The Effects of Flavomycin and Colistin Sulfate Pre-Treatment on Ileal Bacterial Community Composition, the Response to Salmonella typhimurium and Host Gene Expression in Broiler Chickens

Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sample Collection
2.2. DNA Extraction and 16S rDNA Sequencing of Ileal Microbes
2.3. Ileum Histomorphology
2.4. Detection of Ileal Permeability by Serum Method
2.5. Myeloperoxidase (MPO) Activity in the Ileal Mucosa
2.6. Total RNA Extraction and Quantitative Real Time PCR
2.7. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Ileal Morphology
3.3. Ileal Gene Expression on the 14th Day
3.4. Ileal Gene Expression on the 21st Day
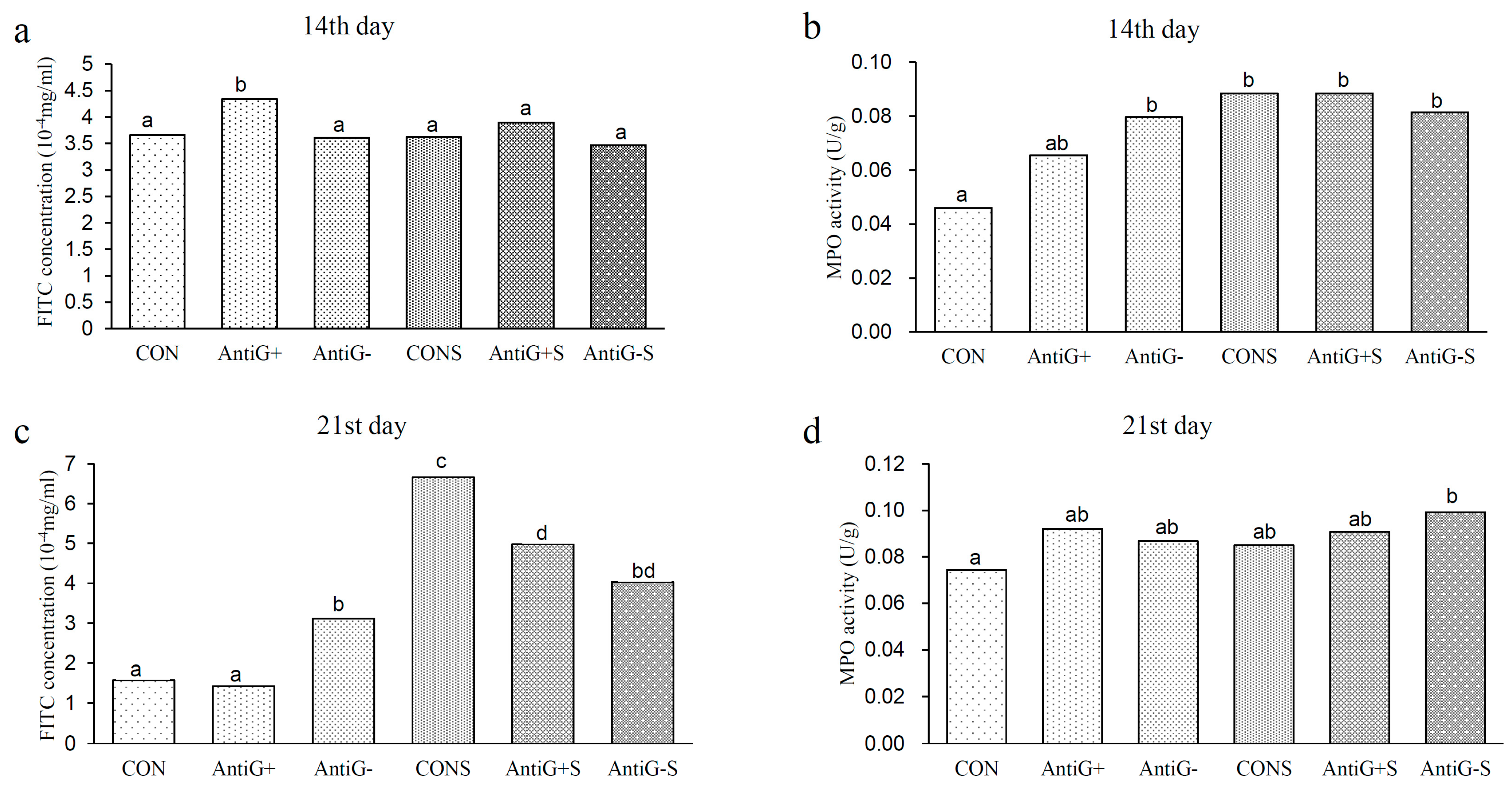
3.5. Intestinal Permeability and MPO Activity
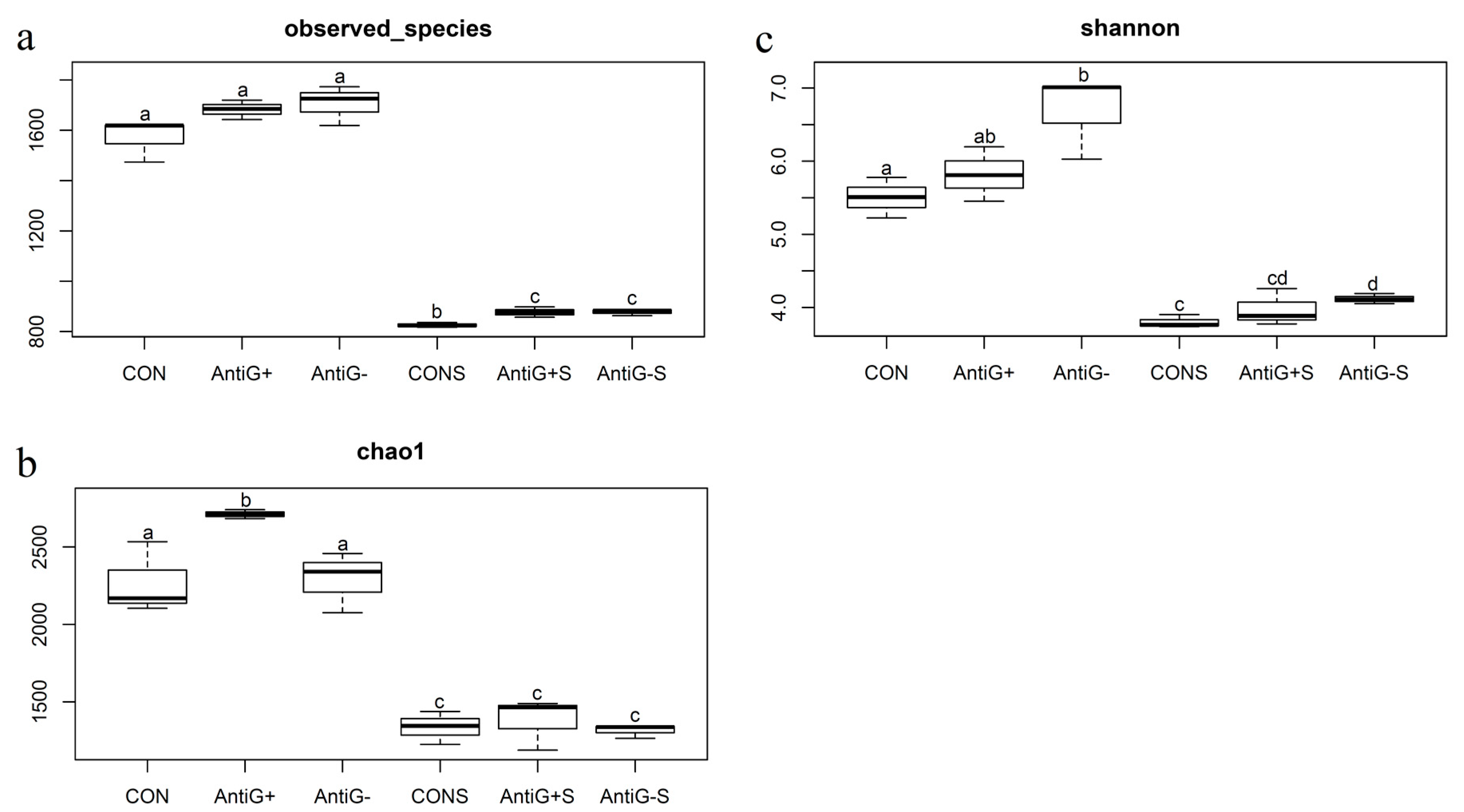
3.6. Alpha Diversity Analysis
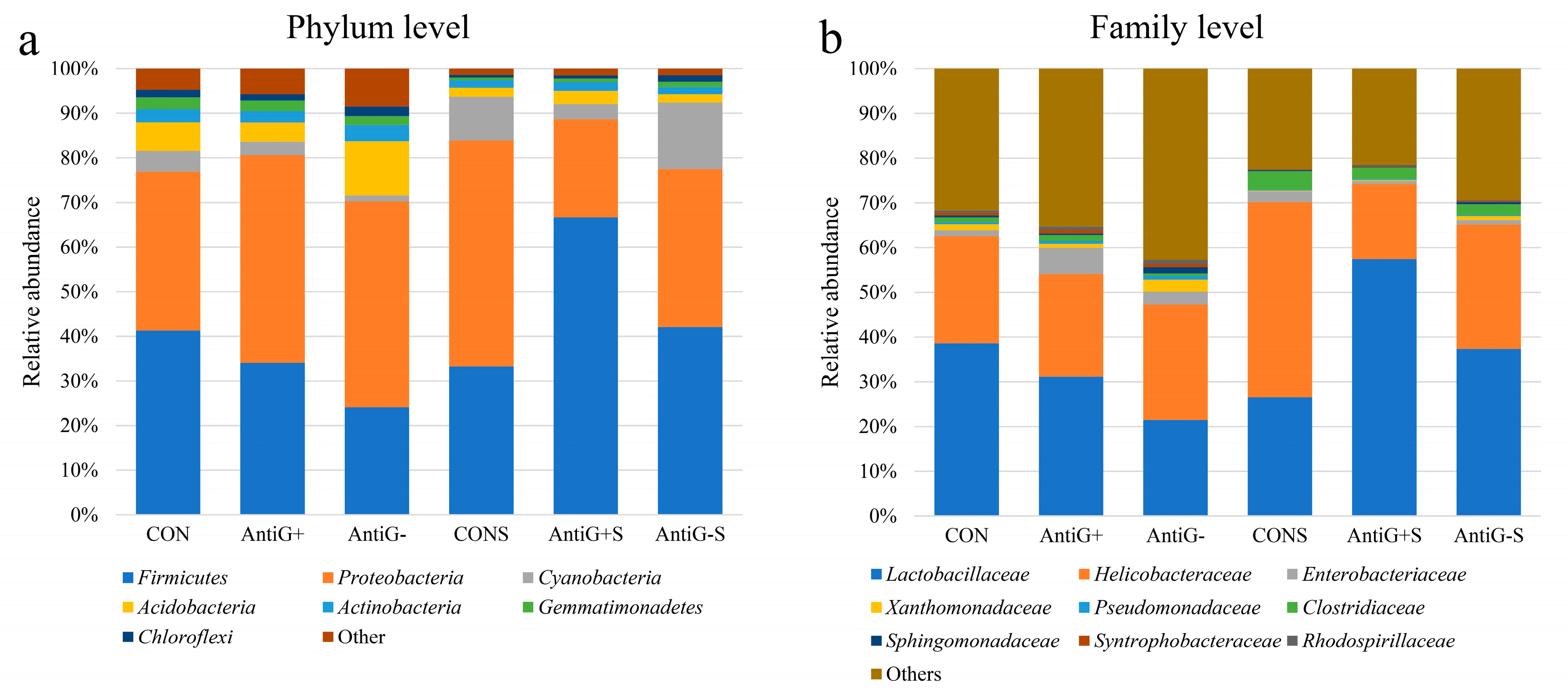
3.7. Taxonomic Analysis
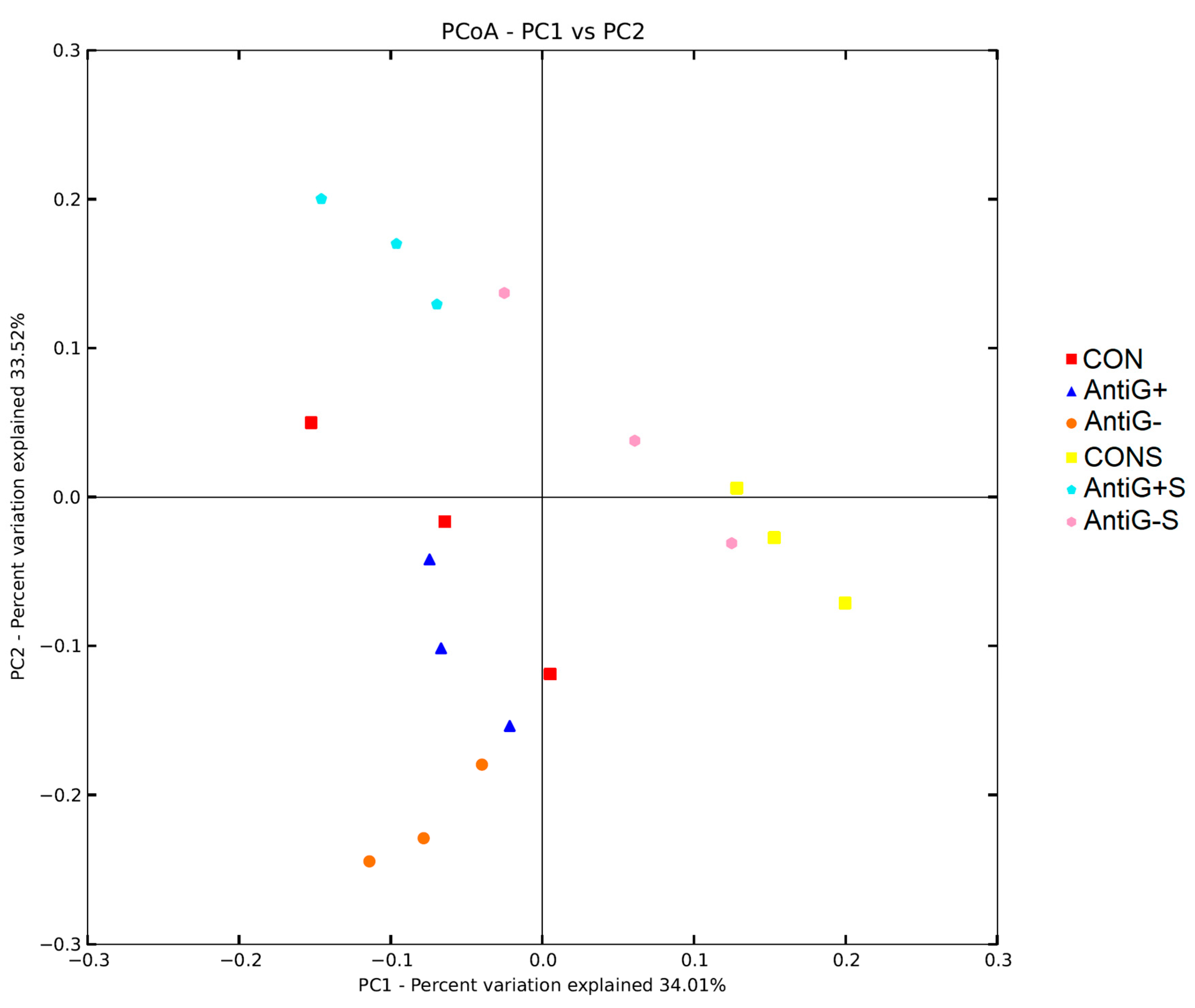
3.8. Beta Diversity
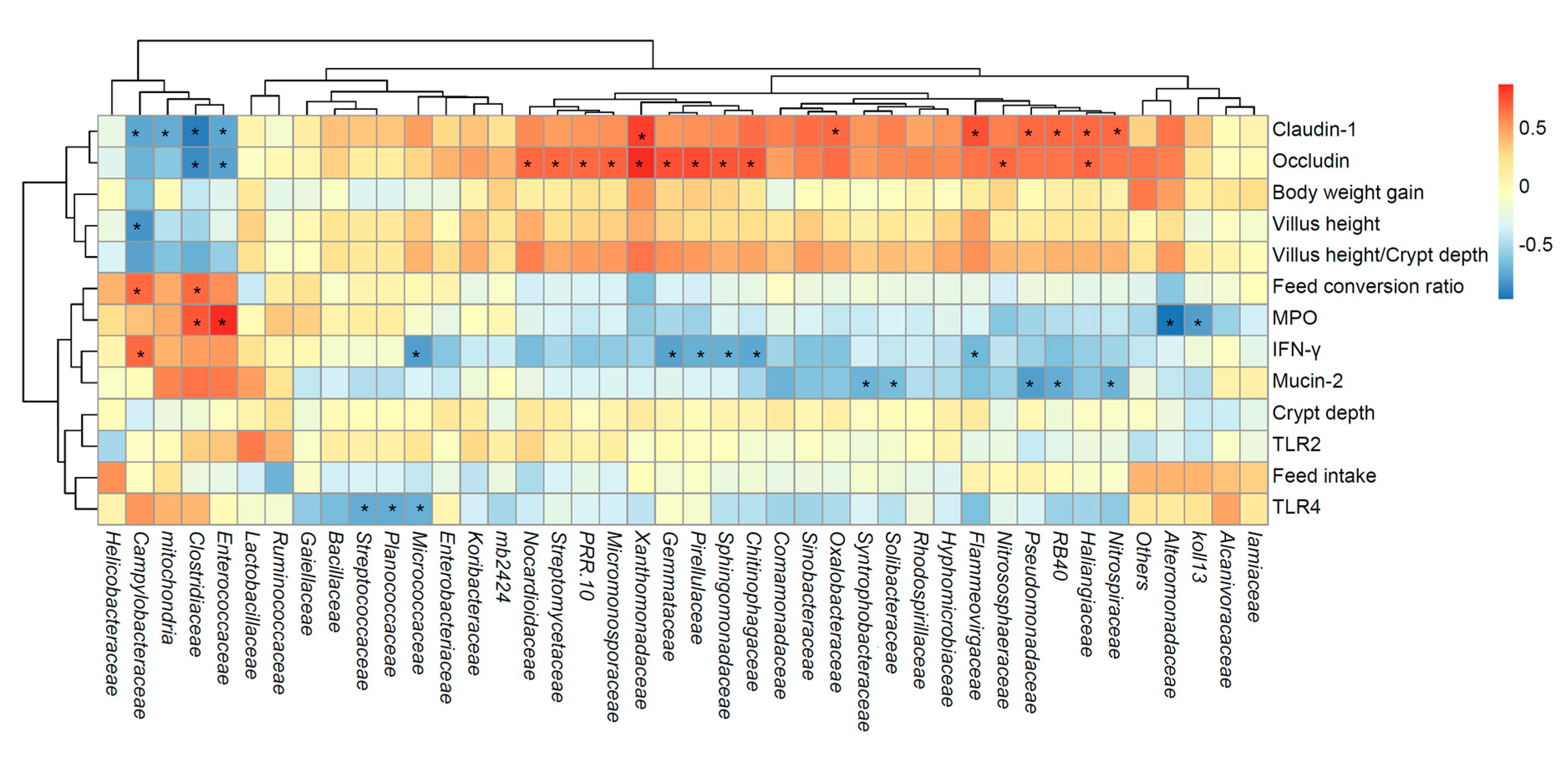
3.9. Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pulido-Landinez, M. Food safety-Salmonella update in broilers. Anim. Feed Sci. Tech. 2019, 250, 53–58. [Google Scholar] [CrossRef]
- Lamas, A.; Miranda, J.M.; Regal, P.; Vázquez, B.; Franco, C.M.; Cepeda, A. A comprehensive review of non- enterica subspecies of Salmonella enterica. Microbiol. Res. 2018, 206, 60–73. [Google Scholar] [CrossRef] [PubMed]
- Thung, T.Y.; Mahyudin, N.A.; Basri, D.F.; Radzi, C.W.J.W.; Nakaguchi, Y.; Nishibuchi, M.; Radu, S. Prevalence and antibiotic resistance of Salmonella Enteritidis and Salmonella Typhimurium in raw chicken meat at retail markets in Malaysia. Poult. Sci. 2016, 95, 1888–1893. [Google Scholar] [CrossRef] [PubMed]
- Lamas, A.; Fernandez-No, I.C.; Miranda, J.M.; Vazquez, B.; Cepeda, A.; Franco, C.M. Prevalence, molecular characterization and antimicrobial resistance of Salmonella serovars isolated from northwestern Spanish broiler flocks (2011–2015). Poult. Sci. 2016, 95, 2097–2105. [Google Scholar] [CrossRef] [PubMed]
- Wyrsch, E.R.; Hawkey, J.; Judd, L.M.; Haites, R.; Holt, K.E.; Djordjevic, S.P.; Billman-Jacobe, H. Z/I1 Hybrid Virulence Plasmids Carrying Antimicrobial Resistance genes in S. Typhimurium from Australian Food Animal Production. Microorganisms 2019, 7, 299. [Google Scholar] [CrossRef] [PubMed]
- Willis, W.L.; King, K.; Iskhuemhen, O.S.; Ibrahim, S.A. Administration of mushroom extract to broiler chickens for bifidobacteria enhancement and Salmonella reduction. J. Appl. Poult. Res. 2009, 18, 658–664. [Google Scholar] [CrossRef]
- LeBlanc, D.; de Moreno, A.; Castillo, N.A.; Perdigon, G. Anti-infective mechanisms induced by a probiotic Lactobacillus strain against Salmonella enterica serovar Typhimurium infection. Int. J. Food Microbiol. 2010, 138, 223–231. [Google Scholar] [CrossRef]
- Lin, S.; Guo, L. Flavomycin and its applications in the breeding industry. Nat. Prod. Res. Dev. 2004, 16. [Google Scholar]
- Yuan, L.; Li, W.; Huo, Q.; Du, C.; Wang, Z.; Yi, B.; Wang, M. Effects of xylo-oligosaccharide and flavomycin on the immune function of broiler chickens. Peerj 2018, 6, e4435. [Google Scholar] [CrossRef]
- Witte, W. Impact of Antibiotic use in Animal Feeding on Resistance of Bacterial Pathogens in Humans. Ciba Found. Symp. 2007, 207, 61–71. [Google Scholar]
- Ateya, A.I.; Arafat, N.; Saleh, R.M.; Ghanem, H.M.; Naguib, D.; Radwan, H.A.; Elseady, Y.Y. Intestinal gene expressions in broiler chickens infected with Escherichia coli and dietary supplemented with probiotic, acidifier and synbiotic. Vet. Res. Commun. 2019, 43, 131–142. [Google Scholar] [CrossRef]
- Lorenzo, C.; de Jesús, J.; Ramirez, A.M.; Muñoz, Y.G. Polimixinas en la era de la multidrogorresistencia. Rev. De Enferm. Infecc. En Pediatría 2011, 24, 66–70. [Google Scholar]
- Sui, O.; Matsumoto, K.; Okubo, M.; Yokoyama, Y.; Kizu, J. Development of HPLC with fluorescent detection using NBD-F for the quantification of colistin sulfate in rat plasma and its pharmacokinetic applications. Biomed. Chromatogr. 2018, 32, e4167. [Google Scholar]
- Dar, M.A.; Urwat, U.; Ahmad, S.M.; Ahmad, R.; Kashoo, Z.A.; Dar, T.A.; Bhat, S.A.; Mumtaz, P.T.; Shabir, N.; Shah, R.A.; et al. Gene expression and antibody response in chicken against Salmonella Typhimurium challenge. Poult. Sci. 2019, 98, 2008–2013. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.J.; Ju, T.T.; Piao, X.S. Effect of Flavomycin on Performance, Gut Morphology and Intestinal Microflora in Broilers. J. Ani. Vet. Adv. 2012, 11, 1669–1673. [Google Scholar] [CrossRef]
- Wang, H.L.; Shi, M.; Xu, X.; Pan, L.; Zhao, P.F.; Ma, X.K.; Tian, Q.Y.; Piao, X.S. Effects of Flavomycin, Bacillus licheniformis and Enramycin on Performance, Nutrient Digestibility, Gut Morphology and the Intestinal Microflora of Broilers. J. Poult. Sci. 2016, 53, 128–135. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods. 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Vicu, A.E.A.; Kuttappan, V.A.; Tellez, G.; Hernandez-Velasco, X.; Seeber-Galarza, R.; Latorre, J.D.; Faulkner, O.B.; Wolfenden, A.D.; Hargis, B.M.; Bielke, L.R. Dose titration of FITC-D for optimal measurement of enteric inflammation in broiler chicks. Poult. Sci. 2015, 94, 1353–1359. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hamid, H.; Zhao, L.H.; Ma, G.Y.; Li, W.X.; Shi, H.Q.; Zhang, J.Y.; Ji, C.; Ma, Q.G. Evaluation of the overall impact of antibiotics growth promoters on broiler health and productivity during the medication and withdrawal period. Poult. Sci. 2019, 107, 1479–1488. [Google Scholar] [CrossRef]
- Tayeri, V.; Seidavi, A.; Asadpour, L.; Phillips, C.J.C. A comparison of the effects of antibiotics, probiotics, synbiotics and prebiotics on the performance and carcass characteristics of broilers. Vet. Res. Commun. 2018, 42, 195–207. [Google Scholar] [CrossRef] [PubMed]
- El-Sharkawy, H.; Tahoun, A.; El-Gohary, A.E.A.; El-Abasy, M.; El-Khayat, F.; Gillespie, T.; Kitade, Y.; Hafez, H.M.; Neubauer, H.; El-Adawy, H. Epidemiological, molecular characterization and antibiotic resistance of Salmonella enterica serovars isolated from chicken farms in Egypt. Gut Pathog. 2017, 9, 1–9. [Google Scholar] [CrossRef]
- Withanage, G.; Kaiser, P.; Wigley, P.; Powers, C.; Mastroeni, P.; Brooks, H.; Barrow, P.; Smith, A.; Maskell, D.; McConnell, I. Rapid expression of chemokines and proinflammatory cytokines in newly hatched chickens infected with Salmonella enterica serovar typhimurium. Infect. Immun. 2004, 72, 2152–2159. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Cheng, Q.; Li, Y.; Duan, R.; Hou, Y.; Yi, D.; Ding, B. Effects of dietary coated-oleum cinnamomi supplementation on the immunity and intestinal integrity of broiler chickens. Anim. Sci. J. 2018, 89, 1581–1590. [Google Scholar] [CrossRef]
- Qiutang, L.; Verma, I.M. NF-kappaB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725. [Google Scholar]
- Vallabhapurapu, S.; Karin, M. Regulation and Function of NF-κB Transcription Factors in the Immune System. Annu. Rev. Immunol. 2009, 27, 693–733. [Google Scholar] [CrossRef]
- Kawai, T.; Adachi, O.; Ogawa, T.; Takeda, K.; Akira, S. Unresponsiveness of MyD88-Deficient Mice to Endotoxin. Immunity 1999, 11, 115–122. [Google Scholar] [CrossRef]
- Asa, H.; Goran, M. Gut Microbiota and Inflammation. Nutrients 2011, 3, 637–682. [Google Scholar]
- Li, J.; Hao, H.; Cheng, G.; Wang, X.; Ahmed, S.; Shabbir, M.A.B.; Liu, Z.; Dai, M.; Yuan, Z. The effects of different enrofloxacin dosages on clinical efficacy and resistance development in chickens experimentally infected with Salmonella Typhimurium. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef]
- Sluis, M.V.D.; De Koning, B.A.E.; De Bruijn, A.C.J.M.; Velcich, A.; Meijerink, J.P.P.; Van Goudoever, J.B.; Büller, H.A.; Dekker, J.; Van Seuningen, I.; Renes, I.B.; et al. Muc2-Deficient Mice Spontaneously Develop Colitis, Indicating That MUC2 Is Critical for Colonic Protection. Gastroenterology 2006, 131, 117–129. [Google Scholar] [CrossRef]
- Barmeyer, C.; Fromm, M.; Schulzke, J. Active and passive involvement of claudins in the pathophysiology of intestinal inflammatory diseases. Pflug. Arch. Eur. J. Phy. 2017, 469, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Guo, Y.; Wang, Z. beta-1,3/1,6-Glucan alleviated intestinal mucosal barrier impairment of broiler chickens challenged with Salmonella enterica serovar Typhimurium. Poult. Sci. 2013, 92, 1764–1773. [Google Scholar] [CrossRef] [PubMed]
- Nicholls, S.J.; Hazen, S.L. Myeloperoxidase and cardiovascular disease. Arter. Thromb Vasc. Biol. 2005, 25, 1102–1111. [Google Scholar] [CrossRef] [PubMed]
- Schürmann, N.; Forrer, P.; Casse, O.; Li, J.; Felmy, B.; Burgener, A.; Ehrenfeuchter, N.; Hardt, W.; Recher, M.; Hess, C.; et al. Myeloperoxidase targets oxidative host attacks to Salmonella and prevents collateral tissue damage. Nat. Microbiol. 2017, 2, 2869–2881. [Google Scholar]
- Islam, M.R.; Lepp, D.; Godfrey, D.V.; Orban, S.; Ross, K.; Delaquis, P.; Diarra, M.S. Effects of wild blueberry (Vaccinium angustifolium) pomace feeding on gut microbiota and blood metabolites in free-range pastured broiler chickens. Poult. Sci. 2019, 98, 3739–3755. [Google Scholar] [CrossRef]
- Choi, J.H.; Lee, K.; Kim, D.W.; Kil, D.Y.; Kim, G.B.; Cha, C.J. Influence of dietary avilamycin on ileal and cecal microbiota in broiler chickens. Poult. Sci. 2018, 97, 970–979. [Google Scholar] [CrossRef]
- Mancabelli, L.; Ferrario, C.; Milani, C.; Mangifesta, M.; Turroni, F.; Duranti, S.; Lugli, G.A.; Viappiani, A.; Ossiprandi, M.C.; van Sinderen, D.; et al. Insights into the biodiversity of the gut microbiota of broiler chickens. Environ. Microbiol. 2016, 18, 4727–4738. [Google Scholar] [CrossRef]
- Kim, S.A.; Rubinelli, P.M.; Park, S.H.; Ricke, S.C. Ability of Arkansas LaKast and LaKast Hybrid Rice Bran to Reduce Salmonella Typhimurium in Chicken Cecal Incubations and Effects on Cecal Microbiota. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Shi, D.; Bai, L.; Qu, Q.; Zhou, S.; Yang, M.; Guo, S.; Li, Q.; Liu, C. Impact of gut microbiota structure in heat-stressed broilers. Poult. Sci. 2019, 98, 2405–2413. [Google Scholar] [CrossRef]
- Pan, D.; Yu, Z. Intestinal microbiome of poultry and its interaction with host and diet. Gut Microbes 2014, 5, 108–119. [Google Scholar] [CrossRef]
- Neal-Mckinney, J.M.; Lu, X.; Duong, T.; Larson, C.L.; Call, D.R.; Shah, D.H.; Konkel, M.E. Production of organic acids by probiotic Lactobacilli can be used to reduce pathogen load in poultry. PLoS ONE 2012, 7, e43928. [Google Scholar] [CrossRef] [PubMed]
- Murry, A.C., Jr.; Hinton, A., Jr. Inhibition of Growth of Escherichia coli, Salmonella typhimurium, and Clostridia perfringens on Chicken Feed Media by Lactobacillus salivarius and Lactobacillus plantarum. Int. J. Poult. Sci. 2004, 3, 101–112. [Google Scholar]
- Zhang, L.; Wu, W.; Lee, Y.; Xie, J.; Zhang, H. Spatial Heterogeneity and Co-occurrence of Mucosal and Luminal Microbiome across Swine Intestinal Tract. Front. Microbiol. 2018, 9, 1730–1741. [Google Scholar] [CrossRef] [PubMed]
- Vogel, H.; Shukla, S.P.; Engl, T.; Weiss, B.; Fischer, R.; Steiger, S.; Heckel, D.G.; Kaltenpoth, M.; Vilcinskas, A. The digestive and defensive basis of carcass utilization by the burying beetle and its microbiota. Nat. Commun. 2017, 8, 15186–15198. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (%) | Content | Chemical Composition | Content |
|---|---|---|---|
| Yellow corn | 56.34 | ME (Kcal/kg) | 2.96 |
| Soybean meal | 33.5 | Crude protein (%) | 21 |
| Corn gluten meal (60%) | 3 | Calcium (%) | 1 |
| Soybean oil | 2.9 | Nonphytate P (%) | 0.45 |
| Limestone | 1.25 | Tryptophan | 0.23 |
| Dicalcium phosphate | 1.9 | Methionine (%) | 0.5 |
| Vitamin premix 1 | 0.02 | Lysine (%) | 1.15 |
| Mineral premix 2 | 0.2 | Threonine (%) | 0.78 |
| Salt | 0.35 | ||
| dl-methionine | 0.19 | ||
| l-lysine Hcl | 0.12 | ||
| Choline chloride | 0.2 | ||
| Ethoxyquin | 0.03 |
| Gene Name | Primer Sequence (5‘–3′) | GenBank |
|---|---|---|
| claudin-1 | F: CATACTCCTGGGTCTGGTTGGT | AY750897.1 |
| R:GACAGCCATCCGCATCTTCT | ||
| occludin | F:ACGGCAGCACCTACCTCAA | D21837.1 |
| R:GGGCGAAGAAGCAGATGAG | ||
| mucin-2 | F:TTCATGATGCCTGCTCTTGTG | XM_421035 |
| R:CCTGAGCCTTGGTACATTCTTGT | ||
| IFN-γ | F:AGCTGACGGTGGACCTATTATT | Y07922 |
| R:GGCTTTGCGCTGGATTC | ||
| TLR2 | F:CTGGGAAGTGGATTGTGGA | AB050005.2 |
| R:AAGGCGAAAGTGCGAGAAA | ||
| TLR4 | F:AGTCTGAAATTGCTGAGCTCAAAT | NM-001030693 |
| R:GCGACGTTAAGCCATGGAAG | ||
| NF-κB | F:GTGTGAAGAAACGGGAACTG | NM-205129 |
| R:GGCACGGTTGTCATAGATGG | ||
| MyD88 | F:CTGGCATCTTCTGAGTAGT | NM-001030962 |
| R:TTCCTTATAGTTCTGGCTTCT | ||
| GAPDH | F:CCTAGGATACACAGAGGACCAGGTT | NM-204305 |
| R:GGTGGAGGAATGGCTGTCA |
| Treatment | 1–14 d | 8–14 d | 1–21 d | 15–21 d | 1–42 d | 1–42 d | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Body Weight Gain (g) | Feed Intake (g) | Feed Conversion Ratio | Mortality (%) | Body Weight Gain(g) | Feed Intake(g) | Feed Conversion Ratio | Mortality (%) | Body Weight Gain (g) | Feed Intake (g) | Feed Conversion Ratio | Mortality (%) | ||
| CON | 242.98 | 333.47 | 1.39 | 0 | 533.26 | 879.29 | 1.63 ab | 0 | 2436.8 | 5145.4 | 2.09 a | 0 | |
| Anti G+ | 220.24 | 321.9 | 1.42 | 0 | 499.96 | 844.23 | 1.68 ab | 0 | 2431.8 | 5293.3 | 2.2 b | 0 | |
| Anti G− | 244.29 | 346.33 | 1.4 | 0 | 549.74 | 893.06 | 1.59 a | 0 | 2412.5 | 5321.0 | 2.24 b | 0 | |
| CONS | 222.3 | 321.77 | 1.47 | 0 | 483.93 | 808.05 | 1.72 b | 3.03 | 2365.9 | 5093.8 | 2.24 b | 3.85 | |
| Anti G+S | 222.96 | 319.23 | 1.43 | 1.52 | 490.47 | 804.4 | 1.65 ab | 0 | 2324.3 | 5281.5 | 2.22 b | 2.56 | |
| Anti G−S | 239.52 | 337.05 | 1.44 | 0 | 511.35 | 865.56 | 1.68 ab | 0 | 2386.9 | 5045.5 | 2.15 ab | 0 | |
| SEM | 3.14 | 3.18 | 0.01 | 0 | 6.39 | 11.37 | 0.01 | 0 | 19.38 | 37.37 | 0.02 | 0.01 | |
| Main effect | |||||||||||||
| Bacterial community | CON | 232.64 ab | 327.62 ab | 1.43 | 0 | 508.59 ab | 843.67 | 1.68 | 1.52 | 2401.4 | 5119.6 | 2.17 | 1.93 |
| Anti G+ | 221.60 a | 320.56 a | 1.43 | 0.76 | 495.22 a | 824.31 | 1.67 | 0 | 2378.0 | 5287.4 | 2.21 | 1.28 | |
| Anti G− | 241.90 b | 341.69 b | 1.42 | 0 | 530.55 b | 879.31 | 1.64 | 0 | 2399.7 | 5183.2 | 2.20 | 0 | |
| S. typhimurium | − | 235.32 | 334.76 | 1.4 | 0 | 527.25 | 873.8 | 1.64 | 0 | 2426.9 | 5259.5 | 2.18 | 0 |
| + | 227.89 | 326.7 | 1.45 | 0.01 | 495.59 | 822.17 | 1.69 | 0.01 | 2361.5 | 5155.9 | 2.20 | 2.14 | |
| p value | |||||||||||||
| Feed | 0.02 | 0.019 | 0.902 | 0.38 | 0.027 | 0.114 | 0.253 | 0.099 | 0.895 | 0.166 | 0.504 | 0.436 | |
| S. typhimurium | 0.182 | 0.184 | 0.005 | 0.325 | 0.004 | 0.035 | 0.012 | 0.124 | 0.114 | 0.156 | 0.435 | 0.091 | |
| Bacterial community * S. typhimurium | 0.233 | 0.815 | 0.135 | 0.38 | 0.281 | 0.668 | 0.02 | 0.099 | 0.711 | 0.295 | 0.007 | 0.436 | |
| Treatment | 14 d | 21 d | |||||
|---|---|---|---|---|---|---|---|
| Villus Height (μm) | Crypt Depth (μm) | V/C | Villus Height (μm) | Crypt Depth (μm) | V/C | ||
| CON | 561.84 | 155.89 | 3.67 | 600.61 | 166.34 | 3.66 | |
| Anti G+ | 544.6 | 159.13 | 3.51 | 571.76 | 160.8 | 3.64 | |
| Anti G− | 557.2 | 159.86 | 3.52 | 571.92 | 160.36 | 3.52 | |
| CONS | 554.1 | 164.33 | 3.49 | 551.91 | 157.29 | 3.42 | |
| Anti G+S | 548.22 | 157.3 | 3.47 | 554.59 | 163.61 | 3.47 | |
| Anti G−S | 524.91 | 157.15 | 3.44 | 592.39 | 176.23 | 3.42 | |
| SEM | 5.16 | 1.61 | 0.02 | 6.37 | 2.32 | 0.03 | |
| Main effect | |||||||
| Bacterial community | CON | 557.97 | 157.52 | 3.58 a | 570.95 | 161.81 | 3.54 |
| Anti G+ | 550.86 | 158.22 | 3.49 ab | 563.18 | 161.04 | 3.54 | |
| Anti G− | 543.01 | 155.36 | 3.48 b | 582.15 | 169.5 | 3.48 | |
| S. typhimurium | − | 554.14 | 158.08 | 3.57 | 580.69 | 162.13 | 3.61 |
| + | 541.57 | 160.08 | 3.47 | 566.30 | 166.31 | 3.44 | |
| p value | |||||||
| Feed | 0.421 | 0.875 | 0.025 | 0.418 | 0.421 | 0.358 | |
| S. typhimurium | 0.253 | 0.702 | 0.002 | 0.219 | 0.482 | 0.003 | |
| Bacterial community * S. typhimurium | 0.381 | 0.305 | 0.184 | 0.090 | 0.109 | 0.494 | |
| Treatment | claudin-1 | occludin | mucin-2 | IFN-γ | TLR2 | TLR4 | NF-κB | MyD88 | |
|---|---|---|---|---|---|---|---|---|---|
| CON | 1.03 | 1.02 | 1.02 ab | 1.02 | 1.04 | 1.02 | 0.9 | 1.01 | |
| Anti G+ | 0.8 | 0.84 | 0.99 ab | 1.25 | 1.01 | 1.35 | 0.85 | 0.86 | |
| Anti G− | 0.87 | 1.11 | 0.89 a | 1.24 | 0.87 | 1.18 | 1.06 | 0.94 | |
| CONS | 0.51 | 0.77 | 1.67 c | 1.21 | 1.06 | 1.26 | 1.69 | 1.58 | |
| Anti G+S | 0.43 | 0.75 | 2.12 d | 1.52 | 1.39 | 1.26 | 1.91 | 1.59 | |
| Anti G−S | 0.62 | 0.79 | 1.35 bc | 1.28 | 0.98 | 1.24 | 1.95 | 1.71 | |
| SEM | 0.05 | 0.03 | 0.09 | 0.04 | 0.05 | 0.04 | 0.10 | 0.07 | |
| Main effect | |||||||||
| Bacterial community | CON | 0.77 | 0.89 | 1.34 ab | 1.11 a | 1.05 | 1.14 | 1.33 b | 1.24 |
| Anti G+ | 0.62 | 0.79 | 1.55 a | 1.38 b | 1.2 | 1.3 | 1.39 ab | 1.18 | |
| Anti G− | 0.74 | 0.95 | 1.12 b | 1.26 ab | 0.92 | 1.21 | 1.55 a | 1.32 | |
| S. typhimurium | − | 0.91 | 0.98 | 0.97 | 1.15 | 0.98 | 1.17 | 0.95 | 0.94 |
| + | 0.53 | 0.77 | 1.69 | 1.32 | 1.13 | 1.25 | 1.86 | 1.62 | |
| p value | |||||||||
| Feed | 0.203 | 0.060 | <0.001 | 0.024 | 0.055 | 0.182 | 0.040 | 0.469 | |
| S. typhimurium | <0.001 | 0.001 | <0.001 | 0.035 | 0.058 | 0.342 | <0.001 | <0.001 | |
| Bacterial community * S. typhimurium | 0.268 | 0.164 | 0.004 | 0.499 | 0.222 | 0.194 | 0.281 | 0.393 | |
| Treatment | claudin-1 | occludin | mucin-2 | IFN-γ | TLR2 | TLR4 | NF-κB | MyD88 | |
|---|---|---|---|---|---|---|---|---|---|
| CON | 1.04 | 1.02 | 1.02 a | 1.04 a | 1.03 ab | 1.01 | 1.01 | 1.02 | |
| Anti G+ | 1.12 | 0.93 | 1.20 a | 1.37 ab | 0.88 a | 1.24 | 1.33 | 1.27 | |
| Anti G− | 0.89 | 0.94 | 1.28 a | 1.26 a | 0.95 a | 1.1 | 1.09 | 1.19 | |
| CONS | 0.71 | 0.69 | 1.01 a | 1.74 b | 1.08 ab | 1.32 | 0.98 | 0.96 | |
| Anti G+S | 0.52 | 0.78 | 1.74 b | 1.80 b | 1.32 a | 1.54 | 1.15 | 1.27 | |
| Anti G−S | 0.64 | 0.93 | 1.40 ab | 1.32 ab | 1.15 ab | 1.5 | 1.1 | 1.06 | |
| SEM | 0.05 | 0.04 | 0.06 | 0.06 | 0.04 | 0.05 | 0.03 | 0.03 | |
| Main effect | |||||||||
| Bacterial community | CON | 0.88 | 0.85 | 1.01 a | 1.39 ab | 1.06 | 1.17 | 1.00 a | 0.99 a |
| Anti G+ | 0.82 | 0.85 | 1.47 b | 1.59 a | 1.1 | 1.39 | 1.24 b | 1.27 b | |
| Anti G− | 0.77 | 0.93 | 1.34 b | 1.29 b | 1.05 | 1.3 | 1.11 ab | 1.12 ab | |
| S. typhimurium | − | 1.02 | 0.96 | 1.16 | 1.22 | 0.95 | 1.11 | 1.14 | 1.15 |
| + | 0.63 | 0.81 | 1.35 | 1.62 | 1.18 | 1.46 | 1.08 | 1.10 | |
| p value | |||||||||
| Feed | 0.448 | 0.481 | <0.001 | 0.026 | 0.742 | 0.054 | 0.009 | <0.001 | |
| S. typhimurium | <0.001 | 0.017 | 0.008 | <0.001 | 0.001 | <0.001 | 0.299 | 0.256 | |
| Bacterial community * S. typhimurium | 0.137 | 0.135 | 0.018 | 0.012 | 0.045 | 0.841 | 0.426 | 0.568 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Yang, Y.; Dong, Y.; Yan, C.; Zhang, B. The Effects of Flavomycin and Colistin Sulfate Pre-Treatment on Ileal Bacterial Community Composition, the Response to Salmonella typhimurium and Host Gene Expression in Broiler Chickens. Microorganisms 2019, 7, 574. https://doi.org/10.3390/microorganisms7110574
He Y, Yang Y, Dong Y, Yan C, Zhang B. The Effects of Flavomycin and Colistin Sulfate Pre-Treatment on Ileal Bacterial Community Composition, the Response to Salmonella typhimurium and Host Gene Expression in Broiler Chickens. Microorganisms. 2019; 7(11):574. https://doi.org/10.3390/microorganisms7110574
Chicago/Turabian StyleHe, Yang, Yanyan Yang, Yuanyang Dong, Changliang Yan, and Bingkun Zhang. 2019. "The Effects of Flavomycin and Colistin Sulfate Pre-Treatment on Ileal Bacterial Community Composition, the Response to Salmonella typhimurium and Host Gene Expression in Broiler Chickens" Microorganisms 7, no. 11: 574. https://doi.org/10.3390/microorganisms7110574
APA StyleHe, Y., Yang, Y., Dong, Y., Yan, C., & Zhang, B. (2019). The Effects of Flavomycin and Colistin Sulfate Pre-Treatment on Ileal Bacterial Community Composition, the Response to Salmonella typhimurium and Host Gene Expression in Broiler Chickens. Microorganisms, 7(11), 574. https://doi.org/10.3390/microorganisms7110574