

Isolation and Characterization of a Crude Oil-Tolerant Obligate Halophilic Bacterium from the Great Salt Lake of the United States of America

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation of the Bacterial Strain
2.2. Preliminary Identification, Microscopy, and Biochemical Tests
2.3. Genome Analysis
2.4. Testing Crude Oil Tolerance of the Strain GSL5
2.5. Testing Crude Oil Metabolism by the Strain GSL5
2.6. Biofilm Assay
2.7. Susceptibility to Bacteriophage Infection
3. Results
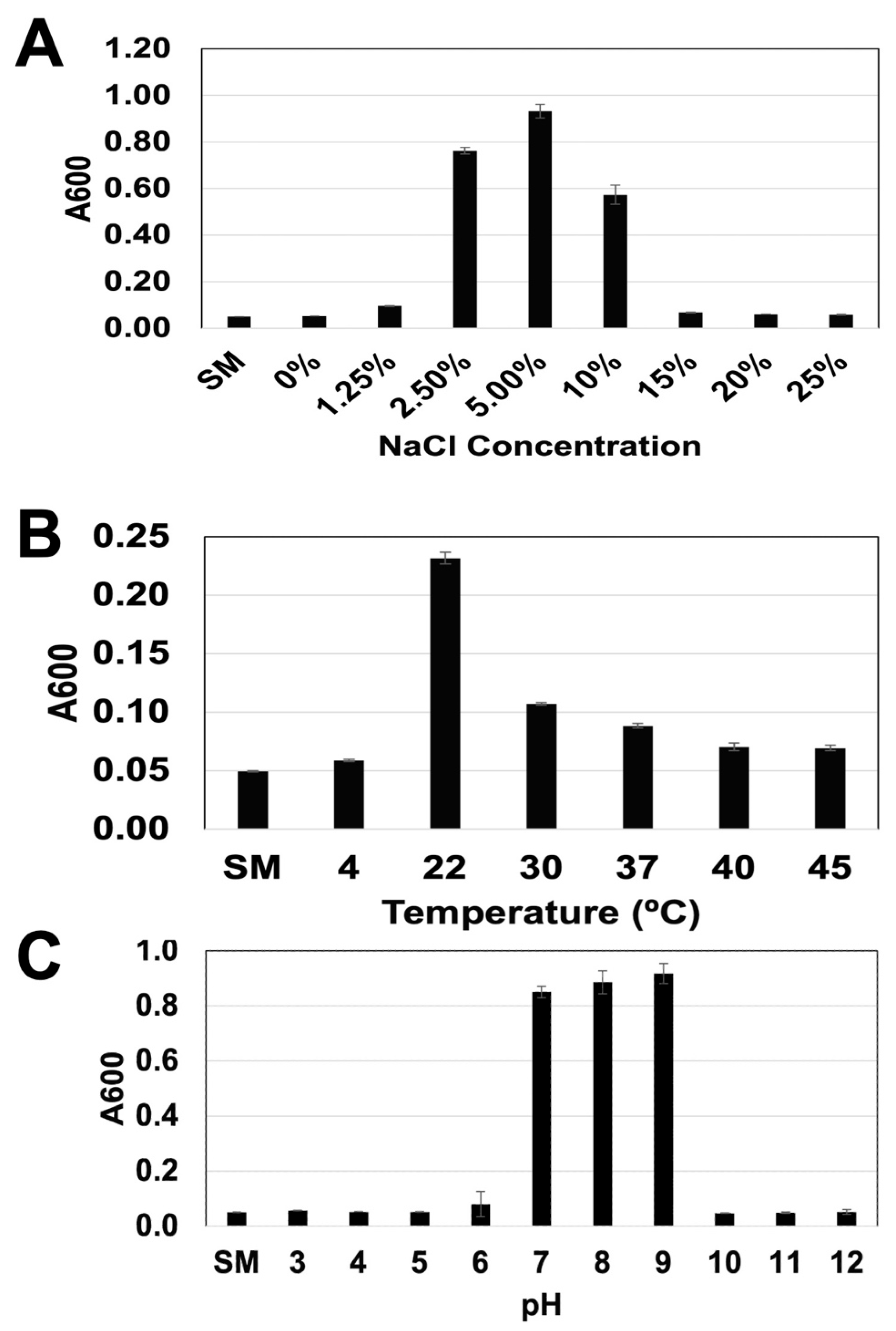
3.1. Microscopic, Biochemical, and Biophysical Characteristics
3.2. Phage Infection
3.3. Ability to Form a Biofilm
3.4. Crude Oil Tolerance and Metabolism
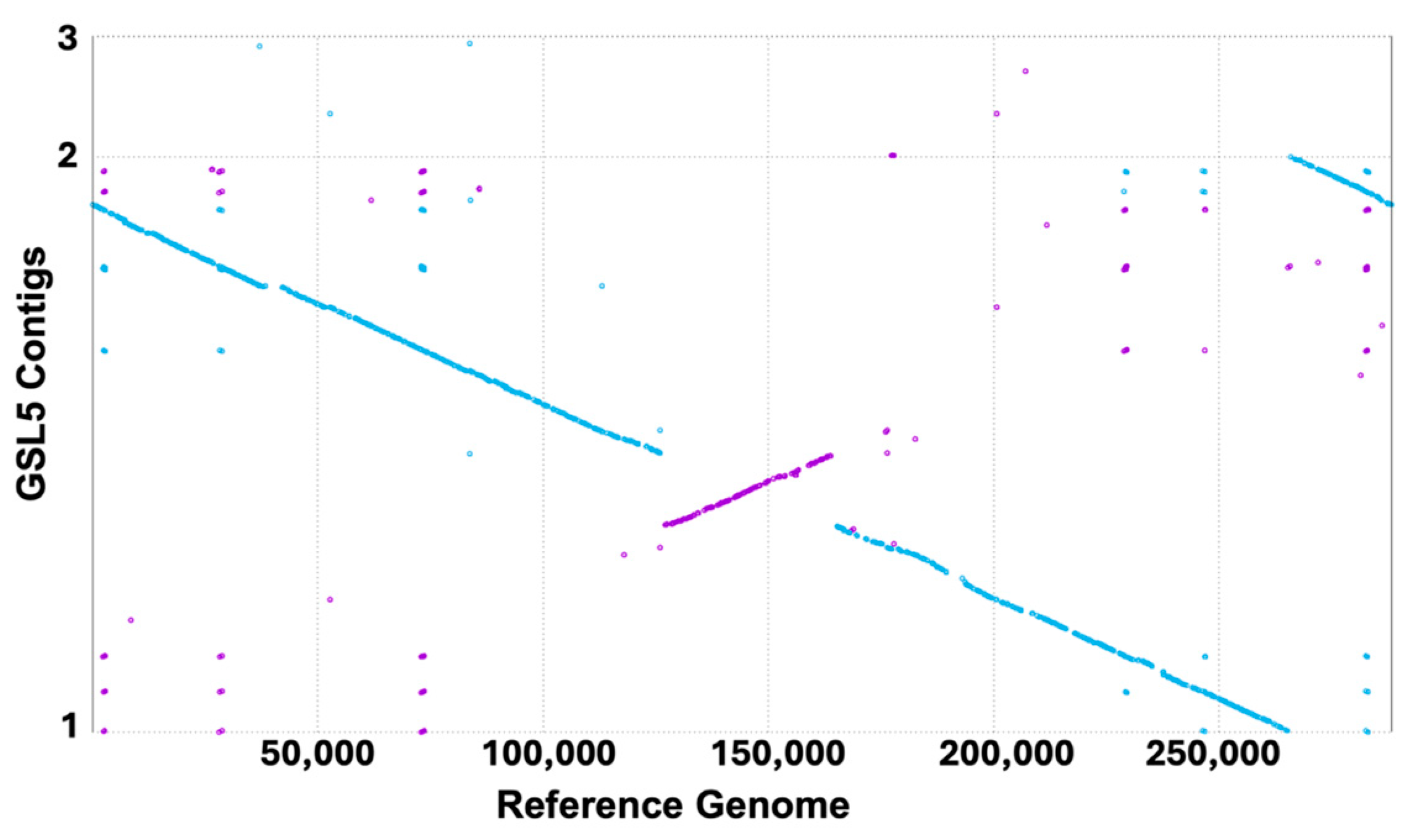
3.5. Genomic Analyses
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATCC | American Type Culture Collection, Manassas, Virginia |
| bbl | barrel |
| EPA | Environmental Protection Agency |
| GM | genetically modified |
| GSL5 | Great Salt Lake Strain 5 of Salinivibrio costicola |
| LB10 | Luria–Bertini broth containing 10% NaCl |
| LBA10 | Luria–Bertini broth containing 10% NaCl and 1.5% agar |
| NCBI | National Center for Biotechnology Information |
| rDNA | ribosomal DNA |
| TM | testing medium |
| TRPH | total recoverable petroleum hydrocarbons |
| ZOI | zone of inhibition |
References
- Goodman, R. Oil spill research: Salt and Freshwater. In Proceedings of the 29th Arctic Marine Oil Spill Program, Vancouver, BC, Canada, 6–8 June 2006; pp. 195–198. [Google Scholar]
- McNutt, M.K.; Camilli, R.; Crone, T.J.; Shaffer, F. Review of flow rate estimates of the Deepwater Horizon oil spill. Proc. Nat. Acad. Sci. USA 2011, 109, 20260–20267. [Google Scholar] [CrossRef] [PubMed]
- Oil Spills: Impact on the Ocean. Available online: http://www.waterencyclopedia.com/Oc-Po/Oil-Spills-Impact-on-the-Ocean.html (accessed on 24 July 2022).
- Manual on Oil Pollution; International Maritime Organization: London, UK, 1998.
- Saadoun, I.M.K. Impact of Oil Spills on Marine Life. In Emerging Pollutants in the Environments: Current and Future Implications; Larramendy, M.L., Soloneski, S., Eds.; InTech: Rijeka, Croatia, 2015. [Google Scholar] [CrossRef]
- Zhang, B.; Matchinksi, E.J.; Chen, B.; Ye, X.; Lee, K. Marine Spills—Oil Pollution, Sources and Effects. In World Seas: An Environmental Evaluation: Volume 3: Ecological Issues and Environmental Impacts, 2nd ed.; Sheppard, C., Ed.; Academic Press: New York, NY, USA, 2019. [Google Scholar]
- Agrawal, M. Ten Methods of Oil Spill Cleanup at Sea, Marine Insight. 2021. Available online: https://www.marineinsight.com/environment/10-methods-for-oil-spill-cleanup-at-sea/ (accessed on 24 July 2024).
- Dave, D.; Ghaly, A.E. Remediation Technologies for Marine Oil Spills: A Critical Review and Comparative Analysis. Am. J. Environ. Sci. 2011, 7, 423440. [Google Scholar] [CrossRef]
- Kalia, A.; Sharma, S.; Semor, N.; Babele, P.K.; Sagar, S.; Bhatia, R.K.; Walia, A. Recent advancements in hydrocarbon bioremediation and future challenges: A review. 3 Biotechnol. 2022, 12, 135. [Google Scholar] [CrossRef]
- Atlas, R.; Bragg, J. Bioremediation of marine oil spills: When and when not—The Exxon Valdez experience. Microb. Biotechnol. 2009, 2, 213–221. [Google Scholar] [CrossRef]
- Joshi, S.J. Microbial Biotechnology and Environmental Bioremediation: Challenges and Prospects. Open Biotechnol. J. 2016, 10, TOBIOTJ-10-287. [Google Scholar] [CrossRef]
- Ranveer, S.A.; Dasriya, V.; Ahmad, M.F.; Dhillon, H.S.; Samtiya, M.; Shama, E.; Anand, T.; Dhewa, T.; Chaudhary, V.; Chaudhary, P. Positive and negative aspects of bacteriophages and their immense role in the food chain. npj Sci. Food 2024, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Kebede, G.; Tafese, T.; Abda, E.M.; Kamaraj, M.; Assefa, F. Factors Influencing the Bacterial Bioremediation of Hydrocarbon Contaminants in the Soil: Mechanisms and Impacts. J. Chem. 2021, 2021, 9823362. [Google Scholar] [CrossRef]
- Hassanshahian, M.; Tebyanian, H.; Cappello, S. Isolation and characterization of two crude oil-degrading yeast strains, Yarrowia lipolytica PG-20 and PG-32, from the Persian Gulf. Mar. Pollut. Bull. 2012, 64, 1386–1391. [Google Scholar] [CrossRef]
- Arachchige, M.S.A.; Yoshida, S.; Toyama, H. Thermo- and salt-tolerant Saccharomyces cerevisiae strains isolated from fermenting coconut toddy from Sri Lanka. Biotechnol. Biotechnol. Equip. 2019, 33, 937–944. [Google Scholar] [CrossRef]
- Ojewumi, M.E.; Okeniyi, J.O.; Ikotun, J.O.; Okeniyi, E.T.; Ejemen, V.A.; Popoola, A.P.I. Bioremediation: Data on Pseudomonas aeruginosa effects on the bioremediation of crude oil-polluted soil. Data Brief. 2018, 19, 101–113. [Google Scholar] [CrossRef]
- Raghavan, P.U.M.; Vivekanandan, M. Bioremediation of oil-spilled sites through seeding of naturally adapted Pseudomonas putida. Int. Biodeterior. Biodegrad. 1999, 44, 29–32. [Google Scholar] [CrossRef]
- Rahman, K.S.M.; Thahira-Rahman, J.; Lakshmanaperumalsamy, P.; Banat, I.M. Towards Efficient Crude Oil Degradation by a Mixed Bacterial Consortium. Bioresour. Technol. 2002, 85, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Yakimov, M.M.; Timmis, K.N.; Golyshin, P.N. Obligate oil-degrading marine bacteria. Curr. Opin. Biotechnol. 2007, 18, 257–266. [Google Scholar] [CrossRef]
- Dell’Anno, F.; Rastelli, E.; Sansone, C.; Brunet, C.; Ianora, A.; Dell’Anno, A. Bacteria, Fungi and Microalgae for the Bioremediation of Marine Sediments Contaminated by Petroleum Hydrocarbons in the Omics Era. Microorganisms 2021, 9, 1695. [Google Scholar] [CrossRef] [PubMed]
- Hassanshahian, M.; Emtiazi, G.; Cappello, S. Isolation and characterization of crude-oil-degrading bacteria from the Persian Gulf and the Caspian Sea. Mar. Pollut. Bull. 2012, 64, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, J.; Yergeau, E.; Fortin, N.; Cobanli, S.; Elias, M.; King, T.L.; Lee, K.; Greer, C.W. Chemical Dispersants Enhance the Activity of Oil- and Gas Condensate-Degrading Marine Bacteria. ISME J. 2017, 11, 2793–2808. [Google Scholar] [CrossRef]
- Martínez-Espinosa, R.M. Halophilic archaea as tools for bioremediation technologies. Appl. Microbiol. Biotechnol. 2024, 108, 401. [Google Scholar] [CrossRef]
- Krzmarzick, M.J.; Taylor, D.K.; Fu, X.; McCutchan, A.L. Diversity and Niche of Archaea in Bioremediation. Archaea 2018, 2018, 3194108. [Google Scholar] [CrossRef]
- Al-Mailem, D.M.; Eliyas, M.; Radwan, S.S. Enhanced haloarchaeal oil removal in hypersaline environments via organic nitrogen fertilization and illumination. Extremophiles 2012, 16, 751–758. [Google Scholar] [CrossRef]
- Maricy, R.L.; Bonfá, M.R.L.; Grossman, M.J.; Mellado, E.; Durrant, L.R. Biodegradation of aromatic hydrocarbons by Haloarchaea and their use for the reduction of the chemical oxygen demand of hypersaline petroleum produced water. Chemosphere 2011, 84, 1671–1676. [Google Scholar] [CrossRef]
- Huwang, J.; Ge, X.; Sun, M. Modified CTAB Protocol Using a Silica Matrix for Isolation of Plant. Genom. DNA Biotechnol. 2000, 28, 432–434. [Google Scholar] [CrossRef]
- Berschick, P. One Primer Pair Amplifies Small Subunit Ribosomal DNA From Mitochondria, Plastids and Bacteria. Biotechniques 1997, 23, 494–498. [Google Scholar] [CrossRef] [PubMed]
- Hudzicki, J.; Kirby-Bauer Disc Diffusion Test Protocol. American Society of Microbiology. Available online: https://asm.org/getattachment/2594ce26-bd44-47f6-8287-0657aa9185ad/Kirby-Bauer-Disk-Diffusion-Susceptibility-Test-Protocol-pdf.pdf (accessed on 24 December 2024).
- Kolmogorov, M.; Yuan, J.; Lin, Y.; Pevzner, P.A. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- The Galaxy Community. The Galaxy platform for accessible, reproducible, and collaborative data analyses: 2024 update. Nucleic Acids Res. 2024, 52, W83–W94. [Google Scholar] [CrossRef]
- Kurtz, S.; Phillippy, A.; Delcher, A.L.; Smoot, M.; Shumway, M.; Antonescu, C.; Salzberg, S.L. Versatile and open software for comparing large genomes. Genome Biol. 2004, 5, R12. [Google Scholar] [CrossRef]
- Liaw, H.J.; Mah, R.A. Isolation and characterization of Haloanaerobacter chitinovorans gen. nov., sp. nov., a halophilic, anaerobic, chitinolytic bacterium from a solar saltern. Appl. Environ. Microbiol. 1992, 58, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.K.; Rao, T.S. An Improved Crystal Violet Assay for Biofilm Quantification in 96-Well Microtitre Plate, BioRxiv-ColdSpring Harbor Laboratory. Available online: https://www.biorxiv.org/content/10.1101/100214v1 (accessed on 25 July 2022).
- Kamiński, B.; Paczesny, J. Bacteriophage Challenges in Industrial Processes: A Historical Unveiling and Future Outlook. Pathogens 2024, 13, 152. [Google Scholar] [CrossRef]
- Huang, C.; Virk, S.M.; Shi, J.; Zhou, Y. Isolation, characterization, and application of bacteriophage LPSE1 against Salmonella enterica in ready-to-eat (RTE) foods. Front. Microbiol. 2018, 9, 1046. [Google Scholar] [CrossRef]
- Miller, I.P.; Laney, A.G.; Zahn, G.; Sheehan, B.J.; Whitley, K.V.; Kuddus, R.H. Isolation and preliminary characterization of a novel bacteriophage vB_KquU_φKuK6 that infects the multidrug-resistant pathogen Klebsiella quasipneumoniae. Front. Microbiol. 2024, 15, 1472729. [Google Scholar] [CrossRef]
- Community Phages. Phage Enrichment. Available online: https://phages.hms.harvard.edu/labprotocols/protocol_06-enrichments.html (accessed on 29 May 2025).
- Pal, S.; Kundu, A.; Banerjee, T.D.; Mohapatra, B.; Roy, A.; Manna, R.; Sar, P.; Kazy, S.K. Genome analysis of crude oil degrading Franconibacter pulveris strain DJ34 revealed its genetic basis for hydrocarbon degradation and survival in oil contaminated environment. Genomics 2017, 109, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Liu, W.; Tian, S.; Wang, W.; Qi, Q.; Jiang, P.; Gao, X.; Li, F.; Li, H.; Yu, H. Petroleum Hydrocarbon-Degrading Bacteria for the Remediation of Oil Pollution Under Aerobic Conditions: A Perspective Analysis. Front. Microbiol. 2018, 9, 2885. [Google Scholar] [CrossRef]
- Yuniati, M.D. Bioremediation of petroleum-contaminated soil: A Review. IOP Conf. Ser. Earth Environ. Sci. 2018, 118, 012063. [Google Scholar] [CrossRef]
- Hassanshahian, M. Isolation and characterization of biosurfactant producing bacteria from Persian Gulf (Bushehr provenance). Mar. Pollut. Bull. 2014, 86, 361–366. [Google Scholar] [CrossRef]
- Atlas, R.M. Microbial degradation of petroleum hydrocarbons: An environmental perspective. Microbiol. Rev. 1981, 45, 180–209. [Google Scholar] [CrossRef] [PubMed]
- Jemli, M.; Aloui, F.; Karray, F.; Sayadi, S. Organic pollutants biodegradation by halophile-isolated bacteria in saline conditions. Desalination Water Treat. 2018, 113, 227–234. [Google Scholar] [CrossRef]
- Utah Division of Wildlife Resources. About the Great Salt Lake. Available online: https://wildlife.utah.gov/gslep/about.html#:~:text=Among%20the%20lake%E2%80%99s%20most%20unique,salina)%20and%20violet%2Dpinkish%20halophiles (accessed on 24 December 2024).
- Utah State University Institute for Land, Water and Air. An Increasing Threat of Toxic Dust from a Drying Great Salt Lake. Available online: https://www.usu.edu/ilwa/reports/2022/great-salt-lake/5d-toxic-dust (accessed on 24 December 2024).
- Wayne, A.; Wurtsbaugh, W.A.; Leavitt, P.R.; Moser, K.A. Effects of a century of mining and industrial production on metal contamination of a model saline ecosystem, Great Salt Lake, Utah. Environ. Pollut. 2020, 266, 115072. [Google Scholar] [CrossRef]
- Huang, C.Y.; Garcia, J.L.; Patel, B.K.; Cayol, J.L.; Baresi, L.; Mah, R.A. Salinivibrio costicola subsp. vallismortis subsp. nov., a halotolerant facultative anaerobe from Death Valley, and emended description of Salinivibrio costicola. Int. J. Syst. Evol. Microbiol. 2000, 50, 615–622. [Google Scholar] [CrossRef]
- Garcia, M.T.; Ventosa1, A.; Ruiz-Berraquero, F.; Kocur, M. Taxonomic Study and Amended Description of Vibrio costicola. Int. J. Syst. Evol. Microbiol. 1987, 37, 251–256. [Google Scholar] [CrossRef]
- Seger, D.A. Introduction to Ocean Sciences, 5th ed. Available online: https://www.reefimages.com/oceansci.php (accessed on 24 December 2024).
- Chattopadhyay, I.; Banu, R.; Usman, T.M.M.; Varjani, S. Exploring the role of microbial biofilm for industrial effluents treatment. Bioengineered 2022, 13, 6420–6440. [Google Scholar] [CrossRef]
- Mohamad, F.; Alzahrani, R.R.; Alsaadi, A.; Alrfaei, B.M.; Yassin, A.E.B.; Alkhulaifi, M.M.; Halwani, M. An Explorative Review on Advanced Approaches to Overcome Bacterial Resistance by Curbing Bacterial Biofilm Formation. Infect. Drug Resist. 2023, 16, 19–49. [Google Scholar] [CrossRef] [PubMed]
- LPSN (List of Prokaryotic Names with Standing Nomenclature). Species Slainivibrio costicola. Available online: https://lpsn.dsmz.de/species/salinivibrio-costicola#:~:text=The%20full%20classification%20is:%20risk%20group%20=,animal%20pathogen%20under%20Canadian%20Food%20Inspection%20Agency (accessed on 24 December 2024).
- Rosenberg, E.; Bittan-Banin, G.; Sharon, G.; Shon, A.; Hershko, G.; Levy, I.; Ron, E.Z. The Phage-Driven Microbial Loop in Petroleum Bioremediation. Microb. Biotechnol. 2010, 3, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Liu, X.; Ai, J.; Wang, J.; Guo, Y.; Liu, X.; He, X.; Deng, Z.; Jiang, Y. Microbial community succession during crude oil-degrading bacterial enrichment cultivation and construction of a degrading consortium. Front. Microbiol. 2022, 13, 1044448. [Google Scholar] [CrossRef]
- Kurylo, C.M.; Alexander, N.; Dass, R.A.; Parks, M.M.; Altman, R.A.; Vincent, C.T.; Mason, C.E.; Blanchard, S.C. Genome Sequence and Analysis of Escherichia coli MRE600, a Colicinogenic, Nonmotile Strain that Lacks RNase I and the Type I Methyltransferase, EcoKI. Genome Biol. Evol. 2016, 8, 742–752. [Google Scholar] [CrossRef]
- Perdigão, R.; Almeida, C.M.R.; Magalhães, C.; Ramos, S.; Carolas, A.L.; Ferreira, B.S.; Carvalho, M.F.; Mucha, A.P. Bioremediation of Petroleum Hydrocarbons in Seawater: Prospects of Using Lyophilized Native Hydrocarbon-Degrading Bacteria. Microorganisms 2021, 9, 2285. [Google Scholar] [CrossRef] [PubMed]
- Alabresm, A.; Chen, Y.P.; Decho, A.W.; Lead, J. A novel method for the synergistic remediation of oil-water mixtures using nanoparticles and oil-degrading bacteria. Sci. Total Environ. 2018, 630, 1292–1297. [Google Scholar] [CrossRef]
- Li, J.; Gao, Y.; Dong, H.; Sheng, G.P. Haloarchaea, excellent candidates for removing pollutants from hypersaline wastewater. Trends Biotechnol. 2022, 40, 226–239. [Google Scholar] [CrossRef]
- Kumar, S.; Zhou, J.; Li, M.; Xiang, H.; Zhao, D. Insights into the metabolism pathway and functional genes of long-chain aliphatic alkane degradation in haloarchaea. Extremophiles 2020, 24, 475–483. [Google Scholar] [CrossRef]
- French, K.E.; Zhou, Z.; Terry, N. Horizontal ‘gene drives’ harness indigenous bacteria for bioremediation. Sci. Rep. 2020, 10, 15091. [Google Scholar] [CrossRef]
- Garbisu, C.; Garaiyurrebaso, O.; Epelde, L.; Grohmann, E.; Alkorta, I. Plasmid-Mediated Bioaugmentation for the Bioremediation of Contaminated Soils. Front. Microbiol. 2017, 8, 1966. [Google Scholar] [CrossRef]
- Samin, G.; Pavlova, M.; Arif, M.I.; Postema, C.P.; Damborsky, J.; Janssen, D.B.; Parales, R.E. A Pseudomonas putida Strain Genetically Engineered for 1,2,3-Trichloropropane Bioremediation. Appl. Environ. Microbiol. 2014, 80, 5467–5476. [Google Scholar] [CrossRef] [PubMed]
- Davidson, J. Towards safer vectors for the field release of recombinant bacteria. Environ. Biosaf. Res. 2002, 1, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Shah, G.; Singhal, K.; Soni, V. Comprehensive insights into the impact of oil pollution on the environment. Reg. Stud. Mar. Sci. 2024, 74, 103516. [Google Scholar] [CrossRef]
- Muhammad, R.; Boothman, C.; Song, H.; Lloyd, J.R.; van Dongen, E.B. Assessing the impacts of oil contamination on microbial communities in a Niger Delta soil. Sci. Total Environ. 2024, 926, 1783813. [Google Scholar] [CrossRef]
- Mataigne, V.; Vannier, N.; Vandenkoornhuyse, P.; Hacquard, S. Microbial Systems Ecology to Understand Cross-Feeding in Microbiomes. Front. Microbiol. 2021, 12, 780469. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Zhang, H. Microbial Consortia Are Needed to Degrade Soil Pollutants. Microorganisms 2022, 10, 261. [Google Scholar] [CrossRef]
- Rebello, S.; Nathan, V.K.; Sindhu, R.; Binod, P.; Awasthi, M.K.; Pandey, A. Bioengineered microbes for soil health restoration: Present status and future. Bioengineered 2021, 12, 12839–12853. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of the Medium | Description |
|---|---|
| TM-0 | LB broth (LB10) |
| TM-1 | Essential minerals [34] |
| TM-2 | Essential minerals and 1% glucose |
| TM-3 | Essential minerals and 1% crude oil |
| TM-4 | Essential minerals, 1% glucose, and 1% crude oil |
| TM-5 | LB broth and 1% crude oil |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oakes, J.; Kuddus, J.N.; Downs, E.; Oakey, C.; Davis, K.; Mohammad, L.; Whitely, K.; Hjelmen, C.E.; Kuddus, R. Isolation and Characterization of a Crude Oil-Tolerant Obligate Halophilic Bacterium from the Great Salt Lake of the United States of America. Microorganisms 2025, 13, 1568. https://doi.org/10.3390/microorganisms13071568
Oakes J, Kuddus JN, Downs E, Oakey C, Davis K, Mohammad L, Whitely K, Hjelmen CE, Kuddus R. Isolation and Characterization of a Crude Oil-Tolerant Obligate Halophilic Bacterium from the Great Salt Lake of the United States of America. Microorganisms. 2025; 13(7):1568. https://doi.org/10.3390/microorganisms13071568
Chicago/Turabian StyleOakes, Jonathan, Johurimam Noah Kuddus, Easton Downs, Clark Oakey, Kristina Davis, Laith Mohammad, Kiara Whitely, Carl E. Hjelmen, and Ruhul Kuddus. 2025. "Isolation and Characterization of a Crude Oil-Tolerant Obligate Halophilic Bacterium from the Great Salt Lake of the United States of America" Microorganisms 13, no. 7: 1568. https://doi.org/10.3390/microorganisms13071568
APA StyleOakes, J., Kuddus, J. N., Downs, E., Oakey, C., Davis, K., Mohammad, L., Whitely, K., Hjelmen, C. E., & Kuddus, R. (2025). Isolation and Characterization of a Crude Oil-Tolerant Obligate Halophilic Bacterium from the Great Salt Lake of the United States of America. Microorganisms, 13(7), 1568. https://doi.org/10.3390/microorganisms13071568