Gut Anaerobes Capable of Chicken Caecum Colonisation

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Bacterial Isolates
2.3. Chicken Inoculation with Gut Anaerobes and S. Enteritidis Challenge
2.4. Oral Inoculation with Defined Mixtures of Gut Anaerobes
2.5. Salmonella Enteritidis Challenge
2.6. Chicken Gene Expression
2.7. Sequencing of V3/V4 Region of 16S rRNA Genes
2.8. Statistics
3. Results
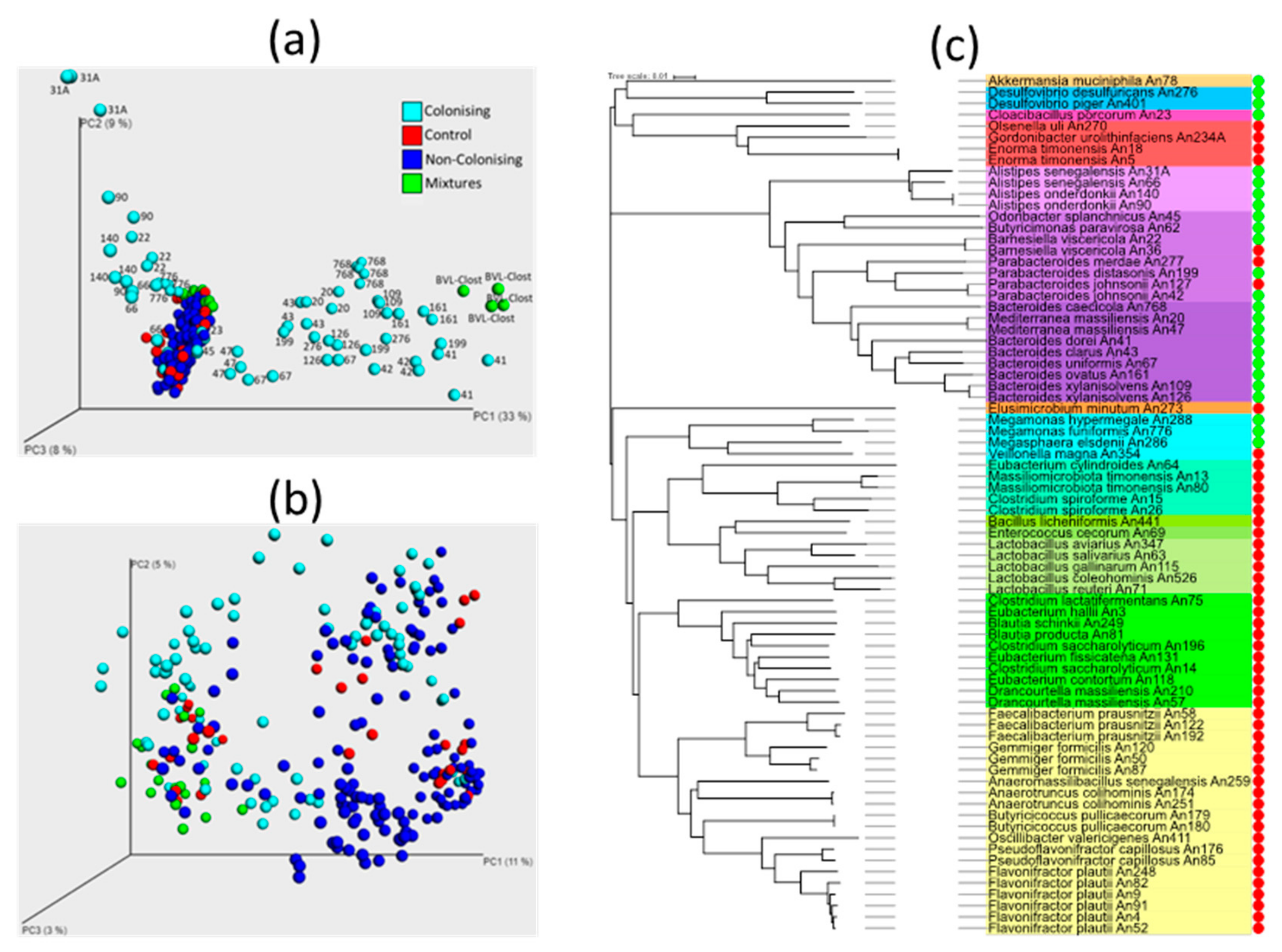
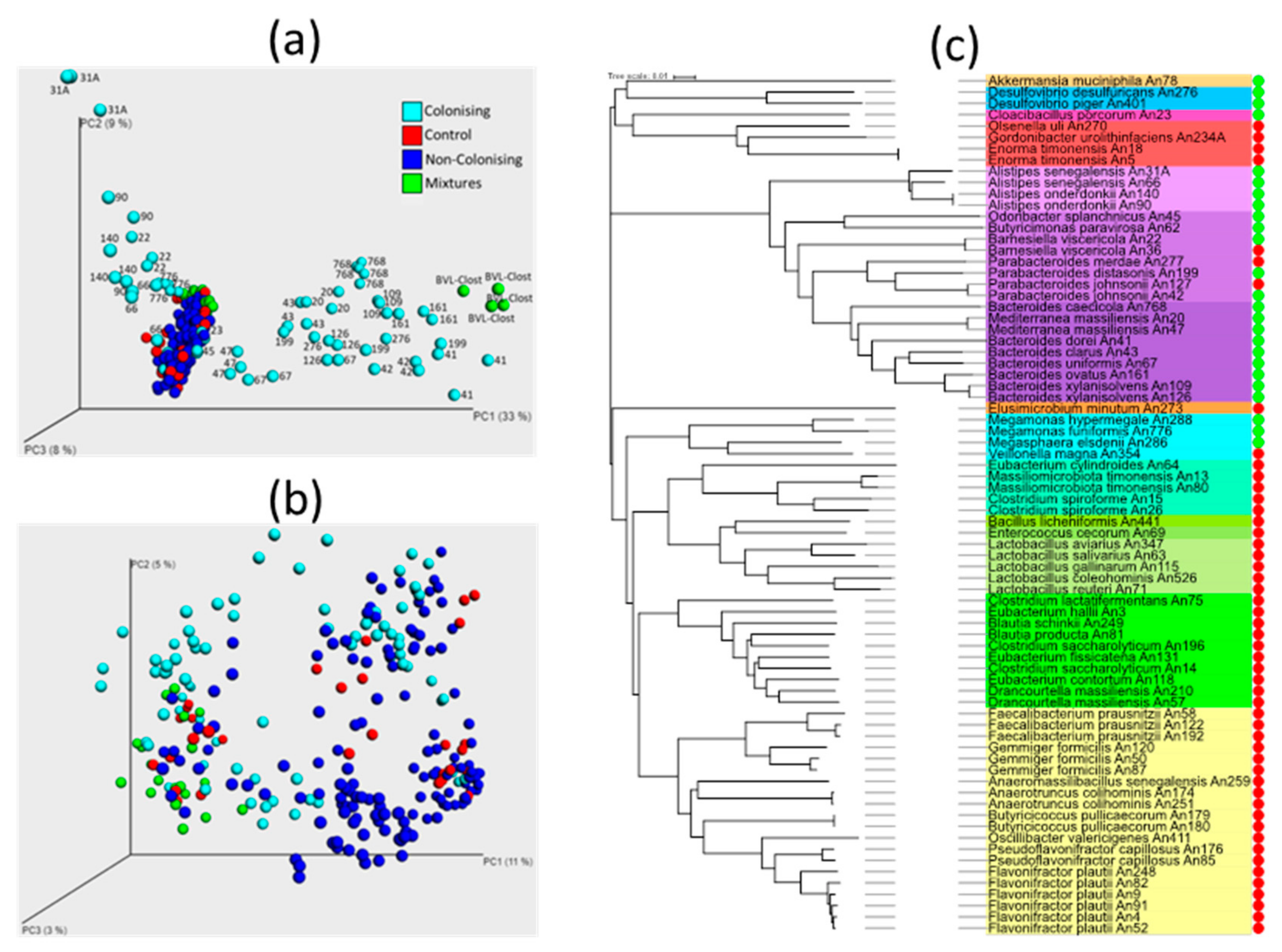
3.1. Inoculation of Newly Hatched Chickens with Individual Bacterial Isolates
3.2. Additional Experiments to Test (In)Ability of Gram Positive Bacteria to Colonise The Chicken Caecum
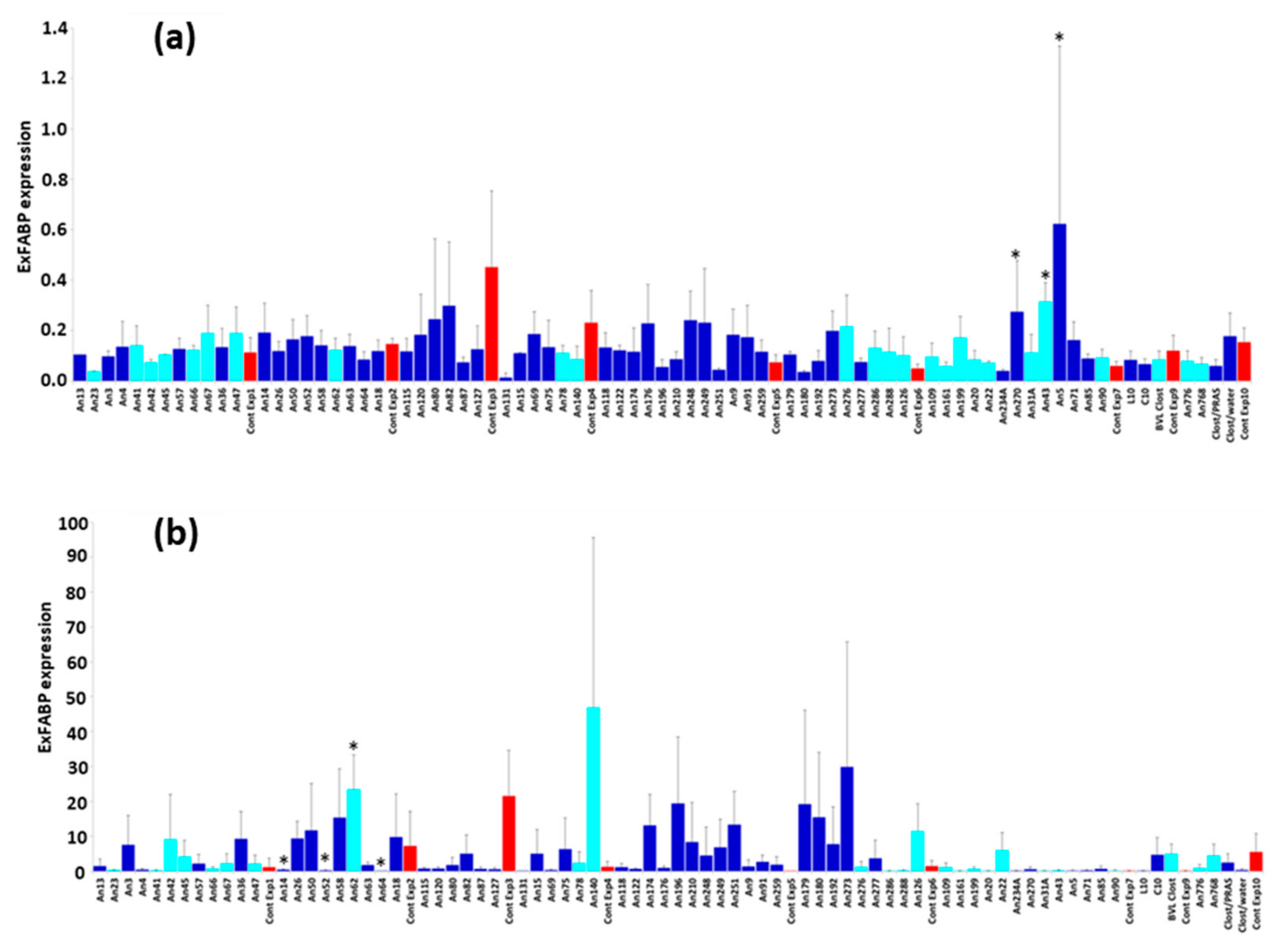
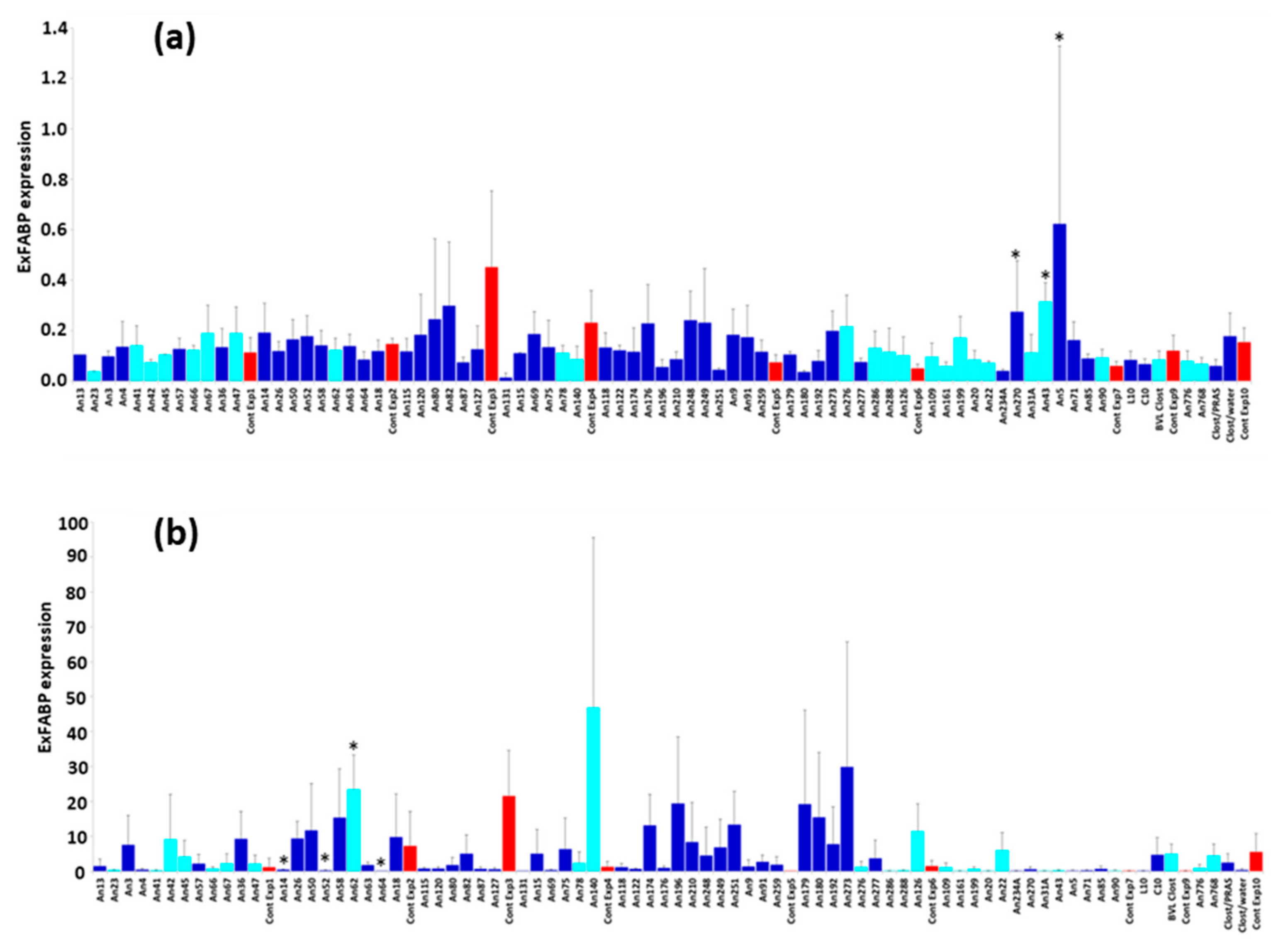
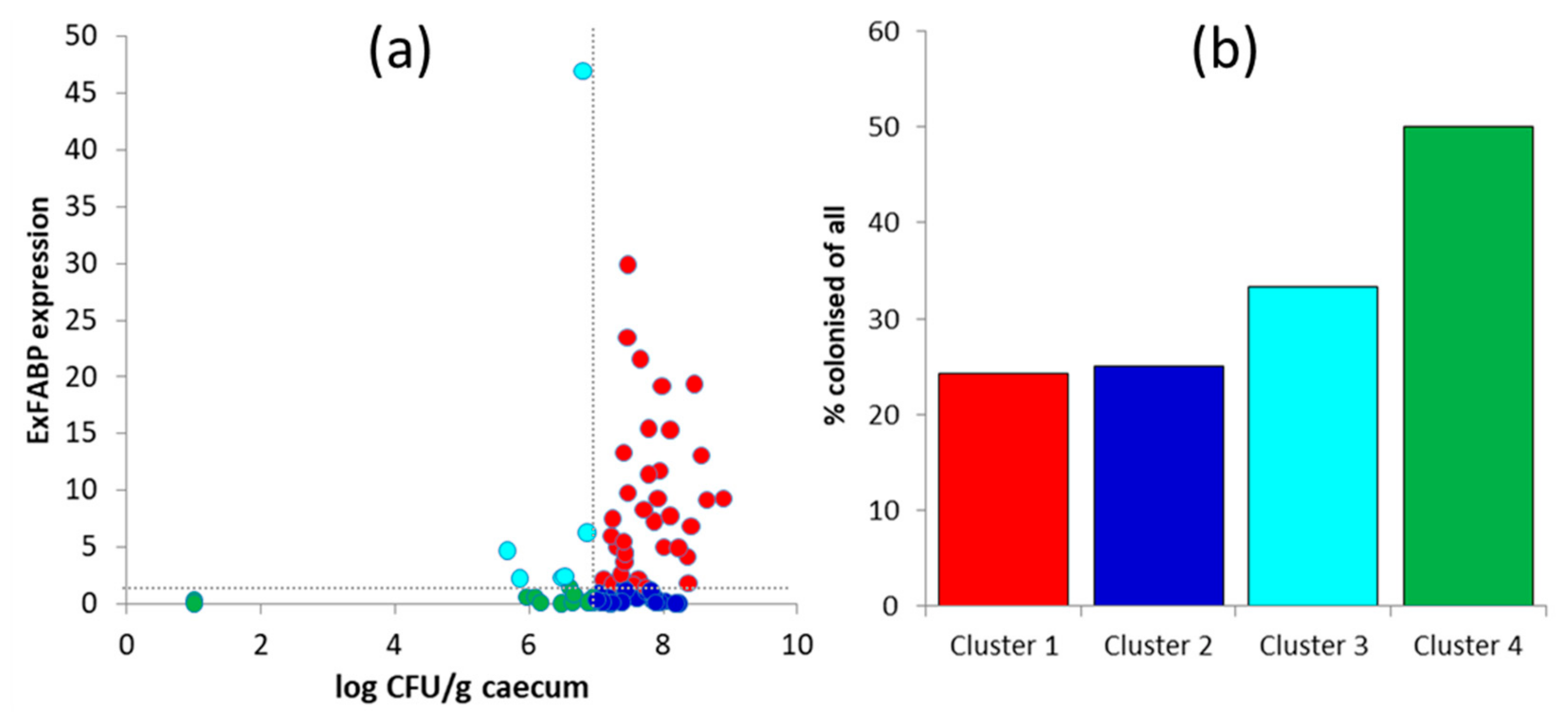
3.3. Chicken Response to Colonisation with Tested Anaerobes
3.4. Chicken Response to S. Enteritidis Infection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Awad, W.A.; Mann, E.; Dzieciol, M.; Hess, C.; Schmitz-Esser, S.; Wagner, M.; Hess, M. Age-related differences in the luminal and mucosa-associated gut microbiome of broiler chickens and shifts associated with Campylobacter jejuni infection. Front. Cell. Infect. Microbiol. 2016, 6, 154. [Google Scholar] [CrossRef]
- Ranjitkar, S.; Lawley, B.; Tannock, G.; Engberg, R.M. Bacterial succession in the broiler gastrointestinal tract. Appl. Environ. Microbiol. 2016, 82, 2399–2410. [Google Scholar] [CrossRef]
- Videnska, P.; Sedlar, K.; Lukac, M.; Faldynova, M.; Gerzova, L.; Cejkova, D.; Sisak, F.; Rychlik, I. Succession and replacement of bacterial populations in the caecum of egg laying hens over their whole life. PLoS ONE 2014, 9, e115142. [Google Scholar] [CrossRef]
- Videnska, P.; Rahman, M.M.; Faldynova, M.; Babak, V.; Matulova, M.E.; Prukner-Radovcic, E.; Krizek, I.; Smole-Mozina, S.; Kovac, J.; Szmolka, A.; et al. Characterization of egg laying hen and broiler fecal microbiota in poultry farms in Croatia, Czech Republic, Hungary and Slovenia. PLoS ONE 2014, 9, e110076. [Google Scholar] [CrossRef]
- Varmuzova, K.; Kubasova, T.; Davidova-Gerzova, L.; Sisak, F.; Havlickova, H.; Sebkova, A.; Faldynova, M.; Rychlik, I. Composition of gut microbiota influences resistance of newly hatched chickens to Salmonella enteritidis infection. Front. Microbiol. 2016, 7, 957. [Google Scholar] [CrossRef] [PubMed]
- Polansky, O.; Sekelova, Z.; Faldynova, M.; Sebkova, A.; Sisak, F.; Rychlik, I. Important metabolic pathways and biological processes expressed by chicken cecal microbiota. Appl. Environ. Microbiol. 2015, 82, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Kaldhusdal, M.; Schneitz, C.; Hofshagen, M.; Skjerve, E. Reduced incidence of Clostridium perfringens-associated lesions and improved performance in broiler chickens treated with normal intestinal bacteria from adult fowl. Avian Dis. 2001, 45, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Salvat, G.; Lalande, F.; Humbert, F.; Lahellec, C. Use of a competitive exclusion product (Broilact) to prevent Salmonella colonization of newly hatched chicks. Int. J. Food Microbiol. 1992, 15, 307–311. [Google Scholar] [CrossRef]
- Nisbet, D. Defined competitive exclusion cultures in the prevention of enteropathogen colonisation in poultry and swine. Antonie Van Leeuwenhoek 2002, 81, 481–486. [Google Scholar] [CrossRef]
- Rantala, M.; Nurmi, E. Prevention of the growth of Salmonella infantis in chicks by the flora of the alimentary tract of chickens. Br. Poult. Sci. 1973, 14, 627–630. [Google Scholar] [CrossRef]
- Stavric, S. Defined cultures and prospects. Int. J. Food Microbiol. 1992, 15, 245–263. [Google Scholar] [CrossRef]
- Baldwin, S.; Hughes, R.J.; Hao Van, T.T.; Moore, R.J.; Stanley, D. At-hatch administration of probiotic to chickens can introduce beneficial changes in gut microbiota. PLoS ONE 2018, 13, e0194825. [Google Scholar] [CrossRef] [PubMed]
- Beirao, B.C.B.; Ingberman, M.; Favaro, C., Jr.; Mesa, D.; Bittencourt, L.C.; Fascina, V.B.; Caron, L.F. Effect of an Enterococcus faecium probiotic on specific IgA following live Salmonella Enteritidis vaccination of layer chickens. Avian Pathol. 2018, 47, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Mazanko, M.S.; Gorlov, I.F.; Prazdnova, E.V.; Makarenko, M.S.; Usatov, A.V.; Bren, A.B.; Chistyakov, V.A.; Tutelyan, A.V.; Komarova, Z.B.; Mosolova, N.I.; et al. Bacillus probiotic supplementations improve laying performance, egg quality, hatching of laying hens, and sperm quality of roosters. Probiotics Antimicrob. Proteins 2018, 10, 367–373. [Google Scholar] [CrossRef] [PubMed]
- Weinack, O.M.; Snoeyenbos, G.H.; Soerjadi-Liem, A.S. Further studies on competitive exclusion of Salmonella typhimurium by lactobacilli in chickens. Avian Dis. 1985, 29, 1273–1276. [Google Scholar] [CrossRef] [PubMed]
- Adler, H.E.; DaMassa, A.J. Effect of ingested Lactobacilli on Salmonella infantis and Escherichia coli and on intestinal flora, pasted vents, and chick growth. Avian Dis. 1980, 24, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Medvecky, M.; Cejkova, D.; Polansky, O.; Karasova, D.; Kubasova, T.; Cizek, A.; Rychlik, I. Whole genome sequencing and function prediction of 133 gut anaerobes isolated from chicken caecum in pure cultures. BMC Genom. 2018, 19, 561. [Google Scholar] [CrossRef]
- Crhanova, M.; Karasova, D.; Juricova, H.; Matiasovicova, J.; Jahodarova, E.; Kubasova, T.; Seidlerova, Z.; Cizek, A.; Rychlik, I. Systematic culturomics shows that half of chicken caecal microbiota members can be grown in vitro except for two lineages of Clostridiales and a single lineage of Bacteroidetes. Microorganisms 2019, 7, 496. [Google Scholar] [CrossRef]
- Methner, U.; Barrow, P.A.; Gregorova, D.; Rychlik, I. Intestinal colonisation-inhibition and virulence of Salmonella phoP, rpoS and ompC deletion mutants in chickens. Vet. Microbiol. 2004, 98, 37–43. [Google Scholar] [CrossRef]
- Matulova, M.; Rajova, J.; Vlasatikova, L.; Volf, J.; Stepanova, H.; Havlickova, H.; Sisak, F.; Rychlik, I. Characterization of chicken spleen transcriptome after infection with Salmonella enterica serovar Enteritidis. PLoS ONE 2012, 7, e48101. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Stanley, D.; Hughes, R.J.; Geier, M.S.; Moore, R.J. Bacteria within the gastrointestinal tract microbiota correlated with improved growth and feed conversion: Challenges presented for the identification of performance enhancing probiotic bacteria. Front. Microbiol. 2016, 7, 187. [Google Scholar] [CrossRef]
- Impey, C.S.; Mead, G.C.; George, S.M. Competitive exclusion of salmonellas from the chick caecum using a defined mixture of bacterial isolates from the caecal microflora of an adult bird. J. Hyg. Lond. 1982, 89, 479–490. [Google Scholar] [CrossRef] [PubMed]
- Kubasova, T.; Kollarcikova, M.; Crhanova, M.; Karasova, D.; Cejkova, D.; Sebkova, A.; Matiasovicova, J.; Faldynova, M.; Pokorna, A.; Cizek, A.; et al. Contact with adult hen affects development of caecal microbiota in newly hatched chicks. PLoS ONE 2019, 14, e0212446. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Species | ID | Phylum | % of Total Microbiota |
|---|---|---|---|
| Alistipes onderdonkii | An140 | Bacteroidetes | 34.24 |
| Alistipes onderdonkii | An90 | Bacteroidetes | 33.54 |
| Alistipes senegalensis | An31A | Bacteroidetes | 78.03 |
| Alistipes senegalensis | An66 | Bacteroidetes | 1.74 |
| Bacteroides caecicola | An768 | Bacteroidetes | 63.01 |
| Bacteroides clarus | An43 | Bacteroidetes | 29.92 |
| Bacteroides dorei | An41 | Bacteroidetes | 52.51 |
| Bacteroides ovatus | An161 | Bacteroidetes | 31.35 |
| Bacteroides uniformis | An67 | Bacteroidetes | 23.85 |
| Bacteroides xylanisolvens | An109 | Bacteroidetes | 13.09 |
| Bacteroides xylanisolvens | An126 | Bacteroidetes | 4.96 |
| Odoribacter splanchnicus | An45 | Bacteroidetes | 0.83 |
| Butyricimonas paravirosa | An62 | Bacteroidetes | 0.72 |
| Barnesiella viscericola | An22 | Bacteroidetes | 65.08 |
| Mediterranea massiliensis | An20 | Bacteroidetes | 47.46 |
| Mediterranea massiliensis | An47 | Bacteroidetes | 11.36 |
| Parabacteroides distasonis | An199 | Bacteroidetes | 39.76 |
| Parabacteroides johnsonii | An42 | Bacteroidetes | 25.18 |
| Megamonas funiformis | An776 | Firmicutes/Negativicutes | 53.32 |
| Megamonas hypermegale | An288 | Firmicutes/Negativicutes | 8.62 |
| Megasphaera elsdenii | An286 | Firmicutes/Negativicutes | 0.98 |
| Desulfovibrio piger | An401 | Proteobacteria | 1.64 |
| Desulfovibrio desulfuricans | An276 | Proteobacteria | 0.65 |
| Akkermansia muciniphila | An78 | Verrucomicrobia | 13.08 |
| Cloacibacillus porcorum | An23 | Synergistetes | 0.25 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kubasova, T.; Kollarcikova, M.; Crhanova, M.; Karasova, D.; Cejkova, D.; Sebkova, A.; Matiasovicova, J.; Faldynova, M.; Sisak, F.; Babak, V.; et al. Gut Anaerobes Capable of Chicken Caecum Colonisation. Microorganisms 2019, 7, 597. https://doi.org/10.3390/microorganisms7120597
Kubasova T, Kollarcikova M, Crhanova M, Karasova D, Cejkova D, Sebkova A, Matiasovicova J, Faldynova M, Sisak F, Babak V, et al. Gut Anaerobes Capable of Chicken Caecum Colonisation. Microorganisms. 2019; 7(12):597. https://doi.org/10.3390/microorganisms7120597
Chicago/Turabian StyleKubasova, Tereza, Miloslava Kollarcikova, Magdalena Crhanova, Daniela Karasova, Darina Cejkova, Alena Sebkova, Jitka Matiasovicova, Marcela Faldynova, Frantisek Sisak, Vladimir Babak, and et al. 2019. "Gut Anaerobes Capable of Chicken Caecum Colonisation" Microorganisms 7, no. 12: 597. https://doi.org/10.3390/microorganisms7120597
APA StyleKubasova, T., Kollarcikova, M., Crhanova, M., Karasova, D., Cejkova, D., Sebkova, A., Matiasovicova, J., Faldynova, M., Sisak, F., Babak, V., Pokorna, A., Cizek, A., & Rychlik, I. (2019). Gut Anaerobes Capable of Chicken Caecum Colonisation. Microorganisms, 7(12), 597. https://doi.org/10.3390/microorganisms7120597