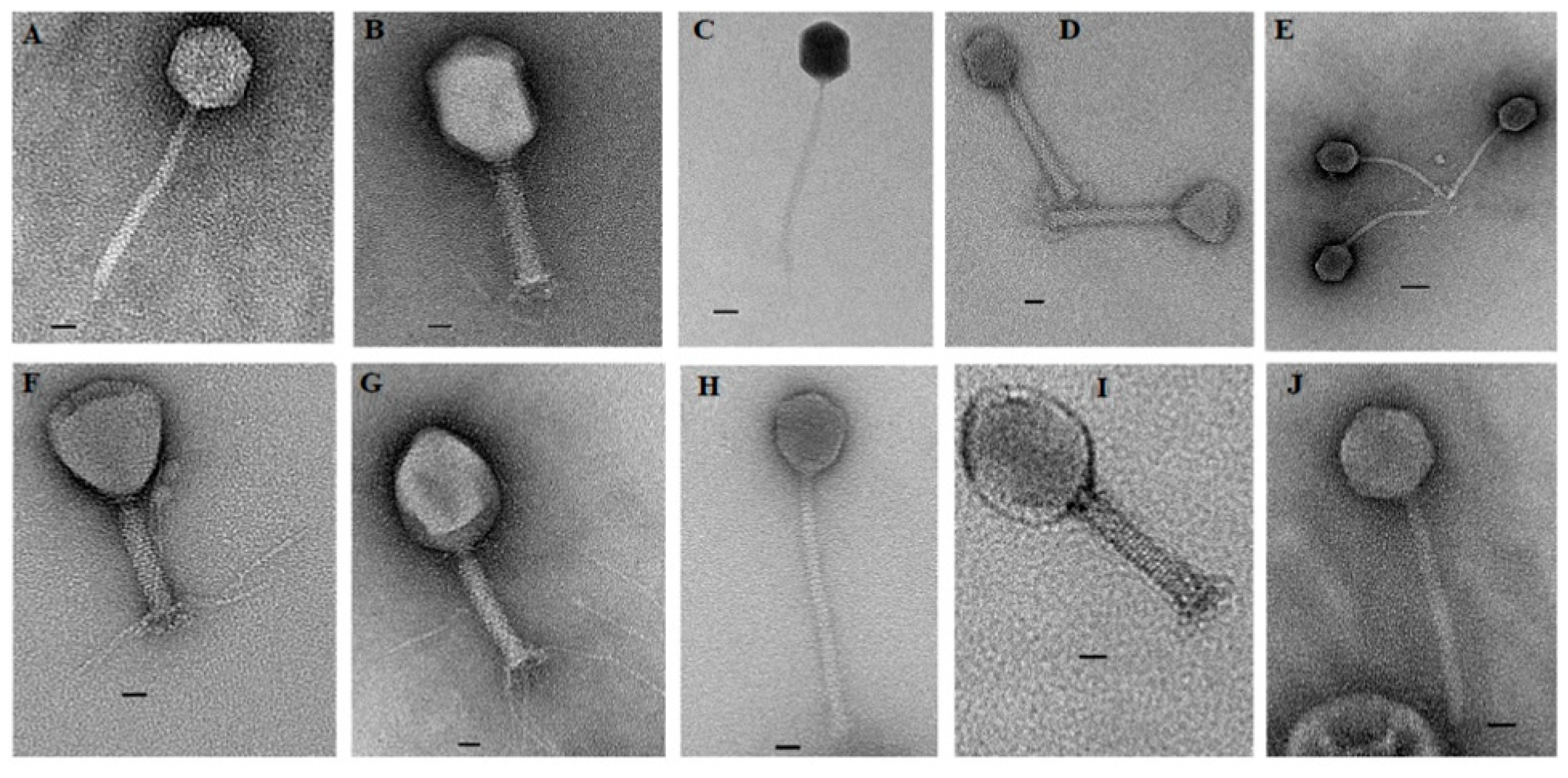
2.2. Transmission Electron Microscopy (TEM)
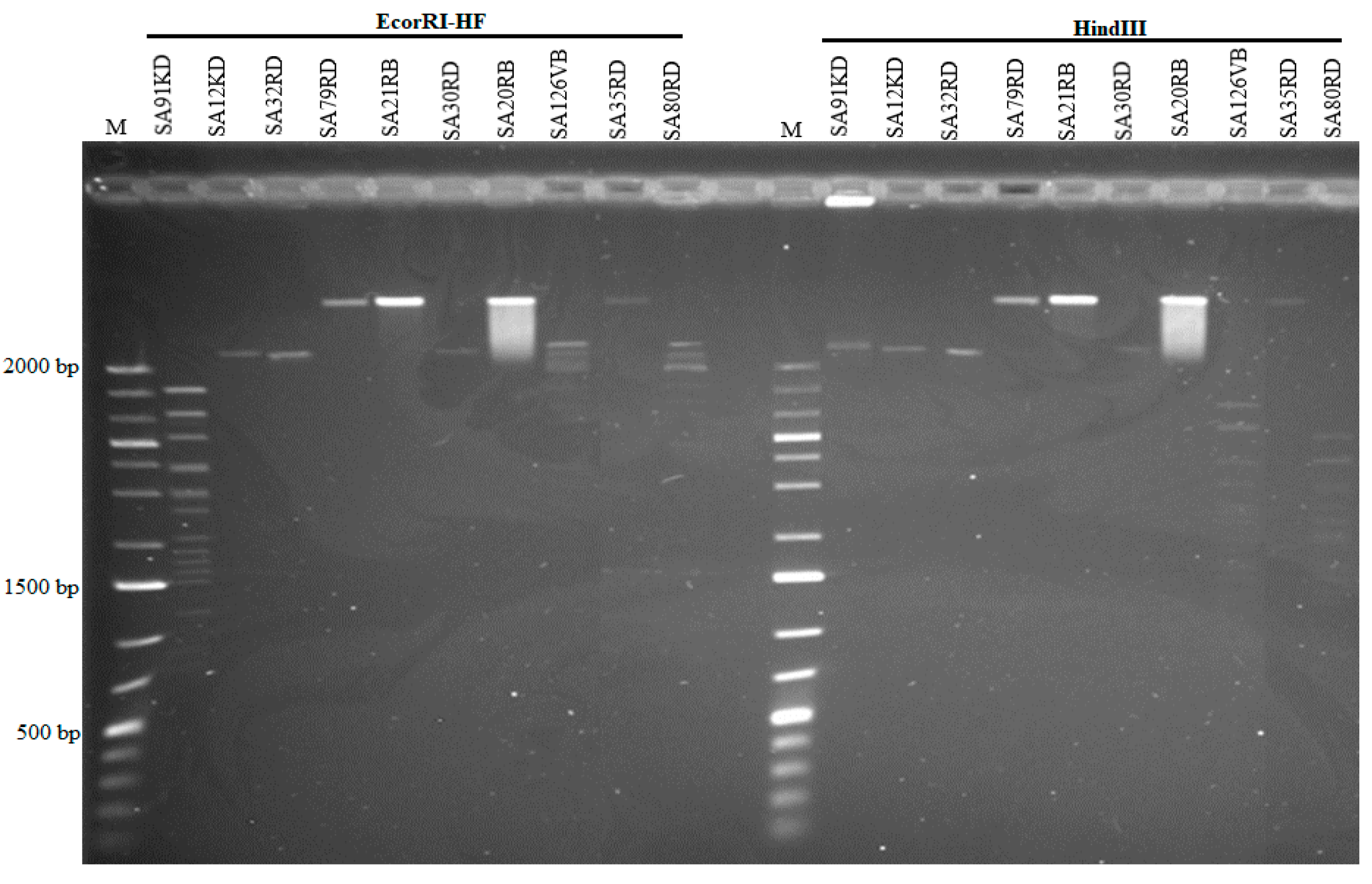
2.3. Genome Size Estimation and Restriction Fragment Length Polymorphism
3. Results
3.1. Isolation and Morphology of the Phages
3.2. Host Range
3.3. Phage Genome Size and Restriction Fragment Length Polymorphisms Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Karmali, M.A. Emerging public health challenges of Shiga toxin–producing Escherichia coli related to changes in the pathogen, the population, and the environment. Clin. Infect. Dis. 2016, 64, 371–376. [Google Scholar]
- USDA-FSIS. Shiga toxin-producing Escherichia coli in certain raw beef products. Fed Reg. 2012, 77, 31975–31981. [Google Scholar]
- Frieri, M.; Kumar, K.; Boutin, A. Antibiotic resistance. J. Infect. Public. Heal. 2017, 10, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Hughes, J.M.; Wilson, M.E.; Johnson, K.E.; Thorpe, C.M.; Sears, C.L. The emerging clinical importance of non-O157 Shiga toxin—producing Escherichia coli. Clin. Infect. Dis. 2006, 43, 1587–1595. [Google Scholar] [CrossRef]
- Valilis, E.; Ramsey, A.; Sidiq, S.; DuPont, H.L. Non-O157 Shiga Toxin-Producing Escherichia coli− a Poorly Appreciated Enteric Pathogen: Systematic Review. Int. J. Infect. Dis. 2018, 76, 82–87. [Google Scholar] [CrossRef]
- Bumunang, E.W.; McAllister, T.A.; Zaheer, R.; Ortega Polo, R.; Stanford, K.; King, R.; Niu, Y.D.; Ateba, C.N. Characterization of Non-O157 Escherichia coli from Cattle Faecal Samples in the North-West Province of South Africa. Microorganisms 2019, 7, 272. [Google Scholar] [CrossRef]
- Smith, J.L.; Fratamico, P.M.; Gunther, N.W., IV. Shiga toxin-producing Escherichia coli. Adv. Appl. Microbiol. 2014, 86, 145–197. [Google Scholar]
- Kortright, K.E.; Chan, B.K.; Koff, J.L.; Turner, P.E. Phage therapy: A renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe. 2019, 25, 219–232. [Google Scholar] [CrossRef]
- Summers, W.C. Felix D’herelle and the Origins of Molecular Biology; Yale University Press: New Haven, CO, USA, 1999; pp. 47–59. [Google Scholar]
- Summers, W.C. Bacteriophage therapy. Annu. Rev. Microbiol. 2001, 55, 437–451. [Google Scholar] [CrossRef]
- Mertz, L. Battling Superbugs: How Phage Therapy Went From Obscure to Promising. IEEE Pulse 2019, 10, 3–9. [Google Scholar] [CrossRef]
- Garrett, L. Seven circles of antimicrobial hell. The Lancet. 2019, 393, 865–867. [Google Scholar] [CrossRef]
- Rohwer, F.; Edwards, R. The Phage Proteomic Tree: a genome-based taxonomy for phage. J. Bacteriol. 2002, 184, 4529–4535. [Google Scholar] [CrossRef] [PubMed]
- Torres-Barceló, C. The disparate effects of bacteriophages on antibiotic-resistant bacteria. Emerg. Microbes. Infect. 2018, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Schmelcher, M.; Loessner, M.J. Application of bacteriophages for detection of foodborne pathogens. Bacteriophage. 2014, 4, e28137. [Google Scholar] [CrossRef] [PubMed]
- Denyes, J.M.; Dunne, M.; Steiner, S.; Mittelviefhaus, M.; Weiss, A.; Schmidt, H.; Klumpp, J.; Loessner, M.J. Modified bacteriophage S16 long tail fiber proteins for rapid and specific immobilization and detection of Salmonella cells. Appl. Environ. Microbiol. 2017, 83. [Google Scholar] [CrossRef]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; De Silva, N.R.; Gargouri, N. World Health Organization global estimates and regional comparisons of the burden of foodborne disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar]
- Fernández, L.; Gutiérrez, D.; Rodríguez, A.; García, P. Application of bacteriophages in the agro-food sector: a long way towards approval. Front. Cell. Infect. Microbiol. 2018, 8, 296. [Google Scholar] [CrossRef]
- FDA. Food additives permitted for direct addition to food for human consumption: bacteriophage preparation. Fed. Regist. 2006, 71, 47729–47732. [Google Scholar]
- Sulakvelidze, A. Using lytic bacteriophages to eliminate or significantly reduce contamination of food by foodborne bacterial pathogens. J. Sci. Food. Agric. 2013, 93, 3137–3146. [Google Scholar] [CrossRef]
- Liao, Y.-T.; Salvador, A.; Harden, L.A.; Liu, F.; Lavenburg, V.M.; Li, R.W.; Wu, V.C. Characterization of a Lytic Bacteriophage as an Antimicrobial Agent for Biocontrol of Shiga Toxin-Producing Escherichia coli O145 Strains. Antibiotics 2019, 8, 74. [Google Scholar] [CrossRef]
- Lopes, A.; Pereira, C.; Almeida, A. Sequential combined effect of phages and antibiotics on the inactivation of Escherichia coli. Microorganisms 2018, 6, 125. [Google Scholar] [CrossRef] [PubMed]
- Ateba, C.N.; Akindolire, M.A. Isolation and Characterisation of Bacteriophages with Lytic Activity Against Virulent Escherichia coli O157: H7: Potential Bio-Control Agents. Preprints 2019. [Google Scholar]
- Wang, J.; Niu, Y.D.; Chen, J.; Anany, H.; Ackermann, H.W.; Johnson, R.P.; Ateba, C.N.; Stanford, K.; McAllister, T.A. Feces of feedlot cattle contain a diversity of bacteriophages that lyse non-O157 Shiga toxin-producing Escherichia coli. Can. J. Microbiol. 2015, 61, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Litt, P.K.; Saha, J.; Jaroni, D. Characterization of bacteriophages targeting Non-O157 Shiga toxigenic Escherichia coli. J. Food. Protect. 2018, 81, 785–794. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.-T.; Quintela, I.A.; Nguyen, K.; Salvador, A.; Cooley, M.B.; Wu, V.C. Investigation of prevalence of free Shiga toxin-producing Escherichia coli (STEC)-specific bacteriophages and its correlation with STEC bacterial hosts in a produce-growing area in Salinas, California. PloS ONE 2018, 13, e0190534. [Google Scholar] [CrossRef]
- Van Twest, R.; Kropinski, A.M. Bacteriophage Enrichment from Water and Soil, Bacteriophages; Springer: Berlin/Heidelberg, Germany, 2009; pp. 15–21. [Google Scholar]
- Sambrook, J.; Russell, D. Molecular Cloning: A Laboratory Manual 1, 3rd ed.; Cold Spring Harbor: New York, NY, USA, 2001. [Google Scholar]
- Niu, Y.D.; Stanford, K.; Ackermann, H.-W.; McAllister, T.A. Characterization of 4 T1-like lytic bacteriophages that lyse Shiga-toxin Escherichia coli O157: H7. Can. J. Microbiol. 2012, 58, 923–927. [Google Scholar] [CrossRef]
- Lingohr, E.; Frost, S.; Johnson, R.P. Determination of bacteriophage genome size by pulsed-field gel electrophoresis. Methods. Mol. Biol. 2009, 502, 19–25. [Google Scholar]
- Kropinski, A.M.; Prangishvili, D.; Lavigne, R.J.E.M. Position paper: the creation of a rational scheme for the nomenclature of viruses of Bacteria and Archaea. Environ. Microbiol. 2009, 11, 2775–2777. [Google Scholar] [CrossRef]
- Ackermann, H. 5500 Phages examined in the electron microscope. Arch. Virol. 2007, 152, 227–243. [Google Scholar] [CrossRef]
- Anany, H.; Switt, A.I.M.; De Lappe, N.; Ackermann, H.-W.; Reynolds, D.M.; Kropinski, A.M.; Wiedmann, M.; Griffiths, M.W.; Tremblay, D.; Moineau, S. A proposed new bacteriophage subfamily: “Jerseyvirinae”. Arch. Virol. 2015, 160, 1021–1033. [Google Scholar] [CrossRef]
- Korf, I.H.; Meier-Kolthoff, J.P.; Adriaenssens, E.M.; Kropinski, A.M.; Nimtz, M.; Rohde, M.; van Raaij, M.J.; Wittmann, J. Still Something to Discover: Novel Insights into Escherichia coli Phage Diversity and Taxonomy. Viruses 2019, 11, 454. [Google Scholar] [CrossRef]
- Turner, D.; Hezwani, M.; Nelson, S.; Salisbury, V.; Reynolds, D. Characterization of the Salmonella bacteriophage vB_SenS-Ent. J. Gen. Virol. 2012, 93, 2046–2056. [Google Scholar] [CrossRef]
- Kang, H.-W.; Kim, J.-W.; Jung, T.-S.; Woo, G.-J. wksl3, a New biocontrol agent for Salmonella enterica serovars enteritidis and typhimurium in foods: characterization, application, sequence analysis, and oral acute toxicity study. Appl. Environ. Microbiol. 2013, 79, 1956–1968. [Google Scholar] [CrossRef]
- Ackermann, H.-W.; Gershman, M. Morphology of phages of a general Salmonella typing set. Res. Virol. 1992, 143, 303–310. [Google Scholar] [CrossRef]
- Chibani-Chennoufi, S.; Sidoti, J.; Bruttin, A.; Dillmann, M.-L.; Kutter, E.; Qadri, F.; Sarker, S.A.; Brüssow, H. Isolation of Escherichia coli bacteriophages from the stool of pediatric diarrhea patients in Bangladesh. J. Bacteriol. 2004, 186, 8287–8294. [Google Scholar] [CrossRef]
- Joshi, A.; Siddiqi, J.; Rao, G.; Chakravorty, M. MB78, a virulent bacteriophage of Salmonella typhimurium. J. Virol. 1982, 41, 1038–1043. [Google Scholar]
- Yan, J.; Mao, J.; Xie, J. Bacteriophage polysaccharide depolymerases and biomedical applications. BioDrugs 2014, 28, 265–274. [Google Scholar] [CrossRef]
- Pires, D.P.; Oliveira, H.; Melo, L.D.; Sillankorva, S.; Azeredo, J. Bacteriophage-encoded depolymerases: their diversity and biotechnological applications. Appl. Microbiol. Biotechnol. 2016, 100, 2141–2151. [Google Scholar] [CrossRef]
- Guo, Z.; Huang, J.; Yan, G.; Lei, L.; Wang, S.; Yu, L.; Zhou, L.; Gao, A.; Feng, X.; Han, W. Identification and characterization of Dpo42, a novel depolymerase derived from the Escherichia coli phage vB_EcoM_ECOO78. Front. Microbiol. 2017, 8, 1460. [Google Scholar]
- Gutiérrez, D.; Briers, Y.; Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; Lavigne, R.; García, P. Role of the pre-neck appendage protein (Dpo7) from phage vB_SepiS-phiIPLA7 as an anti-biofilm agent in Staphylococcal species. Front. Microbiol. 2015, 6, 1315. [Google Scholar] [CrossRef]
- Torres-Barceló, C.; Hochberg, M.E. Evolutionary rationale for phages as complements of antibiotics. Trends. Microbiol. 2016, 24, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T.; Yin, J. Bacteriophage plaques: Theory and analysis, Bacteriophages; Springer: Berlin/Heidelberg, Germany, 2009; pp. 161–174. [Google Scholar]
- Gallet, R.; Kannoly, S.; Wang, N. Effects of bacteriophage traits on plaque formation. BMC. Microbiol. 2011, 11, 181. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.-T.; Sun, X.; Quintela, I.A.; Bridges, D.F.; Liu, F.; Zhang, Y.; Salvador, A.; Wu, V.C. Discovery of Shiga Toxin-Producing Escherichia coli (STEC)-Specific Bacteriophages From Non-fecal Composts Using Genomic Characterization. Front. Microbiol. 2019, 10, 627. [Google Scholar] [CrossRef] [PubMed]
- Casey, E.; van Sinderen, D.; Mahony, J. In vitro characteristics of phages to guide ‘real life’phage therapy suitability. Viruses 2018, 10, 163. [Google Scholar] [CrossRef]
- Bertozzi Silva, J.; Storms, Z.; Sauvageau, D. Host receptors for bacteriophage adsorption. FEMS Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef]
- Hyman, P. Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth. Pharmaceuticals 2019, 12, 35. [Google Scholar] [CrossRef]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317. [Google Scholar] [CrossRef]
- Warren, R. Modified bases in bacteriophage DNAs. Annu. Rev. Microbiol. 1980, 34, 137–158. [Google Scholar] [CrossRef]
- Ghasemian, A.; Moradpour, Z.; Bavand, M. A broad-host range coliphage against a clinically isolated E. coli O157: isolation and characterization. J. Appl. Pharm. Sci. 2017, 7, 123–128. [Google Scholar]
- Shi, D.-Q.; Ali, I.; Tang, J.; Yang, W.-C. New insights into 5hmC DNA modification: generation, distribution and function. Front. Genet. 2017, 8, 100. [Google Scholar] [CrossRef]
- Hattman, S. The first recognized epigenetic signal: DNA glucosylation of T-even bacteriophages. Epigenetics 2009, 4, 150–151. [Google Scholar] [CrossRef]
- Raleigh, E.A.; Wilson, G. Escherichia coli K-12 restricts DNA containing 5-methylcytosine. Proc. Natl. Acad. Sci. USA 1986, 83, 9070–9074. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Area | Phage ID | Family | Subfamily | Head Dimension (nm) | Tail Dimension (nm) | ||
|---|---|---|---|---|---|---|---|
| Width | Height | Width | Height | ||||
| Rooigrond dairy | SA79RD | Myoviridae | - | 100 ± 0 | 133 ± 0 | 24 ± 2 | 133 ± 0 |
| Koster dairy | SA91KD | Myoviridae | - | 70 ± 3 | 73 ± 4 | 23 ± 0 | 154 ± 4 |
| Rooigrond beef | SA20RB | Myoviridae | - | 100 ± 0 | 129 ± 4 | 25 ± 2 | 134 ± 2 |
| Rooigrond beef | SA21RB | Myoviridae | - | 82 ± 2 | 129 ± 4 | 25 ± 4 | 132 ± 2 |
| Rooigrond dairy | SA35RD | Myoviridae | - | 95 ± 7 | 133 ± 2 | 24 ± 2 | 133 ± 2 |
| Rooigrond dairy | SA80RD | Siphoviridae | - | 50 ± 6 | 67 ± 0 | 9 ± 2 | 200 ± 0 |
| Koster dairy | SA12KD | Siphoviridae | “Jerseyvirinae” | 70 ± 0 | 78 ± 2 | 13 ± 2 | 197 ± 0 |
| Vryburg beef | SA126VB | Siphoviridae | “Jerseyvirinae” | 67 ± 7 | 81 ± 4 | 12 ± 2 | 191 ± 2 |
| Rooigrond dairy | SA30RD | Siphoviridae | “Jerseyvirinae” | 67 ± 0 | 67 ± 0 | 10 ± 0 | 200 ± 3 |
| Rooigrond dairy | SA32RD | Siphoviridae | “Jerseyvirinae” | 67 ± 0 | 69 ± 2 | 10 ± 0 | 174 ± 2 |
| Serogroup | Bacteriophage | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| SA20RB | SA79RD | SA126VB | SA30RD | SA32RD | SA35RD | SA21RB | SA80RD | SA12KD | SA91KD | |
| O156:H25 | + | + | - | - | - | + | + | - | - | + |
| O108:H2 | + | + | + | - | - | + | + | - | - | + |
| O136:H30 | - | - | - | - | - | - | - | - | - | - |
| O99:H9 | - | - | - | - | - | - | - | - | - | - |
| wzx-Onovel24:H20 | + | + | - | - | - | + | + | - | - | - |
| O140:H21 | + | - | - | - | - | + | + | - | - | - |
| O102:H4 | - | - | + | - | - | - | - | - | - | - |
| O129:H23 | - | - | - | - | - | - | - | - | - | - |
| O17:H18 | - | + | - | - | + | + | - | + | - | - |
| O76:H34 | - | - | - | - | - | - | - | - | - | - |
| O26:H11 | - | + | - | - | - | + | - | - | - | - |
| O129:H23 | - | - | - | - | - | - | - | - | - | - |
| O154:H10 | + | - | - | - | - | - | + | - | - | - |
| O116:H21 | - | + | - | - | - | + | + | - | - | - |
| wzx-Onovel5:H19 | + | + | + | + | - | + | + | - | - | + |
| O87:H7 | + | + | - | - | - | + | + | - | - | - |
| O129:H21 | - | + | - | - | - | + | + | - | - | - |
| O26:H11 | - | + | - | - | - | + | - | - | - | - |
| O26:H11 | - | - | - | - | - | - | + | - | - | - |
| O163:H19 | + | + | + | + | - | + | - | - | - | + |
| O40:H19 | + | + | - | - | - | + | - | - | - | - |
| O22:H21 | + | - | - | - | - | - | - | - | - | - |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bumunang, E.W.; McAllister, T.A.; Stanford, K.; Anany, H.; Niu, Y.D.; Ateba, C.N. Characterization of Non-O157 STEC Infecting Bacteriophages Isolated from Cattle Faeces in North-West South Africa. Microorganisms 2019, 7, 615. https://doi.org/10.3390/microorganisms7120615
Bumunang EW, McAllister TA, Stanford K, Anany H, Niu YD, Ateba CN. Characterization of Non-O157 STEC Infecting Bacteriophages Isolated from Cattle Faeces in North-West South Africa. Microorganisms. 2019; 7(12):615. https://doi.org/10.3390/microorganisms7120615
Chicago/Turabian StyleBumunang, Emmanuel W., Tim A. McAllister, Kim Stanford, Hany Anany, Yan D. Niu, and Collins N. Ateba. 2019. "Characterization of Non-O157 STEC Infecting Bacteriophages Isolated from Cattle Faeces in North-West South Africa" Microorganisms 7, no. 12: 615. https://doi.org/10.3390/microorganisms7120615
APA StyleBumunang, E. W., McAllister, T. A., Stanford, K., Anany, H., Niu, Y. D., & Ateba, C. N. (2019). Characterization of Non-O157 STEC Infecting Bacteriophages Isolated from Cattle Faeces in North-West South Africa. Microorganisms, 7(12), 615. https://doi.org/10.3390/microorganisms7120615