Genetic Characterization and Pathological Analysis of a Novel Bacterial Pathogen, Pseudomonas tructae, in Rainbow Trout (Oncorhynchus mykiss)

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Sampling and Postmortem Examination
2.2. Bacterial Isolation
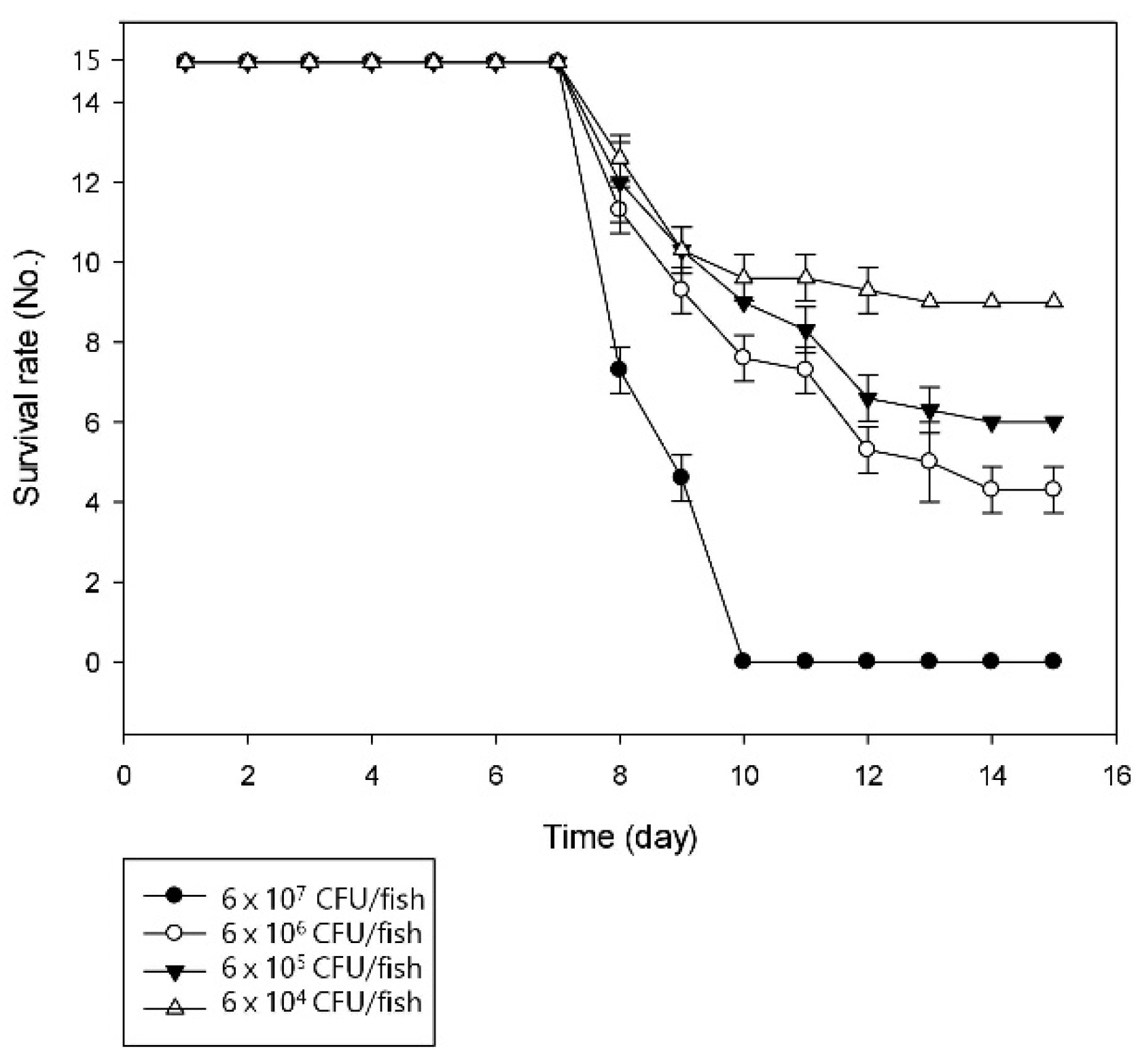
2.3. Challenge Trial
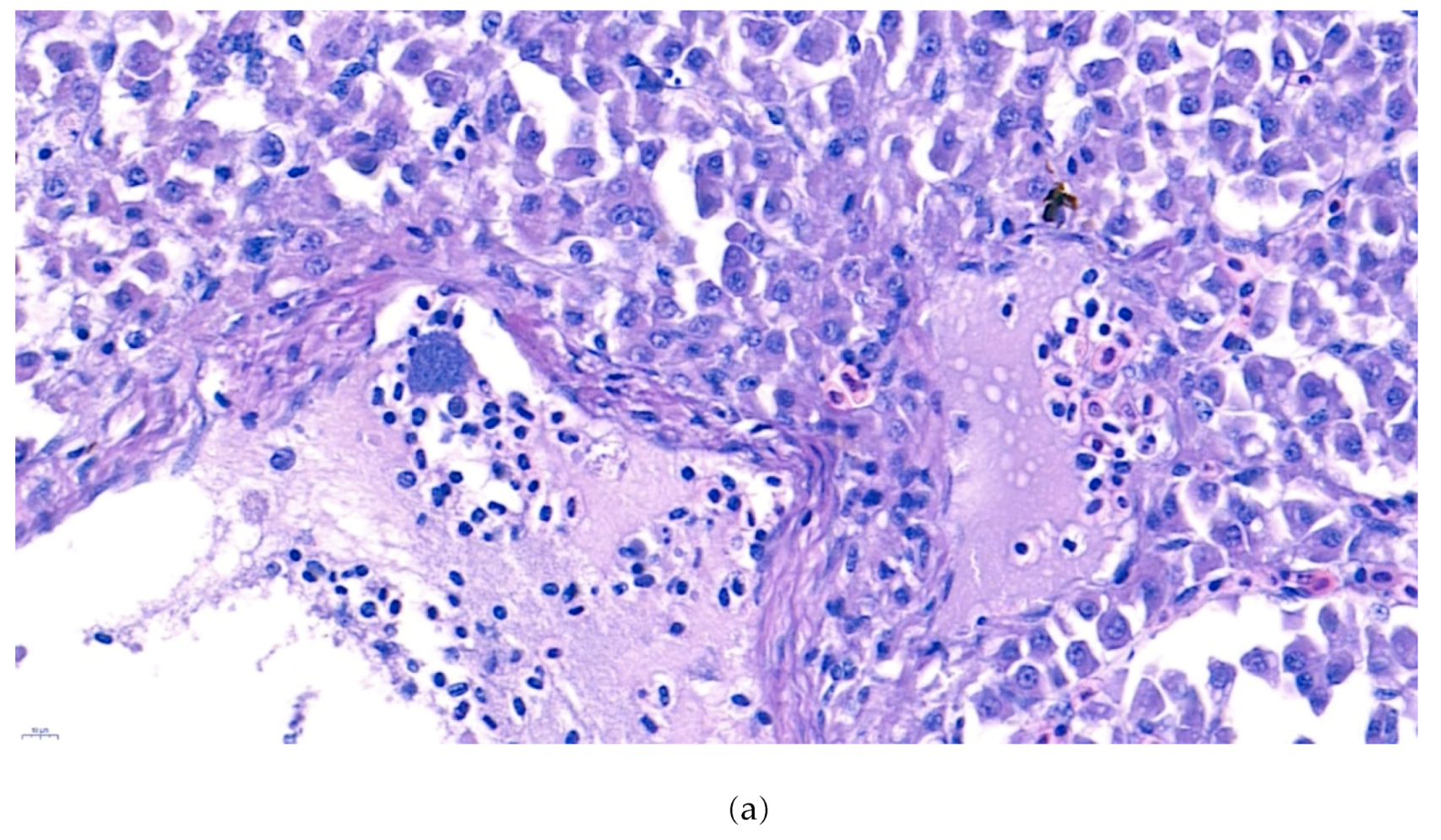
2.4. Histopathological Analysis
2.5. Antibiotic Susceptibility Test
2.6. Genome Characterization
3. Results and Discussion
3.1. Postmortem Examination of the Diseased Fish
3.2. Pathogenicity of the Bacterial Strain
3.3. Histopathological Analysis
3.4. Antibiotic Susceptibility Result
3.5. Genome Features of Strain SNU WT1
3.6. General Information about Strain SNU WT1 in This Study
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cross, A.; Allen, J.R.; Burke, J.; Ducel, G.; Harris, A.; John, J.; Skalova, R. Nosocomial Infections Due to Pseudomonas Aeruginosa: Review of Recent Trends. Rev. Infect. Dis. 1983, 5, S837–S845. [Google Scholar] [CrossRef] [PubMed]
- Gram, L.O.N.E. Inhibitory Effect Against Pathogenic and Spoilage Bacteria of Pseudomonas Strains Isolated from Spoiled and Fresh Fish. Appl. Environ. Microbiol. 1993, 59, 2197–2203. [Google Scholar] [PubMed]
- Dogan, B.; Boor, K.J. Genetic Diversity and Spoilage Potentials among Pseudomonas Spp. Isolated from Fluid Milk Products and Dairy Processing Plants. Appl. Environ. Microbiol. 2003, 69, 130–138. [Google Scholar] [CrossRef] [PubMed]
- Yumoto, I.; Yamazaki, K.; Hishinuma, M.; Nodasaka, Y.; Suemori, A.; Nakajima, K.; Kawasaki, K. Pseudomonas Alcaliphila sp. nov., a novel facultatively Psychrophilic alkaliphile Isolated from seawater. Int. J. Syst. Evol. Microbiol. 2001, 51, 349–355. [Google Scholar] [CrossRef] [PubMed]
- Frasson, D.; Opoku, M.; Picozzi, T.; Torossi, T.; Balada, S.; Smits, T.H.; Hilber, U. Pseudomonas Wadenswilerensis sp. nov. and Pseudomonas Reidholzensis sp. nov., two novel species within the Pseudomonas putida group isolated from forest soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 2853–2861. [Google Scholar] [CrossRef] [PubMed]
- Kominos, S.D.; Copeland, C.E.; Grosiak, B.; Postic, B. Introduction of Pseudomonas Aeruginosa into A Hospital Via Vegetables. Appl. Environ. Microbiol. 1972, 24, 567–570. [Google Scholar] [CrossRef]
- Buell, C.R.; Joardar, V.; Lindeberg, M.; Selengut, J.; Paulsen, I.T.; Gwinn, M.L.; Madupu, R. The Complete Genome Sequence of the Arabidopsis and Tomato Pathogen Pseudomonas Syringae pv. tomato DC3000. Proc. Natl. Acad. Sci. USA 2003, 100, 10181–10186. [Google Scholar] [CrossRef]
- Stover, C.K.; Pham, X.Q.; Erwin, A.L.; Mizoguchi, S.D.; Warrener, P.; Hickey, M.J.; Garber, R.L. Complete Genome Sequence of Pseudomonas Aeruginosa PAO1, an opportunistic pathogen. Nature 2000, 406, 959. [Google Scholar] [CrossRef]
- Lambert, P.A. Mechanisms of antibiotic resistance in Pseudomonas aeruginosa. J. R. Soc. Med. 2002, 95, 22. [Google Scholar]
- Altinok, I.; Kayis, S.; Capkin, E. Pseudomonas putida infection in rainbow trout. Aquaculture 2006, 261, 850–855. [Google Scholar] [CrossRef]
- Gilardi, G.L. Infrequently Encountered Pseudomonas Species Causing Infection in Humans. Ann. Intern. Med. 1972, 77, 211–215. [Google Scholar] [CrossRef]
- De Kievit, T.R.; Parkins, M.D.; Gillis, R.J.; Srikumar, R.; Ceri, H.; Poole, K.; Storey, D.G. Multidrug Efflux Pumps: Expression Patterns and Contribution to Antibiotic Resistance in Pseudomonas Aeruginosa Biofilms. Antimicrob. Agents Chemother. 2001, 45, 1761–1770. [Google Scholar] [CrossRef] [PubMed]
- Lejbkowicz, F.; Belavsky, L.; Kudinsky, R.; Gery, R. Bacteraemia and Sinusitis Due to Flavimonas Oryzihabitans Infection. Scand. J. Infect. Dis. 2003, 35, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Nishimori, E.; Kita-Tsukamoto, K.; Wakabayashi, H. Pseudomonas plecoglossicida sp. nov., the causative agent of bacterial haemorrhagic ascites of ayu, Plecoglossus altivelis. Int. J. Syst. Evol. Microbial. 2000, 50, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Wakabayashi, H.; Sawada, K.; Ninomiya, K.; Nishimori, E. Bacterial Hemorrhagic Ascites of Ayu Caused by Pseudomonas sp. Fish Pathol. 1996, 31, 239–240. [Google Scholar] [CrossRef][Green Version]
- Kusuda, R.; Toyoshima, T. Characteristics of A Pathogenic Pseudomonas Isolated from Cultured Yellowtail. Fish Pathol. 1976, 11, 133–139. [Google Scholar] [CrossRef]
- Berthe, F.C.; Michel, C.; Bernardet, J.F. Identification of Pseudomonas Anguilliseptica Isolated from Several Fish Species in France. Dis. Aquat. Org. 1995, 21, 151–155. [Google Scholar] [CrossRef]
- Miyazaki, T.; Kubota, S.S.; Miyashita, T.A. Histopathological Study of Pseudomonas Fluorescens Infection in Tilapia. Fish Pathol. 1984, 19, 161–166. [Google Scholar] [CrossRef]
- Eissa, N.M.E.; El-Ghiet, E.A.; Shaheen, A.A.; Abbass, A. Characterization of Pseudomonas Species Isolated from Tilapia “Oreochromis niloticus” in Qaroun and Wadi-El-Rayan lakes, Egypt. Glob. Vet. 2010, 5, 116–121. [Google Scholar]
- Sakai, M.; Atsuta, S.; Kobayashi, M. Pseudomonas Fluorescens Isolated from the Diseased Rainbow Trout, Oncorhynchus Mykiss. Kistato Arch. Exp. Med. 1989, 62, 157–162. [Google Scholar]
- Oh, W.T.; Jun, J.W.; Giri, S.S.; Yun, S.K.; Kim, H.J.; Kim, S.W.; Kang, J.W.; Han, S.J.; Kwon, J.; Park, S.C. Pseudomonas Tructae sp. nov., novel specific species isolated from rainbow trout kidney. Int. J. Syst. Evol. Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.; Rodrıguez, S.; Prieto, S.I.P. Nested PCR Improves Detection of Infectious Hematopoietic Necrosis Virus in Cells Coinfected with Infectious Pancreatic Necrosis Virus. J. Virol. Methods. 1999, 81, 1–9. [Google Scholar] [CrossRef]
- Garver, K.A.; Hawley, L.M.; McClure, C.A.; Schroeder, T.; Aldous, S.; Doig, F.; Richard, J. Development and Validation of A Reverse Transcription Quantitative PCR for Universal Detection of Viral Hemorrhagic Septicemia Virus. Dis. Aquat. Organ. 2011, 95, 97–112. [Google Scholar] [CrossRef] [PubMed]
- Dudley, M.N.; Ambrose, P.G.; Bhavnani, S.M.; Craig, W.A.; Ferraro, M.J.; Jones, R.N. Background and Rationale for Revised Clinical and Laboratory Standards Institute Interpretive Criteria (breakpoints) for Enterobacteriaceae and Pseudomonas Aeruginosa: I. Cephalosporins and Aztreonam. Clin. Infect. Dis. 2013, 56, 1301–1309. [Google Scholar] [PubMed]
- Poole, K.; Krebes, K.; McNally, C.; Neshat, S.H.A.D.I. Multiple Antibiotic Resistance in Pseudomonas Aeruginosa: Evidence for Involvement of An Efflux Operon. J. Bacteriol. 1993, 175, 7363–7372. [Google Scholar] [CrossRef] [PubMed]
- Novotny, L.; Dvorska, L.; Lorencova, A.; Beran, V.; Pavlik, I. Fish: A Potential Source of Bacterial Pathogens for Human Beings. A review. Vet. Med. UZPI (Czech Repub.) 2004, 49, 343–358. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Name | Results of Disk Diffusion Assays | Inhibition Zone Diameter (mm) | Results of MIC Assays | MIC (µg/mL) |
|---|---|---|---|---|
| Amoxicillin-clavulanate | R | 0 | R | 256 |
| Ampicillin-sulbactam | R | 0 | − | − |
| Piperacillin-tazobactam | R | 26 | − | − |
| Cefazolin | R | 0 | − | − |
| Cefepime | R | 28 | − | − |
| Cefotaxime | R | 12 | R | 32 |
| Aztreonam | R | 10 | − | − |
| Cefoxitin | R | 0 | − | − |
| Ceftazidime | I | 20 | − | − |
| Imipenem | R | 22 | R | 4 |
| Meropenem | R | 18 | R | 8 |
| Amikacin | R | 24 | R | 64 |
| Tetracycline | S | 24 | S | 4 |
| Ciprofloxacin | S | 42 | S | 0.06 |
| Levofloxacin | R | 32 | I | 4 |
| Trimethoprim-sulfamethoxazole | I | 16 | − | − |
| Chloramphenicol | R | 0 | − | − |
| Ampicillin | R | 6 | R | 256 |
| Amoxycillin | R | 10 | R | 256 |
| Gentamycin | R | 9 | R | 16 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, W.T.; Kim, J.H.; Jun, J.W.; Giri, S.S.; Yun, S.; Kim, H.J.; Kim, S.G.; Kim, S.W.; Han, S.J.; Kwon, J.; et al. Genetic Characterization and Pathological Analysis of a Novel Bacterial Pathogen, Pseudomonas tructae, in Rainbow Trout (Oncorhynchus mykiss). Microorganisms 2019, 7, 432. https://doi.org/10.3390/microorganisms7100432
Oh WT, Kim JH, Jun JW, Giri SS, Yun S, Kim HJ, Kim SG, Kim SW, Han SJ, Kwon J, et al. Genetic Characterization and Pathological Analysis of a Novel Bacterial Pathogen, Pseudomonas tructae, in Rainbow Trout (Oncorhynchus mykiss). Microorganisms. 2019; 7(10):432. https://doi.org/10.3390/microorganisms7100432
Chicago/Turabian StyleOh, Woo Taek, Ji Hyung Kim, Jin Woo Jun, Sib Sankar Giri, Saekil Yun, Hyoun Joong Kim, Sang Guen Kim, Sang Wha Kim, Se Jin Han, Jun Kwon, and et al. 2019. "Genetic Characterization and Pathological Analysis of a Novel Bacterial Pathogen, Pseudomonas tructae, in Rainbow Trout (Oncorhynchus mykiss)" Microorganisms 7, no. 10: 432. https://doi.org/10.3390/microorganisms7100432
APA StyleOh, W. T., Kim, J. H., Jun, J. W., Giri, S. S., Yun, S., Kim, H. J., Kim, S. G., Kim, S. W., Han, S. J., Kwon, J., & Park, S. C. (2019). Genetic Characterization and Pathological Analysis of a Novel Bacterial Pathogen, Pseudomonas tructae, in Rainbow Trout (Oncorhynchus mykiss). Microorganisms, 7(10), 432. https://doi.org/10.3390/microorganisms7100432