Peroxynitrite Generation and Increased Heterotrophic Capacity Are Linked to the Disruption of the Coral–Dinoflagellate Symbiosis in a Scleractinian and Hydrocoral Species

 ,
,  , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
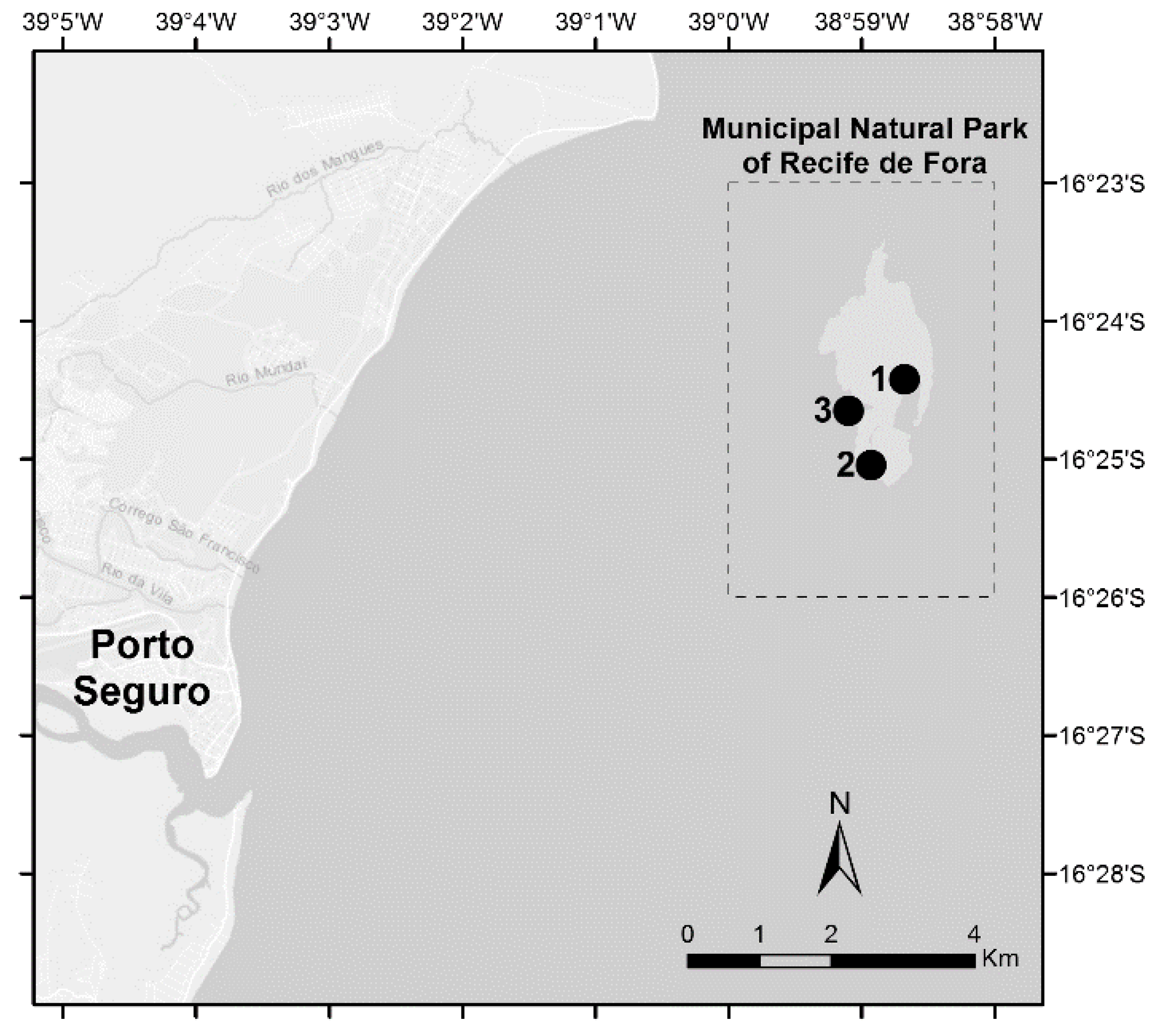
2.1. Reef Sites
2.2. Thermal Stress Assessment
2.3. Biological Survey
2.4. Physiological Measurements
2.4.1. Fatty Acids Extraction and Analysis
2.4.2. Predominant Trophic Mode Index
2.4.3. Peroxynitrite (ONOO−) Quantification
2.5. Data Presentation and Statistical Analyses
3. Results
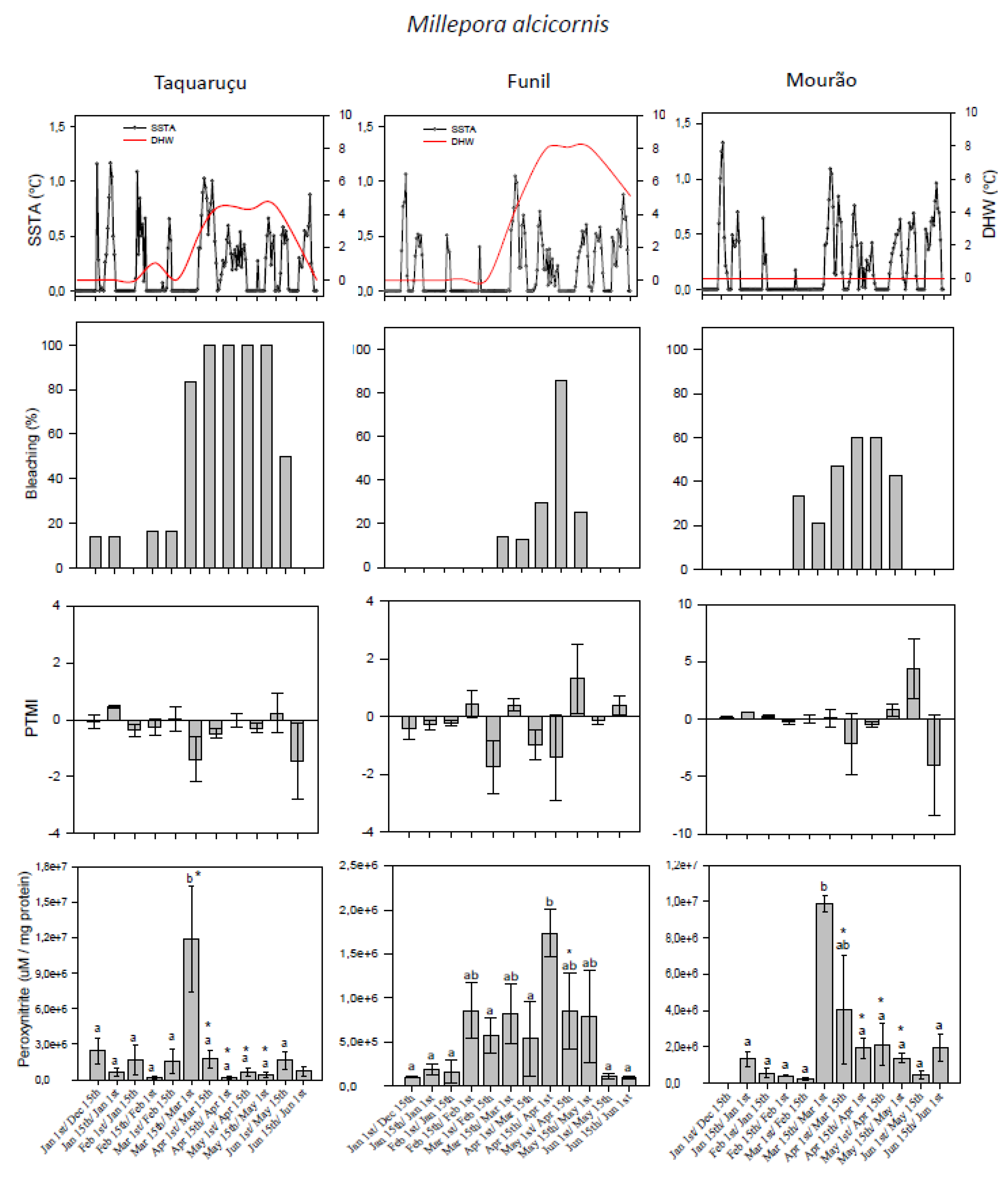
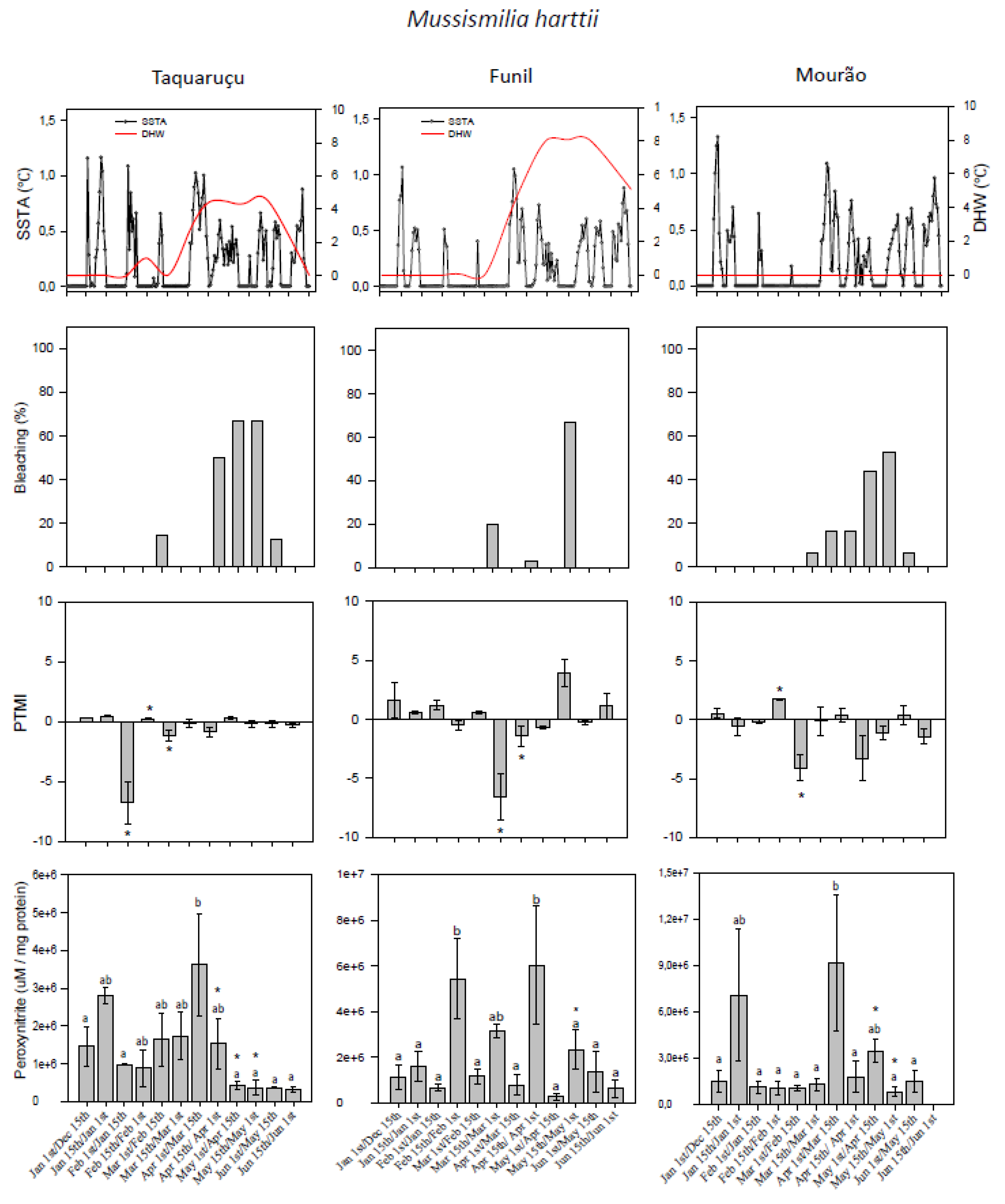
3.1. Thermal Stress and Bleaching Frequency
3.2. Predominant Trophic Mode Index
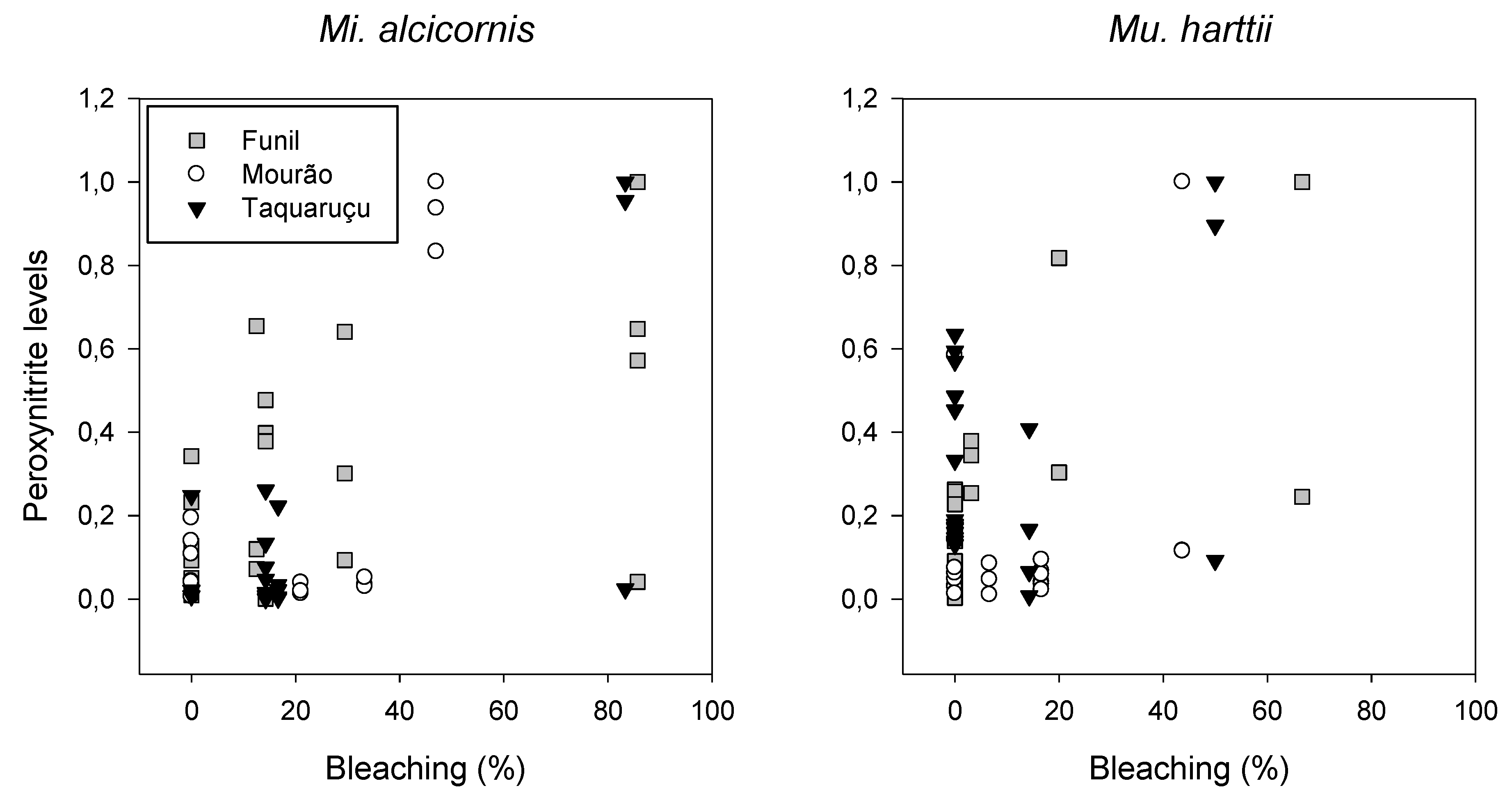
3.3. Peroxynitrite (ONOO−) Generation
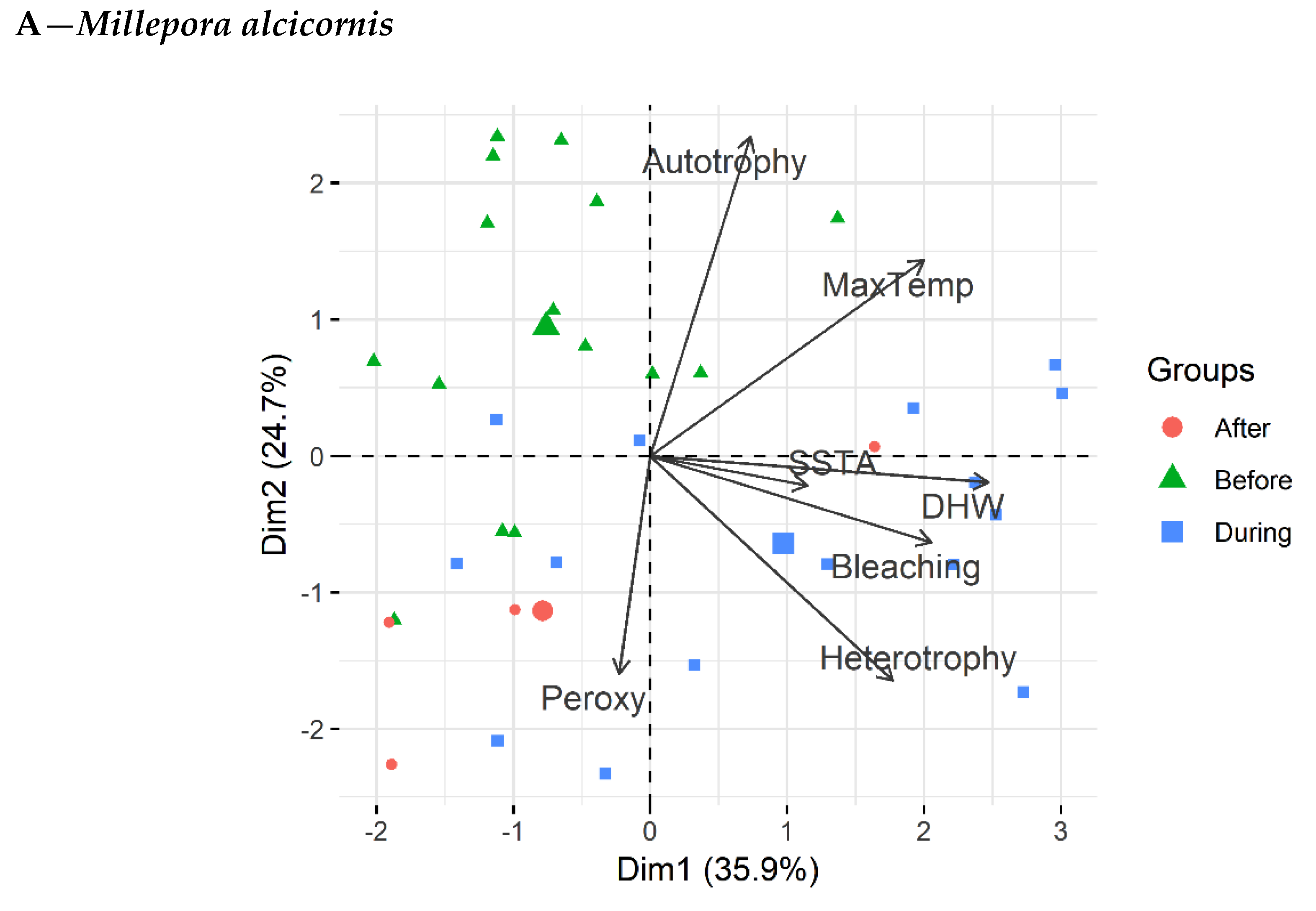
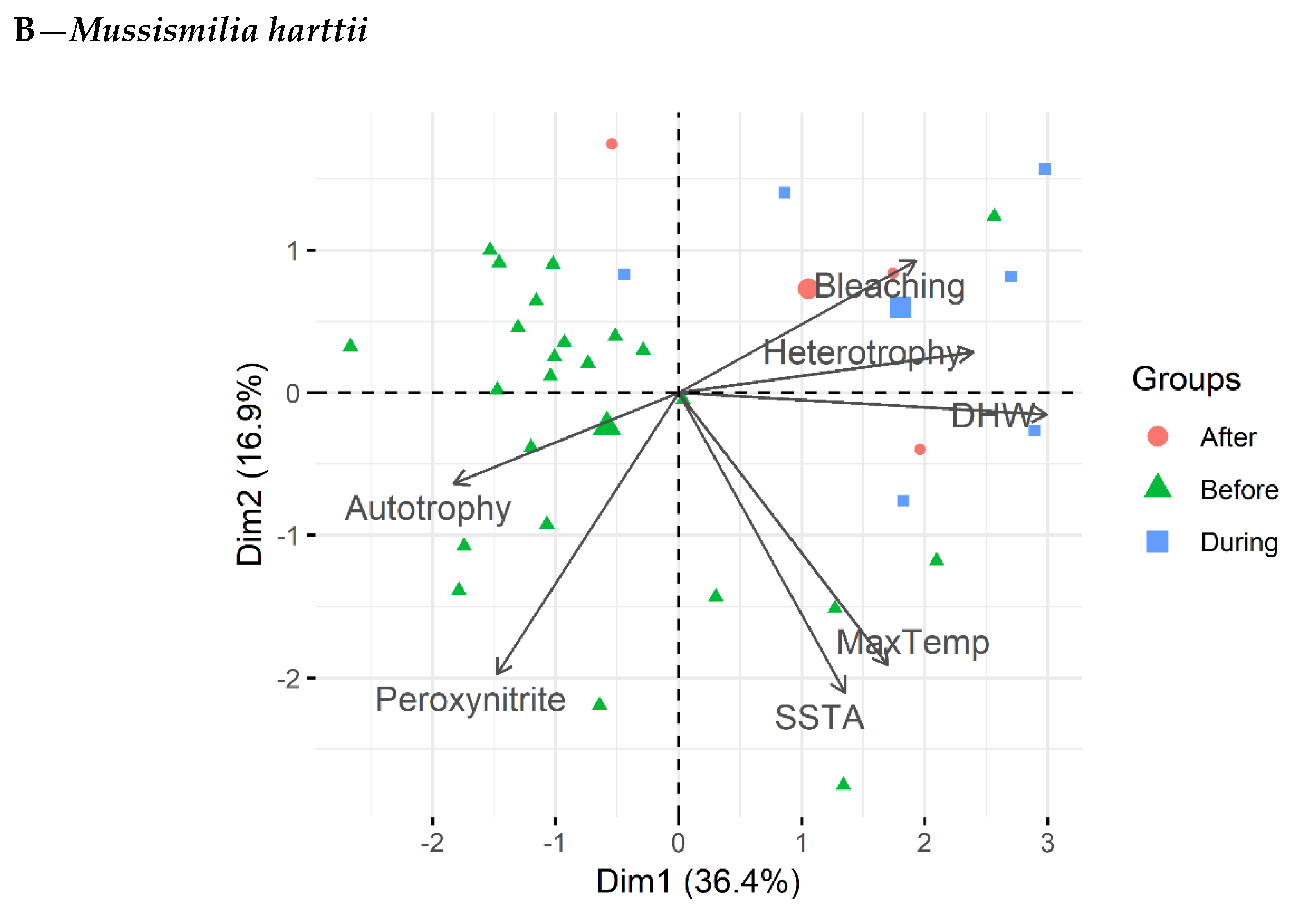
3.4. Principal Component Analysis
4. Discussion
4.1. Thermal Stress, Bleaching Frequency, and Feeding Mode Plasticity
4.2. Peroxynitrite Generation and Bleaching
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C.; et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science. 2018, 359, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Claar, D.C.; Szostek, L.; McDevitt-Irwin, J.M.; Schanze, J.J.; Baum, J.K. Global patterns and impacts of El Niño events on coral reefs: A meta-analysis. PLoS ONE 2018, 13, e0190957. [Google Scholar] [CrossRef]
- Lough, J.M.; Anderson, K.D.; Ughes, T.P. Increasing thermal stress for tropical coral reefs: 1871-2017. Sci. Rep. 2018, 8, 6079. [Google Scholar] [CrossRef] [PubMed]
- Downs, C.A.; Mueller, E.; Phillips, S.; Fauth, J.E.; Woodley, C.M. A molecular biomarker system for assessing the health of coral (Montastrea faveolata) during heat stress. Mar. Biotechnol. 2000, 2, 533–544. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic revision of Symbiodiniaceae highlights the antiquity and diversity of coral endosymbionts. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Muscatine, L.; Porter, J.W. Reef corals: mutualistic symbioses adapted to nutrient-poor environments. Bio. Sci. 1997, 27, 454–460. [Google Scholar] [CrossRef]
- Muscatine, L. The role of symbiotic algae in carbon and energy flux in coral reefs. In Ecosystems of the World; Dubinsky, Z., Ed.; Elsevier: Amsterdam, The Netherlands, 1990; pp. 75–87. [Google Scholar]
- Grottoli, A.G.; Rodrigues, L.J.; Palardy, J.E. Heterotrophic plasticity and resilience in bleached corals. Nature 2006, 440, 1186–1189. [Google Scholar] [CrossRef] [PubMed]
- Hughes, A.D.; Grottoli, A.G. Heterotrophic compensation: a possible mechanism for resilience of coral reefs to global warming or a sign of prolonged stress? PLoS ONE 2012, 8, e81172. [Google Scholar] [CrossRef]
- Mies, M.; Guth, A.Z.; Tenório, A.A.; Banha, T.N.S.; Waters, L.G.; Polito, P.S.; Taniguchi, S.; Bícego, M.C.; Sumida, P.Y.G. In situ shifts of predominance between autotrophic and heterotrophic feeding in the reef-building coral Mussismilia hispida: an approach using fatty acid trophic markers. Coral Reefs 2018, 37, 677–689. [Google Scholar] [CrossRef]
- Lesser, M.P. Elevated temperatures and ultraviolet radiation cause oxidative stress and inhibit photosynthesis in symbiotic dinoflagellates. Limnol. Oceanogr. 1996, 41, 271–283. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative stress causes coral bleaching during exposure to elevated temperatures. Coral Reefs 1997, 16, 187–192. [Google Scholar] [CrossRef]
- Downs, C.A.; Fauth, J.E.; Halas, J.C.; Dustan, P.; Bemiss, J.; Woodley, C.M. Oxidative stress and seasonal coral bleaching. Free Rad Biol. Med. 2002, 33, 533–543. [Google Scholar] [CrossRef]
- Pyper, B.J.; Peterman, R.M. Comparison of methods to account for autocorrelation in correlation analyses of fish data. Can. J. Fish Aquat. Sci. 1998, 55, 2127–2140. [Google Scholar] [CrossRef]
- Wiedenmann, J.; D’Angelo, C.; Smith, E.G.; Hunt, A.N.; Legiret, F.E.; Postle, A.D.; Achterberg, E.P. Nutrient enrichment can increase the susceptibility of reef corals to bleaching. Nat. Clim. Change 2013, 3, 160–164. [Google Scholar] [CrossRef]
- Hawkins, T.D.; Davy, S.K. Nitric oxide and coral bleaching: is peroxynitrite generation required for symbiosis collapse. J. Exp. Biol. 2013, 216, 3185–3188. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, T.D.; Bradley, B.J.; Davy, S.K. Nitric oxide mediates coral bleaching through an apoptotic-like cell death pathway: evidence from a model sea anemone-dinoflagellate symbiosis. FASEB J. 2013, 27, 4790–4798. [Google Scholar] [CrossRef]
- Sebens, K.P.; Koehl, M.A.R. Predation on zooplankton by the benthic anthozoans Alcyonium siderium (Alcyonacea) and Metricium senile (Actiniaria) in the New England subtidal. Mar. Biol. 1984, 81, 255–271. [Google Scholar] [CrossRef]
- Nielsen, D.A.; Petrou, K.; Gates, R.D. Coral bleaching from a single cell perspective. ISME J. 2018, 12, 1558–15675. [Google Scholar] [CrossRef] [PubMed]
- Lesser, M.P. Oxidative stress in marine environments: biochemistry and physiological Ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef]
- Teixeira, C.D.; Leitão, R.L.L.; Ribeiro, F.V.; Moraes, F.C.; Neves, L.M.; Bastos, A.C.; Pereira-Filho, G.H.; Kampel, M.; Salomon, P.S.; Sá, J.A.; et al. Sustained mass coral bleaching (2016-2017) in Brazilian turbid-zone reefs: taxonomic, cross-shelf and habitat-related trends. Coral Reefs 2019, 38, 801–813. [Google Scholar] [CrossRef]
- Tremblay, P.; Gori, A.; Maguer, J.F.; Hoogenboom, M.; Ferrier-Pagès, C. Heterotrophy promotes the re-establishment of photosynthate translocation in a symbiotic coral after heat stress. Sci. Rep. 2016, 6, 38112. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, T.D.; Davy, S.K. Nitric oxide production and tolerance differ among Symbiodinium types exposed to heat stress. Plant Cell Physiol. 2012, 53, 1889–1898. [Google Scholar] [CrossRef] [PubMed]
- Szabó, C.; Ischiropoulos, H.; Radi, R. Peroxynitrite: biochemistry, pathophysiology and development of therapeutics. Nature 2007, 6, 662–680. [Google Scholar] [CrossRef] [PubMed]
- Maynard, J.A.; Turner, P.T.; Anthony, K.R.N.; Baird, A.H.; Berkelmans, R.; Eakin, C.M.; Johnson, J.; Marshall, P.A.; Packer, G.R.; Rea, A.; et al. TeefTemp: An interactive monitoring system for coral bleaching using high-resolution SST and improves stress predictors. Geo. Res. Lett. 2008, 35, L05603. [Google Scholar] [CrossRef]
- Liu, G.; Skirving, W.J.; Strong, A.E. Remote sensing of sea surface temperatures during 2002 barrier reef coral bleaching. Eos, Trans. Am. Geophys. Union 2003, 84, 137–144. [Google Scholar] [CrossRef]
- Liu, G.; Strong, A.E.; Skirving, W.J.; Arzayus, L.F. Overview of NOAA Coral Reef Watch Program’s Near-real-time Satellite Global Bleaching Monitoring Activities. In Proceedings of the 10th international coral reef symposium, Okinawa, Japan, 28 June–2 July 2004; NOAA: Washington, DC, USA, 2006; pp. 1783–1793. [Google Scholar]
- Liu, G.; Rauenzahn, J.L.; Heron, S.F.; Eakin, C.M.; Skirving, W.J.; Christensen, T.R.L.; Strong, A.E. NOAA coral reef watch 50 km satellite sea surface temperature-based decision support system for coral bleaching management. In NOAA technical Report NESDIS 143; NOAA: Washington, DC, USA, 2013; p. 33. [Google Scholar]
- Kayanne, H. Validation of degree heating weeks as a coral bleaching index in the northwestern Pacific. Coral Reefs 2017, 36, 63–70. [Google Scholar] [CrossRef]
- Smith, T.B.; Nemeth, R.S.; Blondeau, J.; Clanan, J.M.; Kadison, E.; Herzlied, S. Assessing coral reef heath across onshore to offshore stress gradients in the US Virgin Islands. Mar. Poll. Bull. 2008, 56, 1983–1991. [Google Scholar] [CrossRef]
- Mies, M.; Chaves-Filho, A.B.; Miyamoto, S.; Guth, A.Z.; Tenório, A.A.; Castro, C.B.; Pires, D.O.; Calderon, E.N.; Sumida, P.Y.G. Production of three symbiosis-related fatty acids by Symbiodinium types in clades A-F associated with marine invertebrate larvae. Coral Reefs 2017, 36, 1319–1328. [Google Scholar] [CrossRef]
- Conlan, J.A.; Rocker, M.M.; Francis, D.S. A comparison of two common sample preparation techniques for lipid and fatty acid analysis in three different coral morphotypes reveals quantitative and qualitative differences. Peer J. 2017, 5, e3645. [Google Scholar] [CrossRef]
- Yuama, I.; Harii, S.; Hidaka, M. Algal symbiont type affects gene expression in juveniles of the coral Acropora tenuis exposed thermal stress. Mar. Envir. Res. 2012, 76, 41–47. [Google Scholar] [CrossRef]
- Fuller, W.A. Introduction to statistical time series; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Roder, C.; Fillinger, L.; Jantzen, C.; Schmidt, G.M.; Khokiattiwong, S.; Richter, C. Trophic response of corals to large amplitude internal waves. Mar. Ecol. Prog. Ser. 2010, 412, 113–128. [Google Scholar] [CrossRef]
- Chelton, D.B. Commentary: Short-term climatic variability in the Northeast Pacific Ocean. In The influence of ocean conditions on the production of salmonids in the North Pacific; Pearcy, W., Ed.; Oregon State University Press: Corvallis, OR, USA, 1984; pp. 87–99. [Google Scholar]
- Allen, J.I.; Moore, M.N. Environmental prognostics: is the current use of biomarkers appropriate for environmental risk evaluation. Mar. Environ. Res. 2004, 58, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’hara, R.B.; Wagner, H. R Package ‘vegan’. Community ecology package, version, 2(9). 2013. [Google Scholar]
- Kassambara, A.; Mundt, F. Factoextra. Extract and Visualize the Results of Multivariate Data Analyses. R package version 2016. [Google Scholar]
- Le Hohaic, M.; Ross, C.L.; Cornwall, C.E.; Comeau SLowe, R.; McCulloch, M.T.; Schoepf, V. Marine heatwave causes unprecedented regional mass bleaching of thermally resistant corals in northwestern Australia. Sci. Rep. 2017, 7, 14999. [Google Scholar] [CrossRef] [PubMed]
- Costa, O.S.; Attrill, M.J.; Nimmo, M. Seasonal and spatial controls on the delivery of excess nutrients to nearshore and offshore coral reefs of Brazil. J. Mar. Sys. 2006, 60, 63–74. [Google Scholar] [CrossRef]
- Glynn, P.W.; Weerdt, W.H. Elimination of two reef-building hydrocorals following the 1982-83 El Niño warming event. Science 1991, 253, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Trapido-Rosenthal, H.G.; Sharp, K.H.; Galloway, T.S.; Morrall, C.E. Nitric oxide and cnidarian–dinoflagellate symbioses: pieces of a puzzle. Amer. Zool. 2001, 41, 247–257. [Google Scholar] [CrossRef][Green Version]
- Weis, V.M. Cellular mechanisms of Cnidarian bleaching: stress causes the collapse of symbiosis. J. Exp. Biol. 2008, 211, 59–66. [Google Scholar] [CrossRef]
- Anthony, K.R.N.; Fabricius, K.E. Shifting roles of heterotrophy and autotrophy in coral energetics under varying turbidity. J. Exp. Mar. Biol. Ecol. 2000, 252, 221–253. [Google Scholar] [CrossRef]
- Nunes, F.; Fukami, H.; Vollmer, S.V.; Norris, R.D.; Knowlton, N. Re-evaluation of the systematics of the endemic corals of Brazil by molecular data. Coral Reefs 2008, 27, 423–432. [Google Scholar] [CrossRef]
- Siebeck, U.E.; Marshall, N.J.; Kluter, A.; Hoegh-Guldberg, O. Monitoring coral bleaching using a colour reference card. Coral Reefs 2006, 25, 453–460. [Google Scholar] [CrossRef]
- Amaral, F.M.D.; Steiner, A.Q.; Broadhurst, M.K.; Cairns, S.D. An overview of the shallow-water calcified hydroid from Brazil (Hydrozoa: Cnidaria), including the description of a new species. Zootaxa 2008, 1930, 56–68. [Google Scholar] [CrossRef]
- Hoogenboom, M.; Rodolfo-Metalpa, R.; Ferrier-Pagès, C. Co-variation between autotrophy and heterotrophy in the Mediterranean coral Cladocora caespitosa. J. Exp. Biol. 2010, 213, 2399–2409. [Google Scholar] [CrossRef] [PubMed]
- Ferrier-Page‘s, C.; Peirano, A.; Abbate, M.; Cocito, S.; Negri, A.; Rottier, C.; Riera, P.; Rodolfo-Metalpa, R.; Reynaud, S. Summer autotrophy and winter heterotrophy in the temperate symbiotic coral Cladocora caespitosa. Limnol. Oceanogr. 2011, 56, 1429–1438. [Google Scholar] [CrossRef]
- Perez SWeis, V. Nitric oxide and cnidarians bleaching: an eviction notice mediates breakdown of a symbiosis. J. Exp. Biol. 2006, 209, 2804–2810. [Google Scholar] [CrossRef] [PubMed]
- Ross, C. Nitric oxide and heat shock protein 90 co-regulate temperature-induced bleaching in the soft coral Eunicea fusca. Coral Reefs 2014, 33, 513–522. [Google Scholar] [CrossRef]
- Jin, Y.K.; Lundgren, P.; Lutz, A.; Raina, J.-B.; Howells, E.J.; Paley, A.S.; Willis, B.L.; van Oppen, M.J.H. Genetic markers for antioxidant capacity in a ree-building coral. Sci. Adv. 2016, 2, e1500842. [Google Scholar] [CrossRef]
- Marangoni, L.F.B.; Dalmolin, C.; Marques, J.A.; Klein, R.D.; Abrantes, D.; Pereira, C.M.; Calderon, E.N.; Castro, C.B.; Bianchini, A. Oxidative stress biomarkers as potential tools in reef degradation monitoring: a study case in a South Atlantic reef under influence of the 2015-2016 El Niño/Southern Oscillation (ENSO). Ecol. Ind. 2019, 106, 105533. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reef Site | Taquaruçu | Funil | Mourão |
|---|---|---|---|
| Minimum temperature | 20.5 | 22.9 | 22.8 |
| Maximum temperature | 27.5 | 31.1 | 34.2 |
| Median | 24.7 | 26.5 | 26.7 |
| Mean | 24.7 | 26.5 | 26.7 |
| Standard deviation | 1.0 | 1.0 | 0.9 |
| Standard error | 0.2 | 0.2 | 0.2 |
| Site | Sea surface temperature anomaly x trophic mode | |||||||
| Millepora alcicornis | Mussimillia harttii | |||||||
| Lag (days) | R | Padj | dfadj | Lag (days) | R | Padj | dfadj | |
| Taquaruçu | - | −0.218 | 0.53 | 10.2 | 15 | −0.692 | 0.03 | 9.5 |
| Funil | - | −0.401 | 0.2 | 11.1 | 15 | −0.638 | 0.05 | 10.8 |
| Mourão | - | −0.577 | 0.053 | 11.4 | 30 | −0.783 | 0.004 | 9.4 |
| Peroxynitrite x bleaching | ||||||||
| Taquaruçu | - | −0.27 | 0.4 | 8.6 | 15 | 0.75 | 0.01 | 6.8 |
| Funil | 0 / 15 | 0.63 / 0.81 | 0.003/0.04 | 7.3 | 15/30 | 0.66 / 0.62 | 0.03 / 0.06 * | 7.6 |
| Mourão | - | 0.36 | 0.39 | 8.2 | - | 0.09 | 0.79 | 8.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marangoni, L.F.d.B.; Mies, M.; Güth, A.Z.; Banha, T.N.S.; Inague, A.; Fonseca, J.d.S.; Dalmolin, C.; Faria, S.C.; Ferrier-Pagès, C.; Bianchini, A. Peroxynitrite Generation and Increased Heterotrophic Capacity Are Linked to the Disruption of the Coral–Dinoflagellate Symbiosis in a Scleractinian and Hydrocoral Species. Microorganisms 2019, 7, 426. https://doi.org/10.3390/microorganisms7100426
Marangoni LFdB, Mies M, Güth AZ, Banha TNS, Inague A, Fonseca JdS, Dalmolin C, Faria SC, Ferrier-Pagès C, Bianchini A. Peroxynitrite Generation and Increased Heterotrophic Capacity Are Linked to the Disruption of the Coral–Dinoflagellate Symbiosis in a Scleractinian and Hydrocoral Species. Microorganisms. 2019; 7(10):426. https://doi.org/10.3390/microorganisms7100426
Chicago/Turabian StyleMarangoni, Laura Fernandes de Barros, Miguel Mies, Arthur Z. Güth, Thomás N. S. Banha, Alex Inague, Juliana da Silva Fonseca, Camila Dalmolin, Samuel Coelho Faria, Christine Ferrier-Pagès, and Adalto Bianchini. 2019. "Peroxynitrite Generation and Increased Heterotrophic Capacity Are Linked to the Disruption of the Coral–Dinoflagellate Symbiosis in a Scleractinian and Hydrocoral Species" Microorganisms 7, no. 10: 426. https://doi.org/10.3390/microorganisms7100426
APA StyleMarangoni, L. F. d. B., Mies, M., Güth, A. Z., Banha, T. N. S., Inague, A., Fonseca, J. d. S., Dalmolin, C., Faria, S. C., Ferrier-Pagès, C., & Bianchini, A. (2019). Peroxynitrite Generation and Increased Heterotrophic Capacity Are Linked to the Disruption of the Coral–Dinoflagellate Symbiosis in a Scleractinian and Hydrocoral Species. Microorganisms, 7(10), 426. https://doi.org/10.3390/microorganisms7100426