
Response of Endophytic Microbial Communities and Quality of Salvia miltiorrhiza to Fertilization Treatments

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experimental Design
2.3. Sample Collection
2.4. Determination of Medicinal Ingredient Content
2.5. Extraction of DNA and Illumina MiSeq Sequencing
2.6. Bioinformatic Analysis
2.7. Soil Physicochemical Properties
2.8. Isolation of Endophytic Fungi and Identification
2.9. Statistical Analysis
3. Results
3.1. Determination of Growth Indexes and Medicinal Ingredient Contents of S. miltiorrhiza
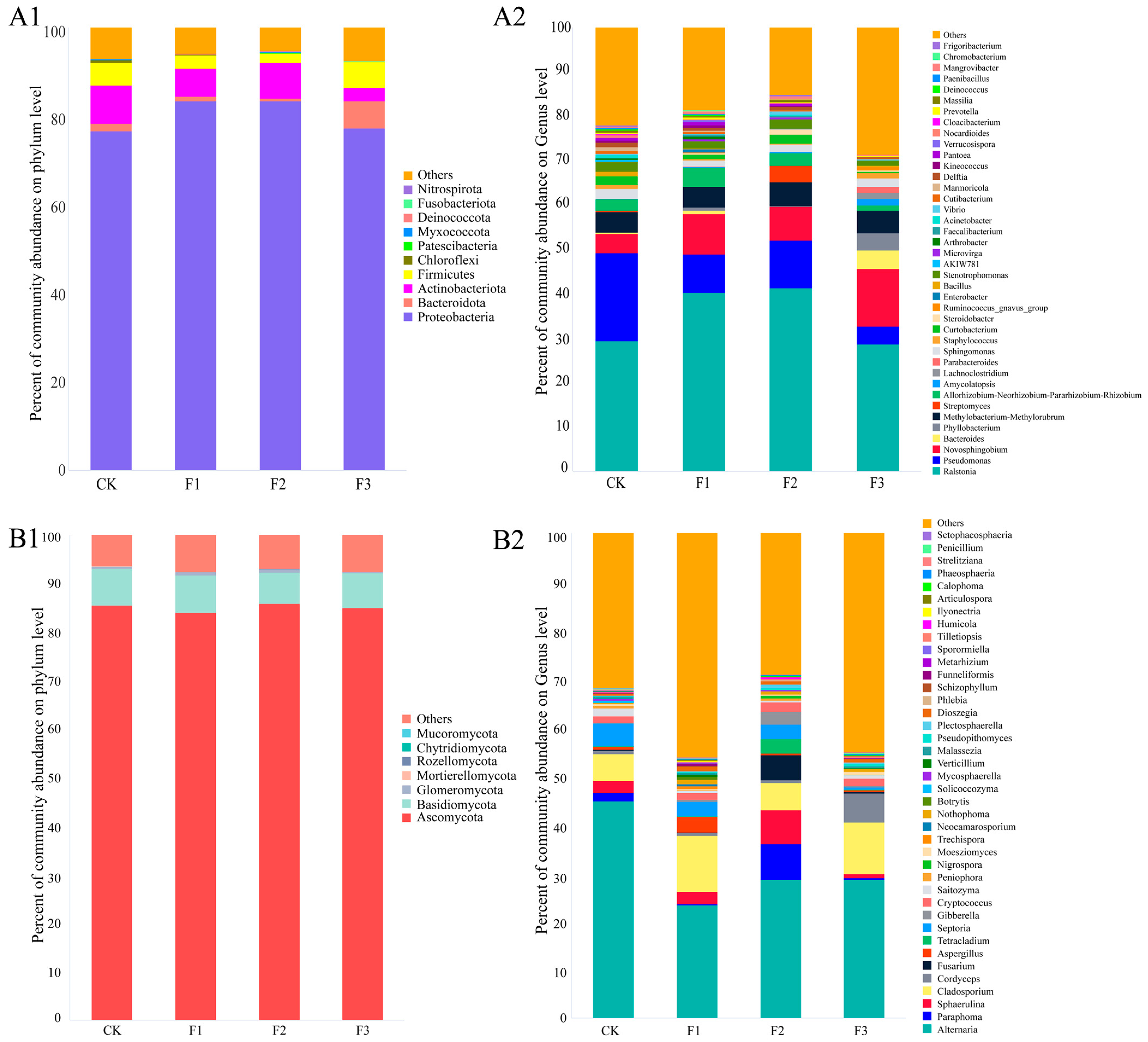
3.2. Changes in the Composition of the Endophyte Community of S. miltiorrhiza
3.3. Changes in the OTU Composition of the Endophyte Community of S. miltiorrhiza
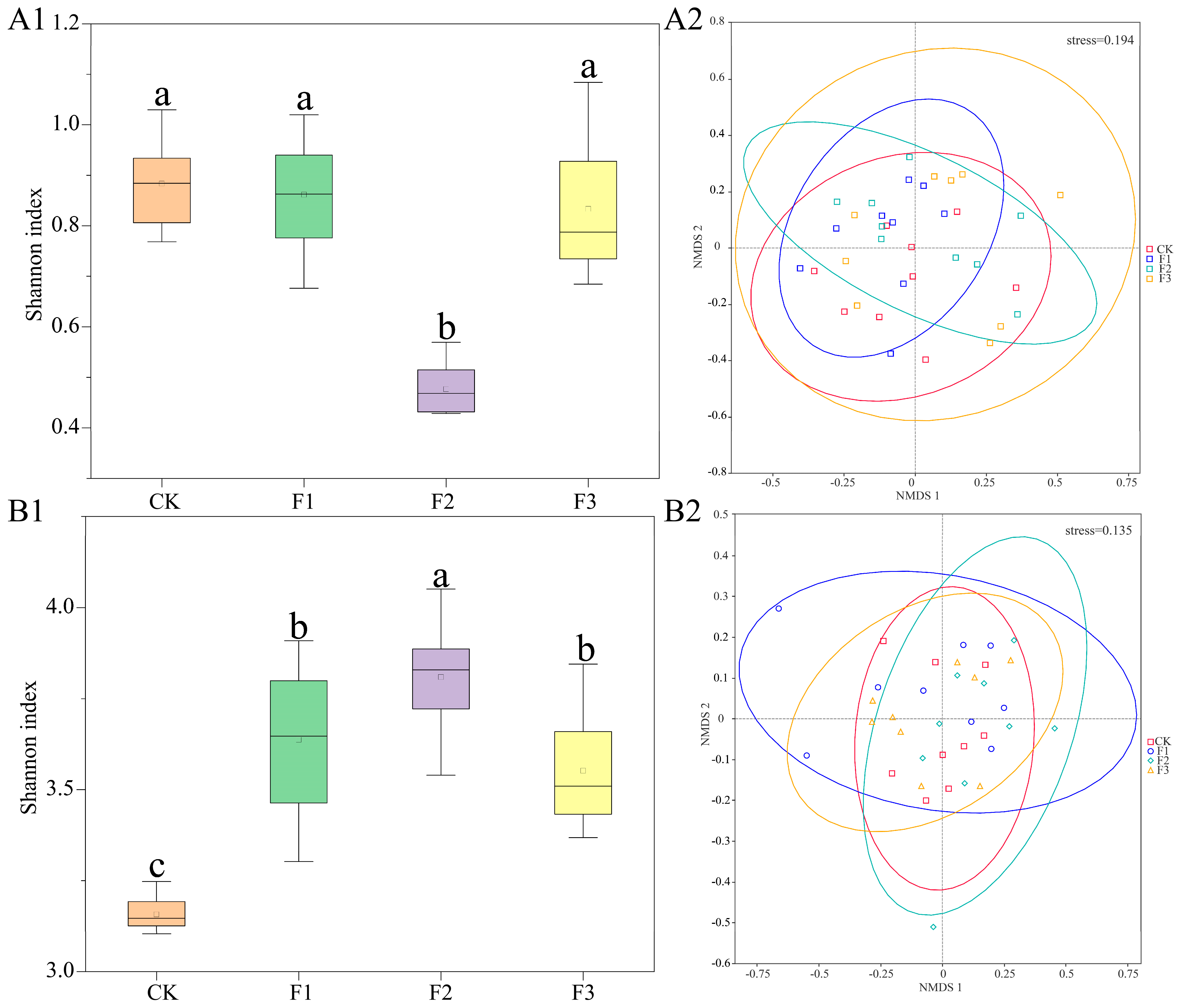
3.4. Changes in the Endophyte Community Diversity of S. miltiorrhiza
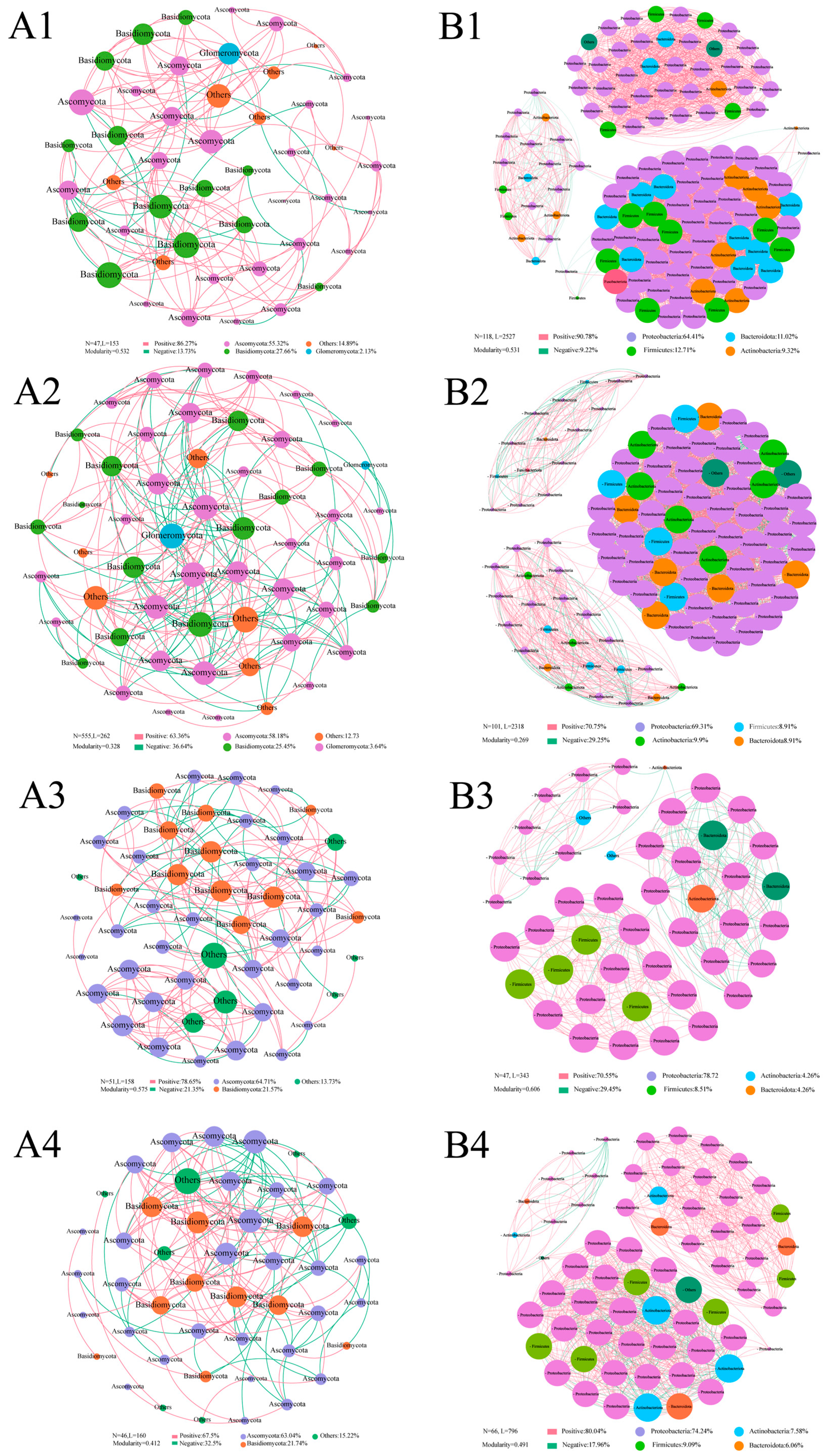
3.5. Co-Occurrence Network Analysis
3.6. Functional Prediction
3.7. Effect of Different Fertilizer Treatments on Soil Parameters
3.8. Correlation Analysis of Endophytic Fungal and Bacterial Communities with Growth Indicators, Medicine Ingredient Contents, and Soil Factors of S. miltiorrhiza
3.9. Variance Partitioning Analysis
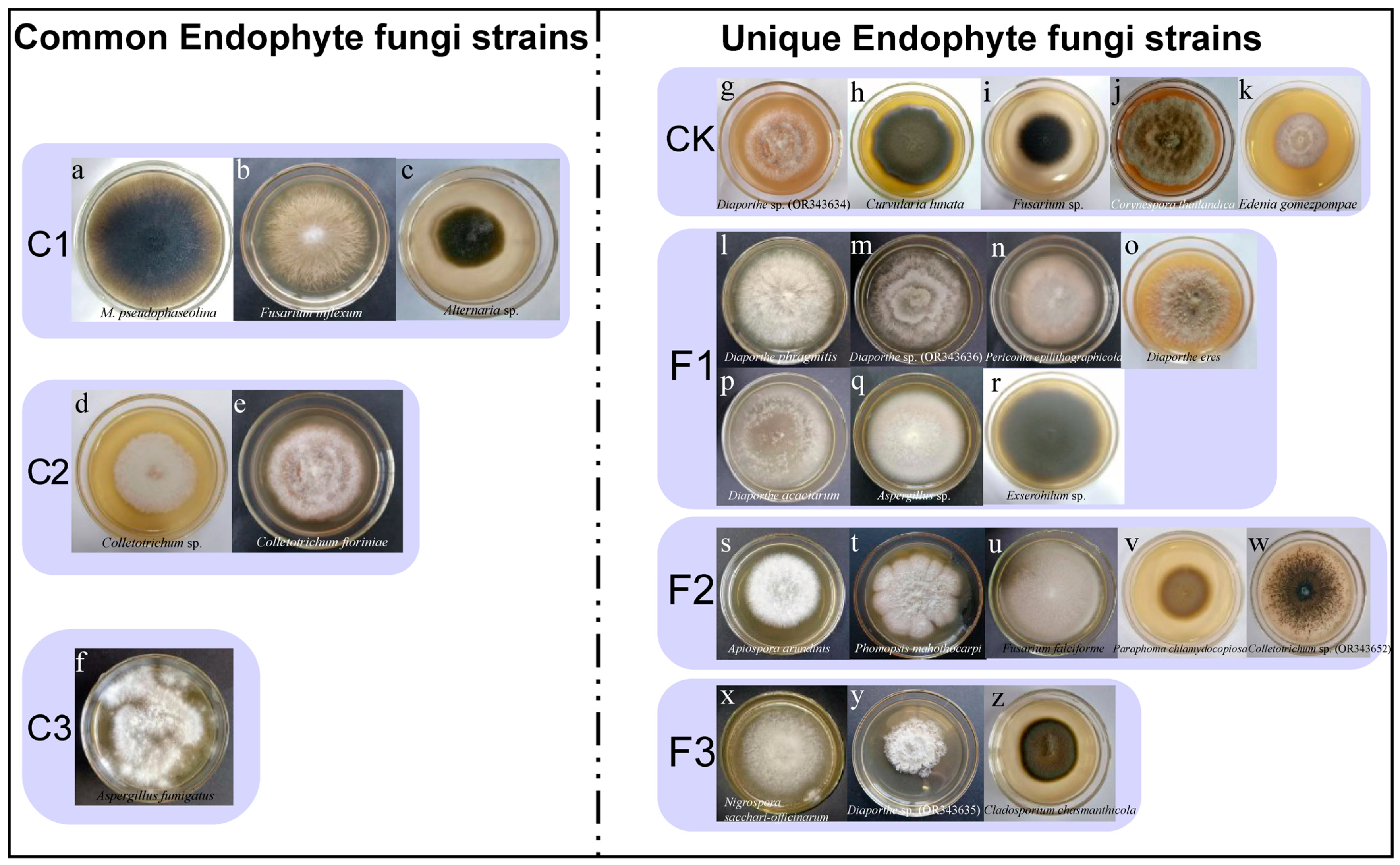
3.10. Diversity of Culturable Endophytic Fungi
4. Discussion
4.1. Effects of Different Fertilization Treatments on Endophytic Bacterial Communities of S. miltiorrhiza
4.2. Effects of Different Fertilization Treatments on Endophytic Fungal Communities of S. miltiorrhiza
4.3. Effect of Fertilization Treatments on Quality and Soil Factors of S. miltiorrhiza
4.4. Effects of Different Fertilization Treatments on Endophytic Microbial Communities of S. miltiorrhiza
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, Z.Q.; Gao, W.; Huang, L.Q. Tanshinones, critical pharmacological components in Salvia miltiorrhiza. Front. Pharmacol. 2019, 10, 202. [Google Scholar] [CrossRef] [PubMed]
- Dabbaghi, M.M.; Soleimani, R.H.; Safaei, R.; Baradaran, R.V.; Reza, F.M.; Reza, A.V. Unveiling the mechanism of protective effects of tanshinone as a new fighter against cardiovascular diseases: A systematic review. Cardiovasc. Toxicol. 2024, 24, 1467–1509. [Google Scholar] [CrossRef]
- Zhang, H.; Liu, Y.Y.; Jiang, Q.; Li, K.R.; Zhao, Y.X.; Cao, C.; Yao, J. Salvianolic acid A protects RPE cells against oxidative stress through activation of Nrf2/HO-1 signaling. Free Radic. Biol. Med. 2014, 69, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Xu, Q.Y.; Zhu, H.L.; Wang, J.Y.; Sun, N.; Bian, H.M.; Li, Y.; Lin, C. Salvianolic acid B promotes angiogenesis and inhibits cardiomyocyte apoptosis by regulating autophagy in myocardial ischemia. Chin. Med. 2023, 18, 155. [Google Scholar] [CrossRef]
- Li, S.J.; Zhang, X.; Wang, X.H.; Zhao, C.Q. Novel natural compounds from endophytic fungi with anticancer activity. Eur. J. Med. Chem. 2018, 156, 316–343. [Google Scholar] [CrossRef]
- Cao, P.; Wang, G.; Wei, X.M.; Chen, S.L.; Han, J.P. How to improve CHMs quality: Enlighten from CHMs ecological cultivation. Chin. Herb. Med. 2021, 13, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Buchwald, W. The effect of mineral fertilization on the content of phenolic acids and mass of roots of Salvia miltiorrhiza Bunge. Acta Agrobot. 2013, 57, 169–173. [Google Scholar] [CrossRef]
- Kwon, M.C.; Kim, Y.X.; Lee, S.; Jung, E.S.; Singh, D.; Sung, J.; Lee, C.H. Comparative metabolomics unravel the effect of magnesium oversupply on tomato fruit quality and associated plant metabolism. Metabolites 2019, 9, 231. [Google Scholar] [CrossRef]
- Januszkiewicz, R.; Kulczycki, G.; Samoraj, M. Foliar fertilization of crop plants in Polish agriculture. Agriculture 2023, 13, 1715. [Google Scholar] [CrossRef]
- Arsic, M.; Le Tougaard, S.; Persson, D.P.; Martens, H.J.; Doolette, C.L.; Lombi, E.; Schjoerring, J.K.; Husted, S. Bioimaging techniques reveal foliar phosphate uptake pathways and leaf phosphorus status. Plant Physiol. 2020, 183, 1472–1483. [Google Scholar] [CrossRef]
- Çolak Esetlili, B.; Pekcan, T.; Bayız, O.; Telli Karaman, H.; Ayata, M.; Sarı, G. Optimizing olive production: The role of foliar fertilizers in boosting yield and quality. Horticulturae 2024, 10, 1147. [Google Scholar] [CrossRef]
- Zhuang, Y.; Wang, X.Y.; Gong, X.; Bao, J.P. Effects of different foliar fertilizer treatments on fruit quality of the Korla fragrant pear. Horticulturae 2023, 10, 51. [Google Scholar] [CrossRef]
- Chen, H.; Song, Y.J.; Wang, S.S.; Fan, K.; Wang, H.; Mao, Y.L.; Zhang, J.; Xu, Y.; Yin, X.Y.; Wang, Y.; et al. Improved phyllosphere microbiome composition of tea plant with the application of small peptides in combination with rhamnolipid. BMC Microbiol. 2023, 23, 302. [Google Scholar] [CrossRef]
- Wang, D.S.; Deng, X.H.; Wang, B.; Zhang, N.; Zhu, C.Z.; Jiao, Z.X.; Li, R.; Shen, Q. Effects of foliar application of amino acid liquid fertilizers, with or without Bacillus amyloliquefaciens SQR9, on cowpea yield and leaf microbiota. PLoS ONE 2019, 14, 0222048. [Google Scholar] [CrossRef]
- Gong, F.; He, C.; Li, X.E.; Wang, K.H.; Li, M.; Zhou, X.Y.; Xu, M.H.; He, X.L. Impacts of fertilization methods on Salvia miltiorrhiza quality and characteristics of the epiphytic microbial community. Front. Plant Sci. 2024, 15, 1395628. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.J.; Wang, L.L.; Li, Q.; Shan, Q.M. Epiphytic and endophytic fungal communities of tomato plants. Hortic. Plant J. 2020, 7, 38–48. [Google Scholar] [CrossRef]
- Rani, S.; Kumar, P.; Dahiya, P.; Maheshwari, R.; Dang, A.S.; Suneja, P. Endophytism: A multidimensional approach to plant-prokaryotic microbe interaction. Front. Microbiol. 2022, 13, 861235. [Google Scholar] [CrossRef]
- Kumar, V.; Nautiyal, C.S. Plant abiotic and biotic stress alleviation: From an endophytic microbial perspective. Curr. Microbiol. 2022, 79, 311. [Google Scholar] [CrossRef]
- Yuan, Z.N.; Liu, Q.; Pang, Z.Q.; Fallah, N.; Liu, Y.M.; Hu, C.H.; Lin, W.X. Sugarcane rhizosphere bacteria community migration correlates with growth stages and soil nutrient. Int. J. Mol. Sci. 2021, 23, 10303. [Google Scholar] [CrossRef]
- Narayanan, Z.; Glick, B.R. Secondary metabolites produced by plant growth-promoting bacterial endophytes. Microorganisms 2022, 10, 2008. [Google Scholar] [CrossRef]
- Li, X.Y.; Lin, Y.L.; Qin, Y.; Han, G.Q.; Wang, H.; Yan, Z.Y. Beneficial endophytic fungi improve the yield and quality of Salvia miltiorrhiza by performing different ecological functions. PeerJ 2024, 12, 16959. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.; Luo, D.; Li, X.Q.; Han, T.; Jia, M.; Kong, Z.Y.; Ji, J.C.; Rahman, K.; Qin, L.P.; Zheng, C.J. Endophyte Chaetomium globosum D38 promotes bioactive constituents accumulation and root production in Salvia miltiorrhiza. Front. Microbiol. 2018, 8, 302133. [Google Scholar] [CrossRef]
- Lv, X.M.; Zhang, W.Y.; Chu, S.Y.; Zhang, H.H.; Wu, Y.Q.; Zhu, Y.; Yang, D.F.; Zhu, Y.H.; Mans, D.R.A.; Chen, H.M.; et al. Endophytic fungus Penicillium steckii DF33 promoted tanshinones biosynthesis in Salvia miltiorrhiza by regulating the expression of CYP450 genes. Gene 2024, 899, 148094. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Moon, C.D.; Zheng, N.; Huws, S.; Zhao, S.G.; Wang, J.Q. Opportunities and challenges of using metagenomic data to bring uncultured microbes into cultivation. Microbiome 2022, 10, 76. [Google Scholar] [CrossRef]
- Sadeghi, F.; Samsampour, D.; Seyahooei, M.A.; Bagheri, A.; Soltani, J. Diversity and spatiotemporal distribution of fungal endophytes associated with Citrus reticulata cv. Siyahoo. Curr. Microbiol. 2019, 76, 279–289. [Google Scholar] [CrossRef]
- Bodenhausen, N.; Horton, M.W.; Bergelson, J. Bacterial communities associated with the leaves and the roots of Arabidopsis thaliana. PLoS ONE 2013, 8, e56329. [Google Scholar] [CrossRef]
- Tepper, C.S.; Gaynor, S.C. Ribosomal internal transcribed spacer (ITS) DNA variation in millepora. J. Mar. Sci. Res. Dev. 2015, 6, 1000177. [Google Scholar] [CrossRef]
- Li, M.J.; Shao, D.T.; Zhou, J.C.; Gu, J.H.; Qin, J.J.; Chen, W.; Wei, W.Q. Signatures within esophageal microbiota with progression of esophageal squamous cell carcinoma. Chin. J. Cancer Res. 2020, 32, 755–767. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Holmes, S.P. Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 2017, 11, 2639–2643. [Google Scholar] [CrossRef]
- Nguyen, N.H.; Song, Z.W.; Bates, S.T.; Branco, S.; Tedersoo, L.; Menke, J.; Schilling, J.S.; Kennedy, P.G. FUNGuild: An open annotation tool for parsing fungal community datasets by ecological guild. Fungal Ecol. 2016, 20, 241–248. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G. PICRUSt2: An improved and customizable approach for metagenome inference. Nat. Biotechnol. 2019, 38, 685–688. [Google Scholar] [CrossRef] [PubMed]
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on ignition as a method for estimating organic and carbonate content in sediments: Reproducibility and comparability of results. J. Paleolimnol. 2001, 25, 101–110. [Google Scholar] [CrossRef]
- Xie, L.L.; He, X.L.; Wang, K.; Hou, L.F.; Sun, Q. Spatial dynamics of dark septate endophytes in the roots and rhizospheres of Hedysarum scoparium in northwest China and the influence of edaphic variables. Fungal Ecol. 2017, 26, 135–143. [Google Scholar] [CrossRef]
- Tarafdar, J.; Marschner, H. Phosphatase activity in the rhizosphere and hyphosphere of VA mycorrhizal wheat supplied with inorganic and organic phosphorus. Soil Biol. Biochem. 1994, 26, 387–395. [Google Scholar] [CrossRef]
- Hoffmann, G.G.; Teicher, K. A colorimetric technique for determining urease activity in soil. Dung Boden 1961, 95, 55–63. [Google Scholar] [CrossRef]
- He, K.; Li, X.P.; Dong, L.L. The effects of flue gas desulfurization gypsum (FGD gypsum) on P fractions in a coastal plain soil. J. Soils Sediments 2018, 18, 804–815. [Google Scholar] [CrossRef]
- Tian, H.Y.; Qiao, J.B.; Zhu, Y.J.; Jia, X.X.; Shao, M.A. Vertical distribution of soil available phosphorus and soil available potassium in the critical zone on the Loess Plateau, China. Sci. Rep. 2021, 11, 3159. [Google Scholar] [CrossRef]
- Cornelissen, J.H.; Lavorel, S.; Garnier, E.; Díaz, S.M.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; Steege, H.T.; Morgan, H.D.; Heijden, M.V.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Bremner, J.M. Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 1960, 55, 11–33. [Google Scholar] [CrossRef]
- Wu, Y.X.; Luo, Z.C.; Qi, L.; Zhang, R.; Wang, Y.X. A study of the relationship between initial grape yield and soil properties based on organic fertilization. Agronomy 2024, 14, 861. [Google Scholar] [CrossRef]
- Yang, C.B.; Feng, M.C.; Song, L.F.; Jing, B.H.; Xie, Y.K.; Wang, C.; Qin, M.X.; Yang, W.D.; Xiao, L.J.; Sun, J.J.; et al. Hyperspectral monitoring of soil urease activity under different water regulation. Plant Soil 2022, 477, 779–792. [Google Scholar] [CrossRef]
- Girish, P.S.; Haunshi, S.; Vaithiyanathan, S.; Rajitha, R.; Ramakrishna, C. A rapid method for authentication of Buffalo (Bubalus bubalis) meat by Alkaline Lysis method of DNA extraction and species specific polymerase chain reaction. J. Food Sci. Technol. 2013, 50, 141–146. [Google Scholar] [CrossRef]
- Rodriguez, R.; Redman, R.S. Fungal life-styles and ecosystem dynamics: Biological aspects of plant pathogens, plant endophytes and saprophytes. Adv. Bot. Res. 1996, 24, 169–193. [Google Scholar]
- Mataranyika, P.N.; Bez, C.; Venturi, V.; Chimwamurombe, P.M.; Uzabakiriho, J.D. Rhizospheric, seed, and root endophytic-associated bacteria of drought-tolerant legumes grown in arid soils of Namibia. Heliyon 2024, 10, e36718. [Google Scholar] [CrossRef]
- He, C.; Zhang, L.; Li, X.E. Plant performance and soil fungal community impacts of enhancing Dioscorea opposita with spraying foliar fertilizer with different nutrient element combinations. Agronomy 2022, 12, 2017. [Google Scholar] [CrossRef]
- Rivera-Zuluaga, K.; Hiles, R.; Barua, P.; Caldwell, D.; Iyer-Pascuzzi, A.S. Getting to the root of Ralstonia invasion. Semin. Cell Dev. Biol. 2023, 148–149, 3–12. [Google Scholar] [CrossRef]
- Hall, A.N.; Hall, B.W.; Kinney, K.J.; Olsen, G.G.; Banta, A.B.; Noguera, D.R.; Donohue, T.J.; Peters, J.M. Tools for genetic engineering and gene expression control in Novosphingobium aromaticivorans and Rhodobacter sphaeroides. Appl. Environ. Microbiol. 2024, 90, e0034824. [Google Scholar] [CrossRef] [PubMed]
- Vives-Peris, V.; Gómez-Cadenas, A.; Pérez-Clemente, R.M. Salt stress alleviation in citrus plants by plant growth-promoting rhizobacteria Pseudomonas putida and Novosphingobium sp. Plant Cell Rep. 2018, 37, 1557–1569. [Google Scholar] [CrossRef]
- Guo, Y.B.; Zheng, H.; Yang, Y.L.; Wang, H.M. Characterization of Pseudomonas corrugata strain P94 isolated from soil in Beijing as a potential biocontrol agent. Curr. Microbiol. 2007, 55, 247–253. [Google Scholar] [CrossRef]
- Dincă, L.C.; Grenni, P.; Onet, C.; Onet, A. Fertilization and soil microbial community: A review. Appl. Sci. 2021, 12, 1198. [Google Scholar] [CrossRef]
- Yao, L.L.; Kong, L.; Yang, Q.L.; Nian, H.J.; Liang, J.P. Analysis of microbial diversity in rhizosphere soil of Panax notoginseng under different water and microbial fertilizer conditions. Agronomy 2024, 14, 922. [Google Scholar] [CrossRef]
- He, M.Q.; Zhao, R.L.; Liu, D.M.; Denchev, T.T.; Begerow, D.; Yurkow, A.; Kemler, M.; Millanes, A.M.; Wedin, A.; McTaggart, A.R.; et al. Species diversity of Basidiomycota. Fungal Divers. 2022, 114, 281–325. [Google Scholar] [CrossRef]
- Ozkilinc, H.; Sevinc, U. Molecular phylogenetic species in Alternaria pathogens infecting pistachio and wild relatives. 3 Biotech 2018, 8, 250. [Google Scholar] [CrossRef]
- Devi, S.; Sharma, M.; Manhas, R.K. Investigating the plant growth promoting and biocontrol potentiality of endophytic Streptomyces SP. SP5 against early blight in Solanum lycopersicum seedlings. BMC Microbiol. 2022, 22, 285. [Google Scholar] [CrossRef] [PubMed]
- Mangwende, E.; Kritzinger, Q.; Truter, M.; Aveling, T.A.S. Alternaria alternata: A new seed-transmitted disease of coriander in South Africa. Eur. J. Plant Pathol. 2018, 152, 409–416. [Google Scholar] [CrossRef]
- Zakaria, L. An overview of Aspergillus species associated with plant iseaseds. Pathogens 2024, 13, 813. [Google Scholar] [CrossRef]
- Tao, J.M.; Cao, P.J.; Xiao, Y.S.; Wang, Z.H.; Huang, Z.H.; Jin, J.J.; Liu, Y.J.; Yin, H.Q.; Liu, T.B.; Zhou, Z.C. Distribution of the potential pathogenic Alternaria on plant leaves determines foliar fungal communities around the disease spot. Environ. Res. 2021, 200, 111715. [Google Scholar] [CrossRef]
- Wang, Y.F.; Li, Y.Q.; Xie, Y.H.; Yang, X.M.; He, Z.Z.; Tian, H.; Duan, M.Y.; Tang, X.R.; Pan, S.G. Effects of nitrogen fertilizer rate under deep placement on grain yield and nitrogen use efficiency in mechanical pot-seedling transplanting rice. J. Plant Growth Regul. 2023, 42, 3100–3110. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, K.F.; Shi, C.; Li, X.Y.; Qiu, Z.L.; Shi, F.C. Responses of fungal assembly and c-occurrence network of rhizosphere soil to Amaranthus palmeri invasion in Northern China. J. Fungi 2023, 9, 509. [Google Scholar] [CrossRef]
- Du, L.; Zhong, S.N.; Luo, K.Y.; Yang, S.Q.; Xia, J.X.; Chen, Q. Effect of metal pollution on the distribution and co-occurrence pattern of bacterial, archaeal and fungal communities throughout the soil profiles. Chemosphere 2023, 315, 137692. [Google Scholar] [CrossRef] [PubMed]
- Yan, T.T.; Xue, J.H.; Zhou, Z.D.; Wu, Y.B. Biochar-based fertilizer amendments improve the soil microbial community structure in a karst mountainous area. Sci. Total Environ. 2021, 794, 148757. [Google Scholar] [CrossRef] [PubMed]
- Geneva, M.P.; Stancheva, I.V.; Boychinova, M.M.; Mincheva, N.H.; Yonova, P.A. Effects of foliar fertilization and arbuscular mycorrhizal colonization on Salvia officinalis L. growth, antioxidant capacity, and essential oil composition. J. Sci. Food Agric. 2010, 90, 696–702. [Google Scholar] [CrossRef]
- Li, D.H.; Qu, C.; Cheng, X.M.; Chen, Y.X.; Yan, H.; Wu, Q.N. Effect of different fertilization strategies on the yield, quality of Euryales Semen and soil microbial community. Front. Microbiol. 2023, 14, 1310366. [Google Scholar] [CrossRef]
- Dutta, A.; Dracatos, P.M.; Khan, G.A. Balancing act: The dynamic relationship between nutrient availability and plant defence. Plant J. 2024, 120, 1724–1734. [Google Scholar] [CrossRef] [PubMed]
- Cox, J.; Hue, N.V.; Ahmad, A.; Kobayashi, K.D. Surface-applied or incorporated biochar and compost combination improves soil fertility, Chinese cabbage and papaya biomass. Biochar 2021, 3, 213–227. [Google Scholar] [CrossRef]
- Xue, Y.B.; Zhu, S.N.; Schultz-Kraft, R.; Liu, G.D.; Chen, Z.J. Dissection of crop metabolome responses to nitrogen, phosphorus, potassium, and other nutrient deficiencies. Int. J. Mol. Sci. 2021, 23, 9079. [Google Scholar] [CrossRef]
- Abrol, V.; Sharma, P.; Chary, G.R.; Srinivasarao, C.; Maruthi Sankar, G.R.; Singh, B.; Kumar, A.; Hashem, A.; Ibrahimova, U.; Abd-Allah, E.F.; et al. Integrated organic and mineral fertilizer strategies for achieving sustainable maize yield and soil quality in dry sub-humid inceptisols. Sci. Rep. 2024, 14, 27227. [Google Scholar] [CrossRef]
- Carreón, Y.J.; Pereyra Zarate, A.A.; Pérez Sánchez, A.E. Surface coating with foliar fertilizers. Coatings 2024, 14, 1007. [Google Scholar] [CrossRef]
- Liu, G.M.; Zhang, X.C.; Wang, X.P.; Shao, H.B.; Yang, J.S.; Wang, X.P. Soil enzymes as indicators of saline soil fertility under various soil amendments. Agric. Ecosyst. Environ. 2017, 237, 274–279. [Google Scholar]
- Jabborova, D.; Sulaymanov, K.; Sayyed, R.Z.; Alotaibi, S.H.; Enakiev, Y.; Azimov, A.; Jabbarov, Z.; Ansari, M.J.; Fahad, S.; Danish, S.; et al. Mineral fertilizers improves the quality of turmeric and soil. Sustainability 2020, 13, 9437. [Google Scholar] [CrossRef]
- Voltr, V.; Menšík, L.; Hlisnikovský, L.; Hruška, M.; Pokorný, E.; Pospíšilová, L. The soil organic matter in connection with soil properties and soil inputs. Agronomy 2021, 11, 779. [Google Scholar] [CrossRef]
- Bai, X.; Liu, K.L.; Ning, T.; Deng, C.J.; Wang, L.; Li, D.Y.; Wang, T.; Li, J. Effects of multiple N, P, and K fertilizer combinations on strawberry growth and the microbial community. PLoS ONE 2023, 18, 0293088. [Google Scholar] [CrossRef]
- Hoffland, E.; Kuyper, T.W.; Comans, R.N.J.; Creamer, R.E. Eco-functionality of organic matter in soils. Plant Soil 2020, 455, 1–22. [Google Scholar] [CrossRef]
- Tao, J.H.; Chen, D.; Wu, S.F.; Zhang, Q.; Xiao, W.D.; Zhao, S.P.; Ye, X.Z.; Chu, T.F. The comprehensive effects of biochar amendments on soil organic carbon accumulation, soil acidification amelioration and heavy metal availability in the soil-rice system. Agronomy 2024, 14, 2498. [Google Scholar] [CrossRef]
- Zhou, D.J.; Mou, R.; Wang, L.H.; Liu, J.R.; Tang, Y.X.; Chen, J.; Heděnec, P.; Xu, Z.F.; Tan, B.; Cui, X.L.; et al. Fertilization effects on soil organic matter chemistry. Soil Tillage Res. 2025, 246, 106346. [Google Scholar] [CrossRef]
- Sarnaik, A.; Liu, A.R.; Nielsen, D.; Varman, A.M. High-throughput screening for efficient microbial biotechnology. Curr. Opin. Biotechnol. 2020, 64, 141–150. [Google Scholar] [CrossRef]
- Lei, Y.; Ding, D.; Duan, J.H.; Luo, Y.; Huang, F.Y.; Kang, Y.K.; Chen, Y.Y.; Li, S.J. Soil microbial community characteristics and their effect on tea quality under different fertilization treatments in two tea plantations. Genes 2024, 15, 610. [Google Scholar] [CrossRef] [PubMed]
- Liao, H.L.; Huang, L.; Li, N.; Ke, W.J.; Xiang, Y.Q.; Ma, Y.T. Auxiliary rapid identification of pathogenic and antagonistic microorganisms associated with Coptis chinensis root rot by high-throughput sequencing. Sci. Rep. 2021, 11, 11141. [Google Scholar] [CrossRef]
- Sun, A.Q.; Jiao, X.Y.; Chen, Q.L.; Wu, A.L.; Zheng, Y.; Lin, Y.X.; He, J.Z.; Hu, H.W. Microbial communities in crop phyllosphere and root endosphere are more resistant than soil microbiota to fertilization. Soil Biol. Biochem. 2021, 153, 108113. [Google Scholar] [CrossRef]
- Khalil, A.M.A.; Hassan, S.E.; Alsharif, S.M.; Eid, A.M.; Ewais, E.E.; Azab, E.; Gobouri, A.A.; Elkelish, A.; Fouda, A. Isolation and characterization of fungal endophytes isolated from medicinal plant Ephedra pachyclada as plant growt-promoting. Biomolecules 2021, 11, 140. [Google Scholar] [CrossRef]
- Mañero, F.J.G.; Algar, E.; Gómez, M.S.M.; Sierra, M.D.S.; Solano, B.R. Elicitation of secondary metabolism in Hypericum perforatum by rhizosphere bacteria and derived elicitors in seedlings and shoot cultures. Pharm. Biol. 2012, 50, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Gill, H.; Vasundhara, M. Isolation of taxol producing endophytic fungus Alternaria brassicicola from non-taxus medicinal plant Terminalia arjuna. World J. Microb. Biot. 2019, 35, 74. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.L.; Wang, S.J.; Wang, R.M.; Chen, Y.L.; Siddique, K.H.M.; Xia, G.M.; Chi, D.C. Ameliorative roles of biochar-based fertilizer on morpho-physiological traits, nutrient uptake and yield in Peanut (Arachis hypogaea L.) under water stress. Agric. Water Manag. 2021, 257, 107129. [Google Scholar] [CrossRef]
- Abramczyk, B.; Marzec-Grządizel, A.; Grządziel, J.; Król, E.; Galązka, A.; Oleszek, W. Biocontrol potential and catabolic profile of endophytic Diaporthe Eres strain 1420S from Prunus domestica L. in Poland-a preliminary study. Agronomy 2022, 12, 165. [Google Scholar] [CrossRef]
- Leff, J.W.; Del Tredici, P.; Friedman, W.E.; Fierer, N. Spatial structuring of bacterial communities within individual Ginkgo biloba trees. Environ. Microbiol. 2015, 17, 2352–2361. [Google Scholar] [CrossRef] [PubMed]
- Viana, D.M.; Negreiros, A.M.P.; da Silveira, L.M.; Cavalcante, A.L.A.; de Sousa Santos Alves, C.P.; Correia, K.C.; Júnior, R.S. Effects of inoculation of Macrophomina pseudophaseolina in Luffa spp. Trop. Plant Pathol. 2025, 50, 24. [Google Scholar]
- Xie, Y.L.; He, X.L.; Wang, D.; Wang, M.H.; Li, W.Y.; Chen, W.J.; Li, X.E.; He, C. Characterization of dark septate endophytes under drought and rehydration and their compensatory mechanisms in Astragalus membranaceus. Microorganisms 2024, 12, 2254. [Google Scholar] [CrossRef]
- Ichikawa, K.; Kawasaki, S.; Tanaka, C.; Tsuda, M. Induced resistance against Fusarium diseases of Cymbidium species by weakly virulent strain HPF-1 (Fusarium sp.). J. Gen. Plant Pathol. 2003, 69, 400–405. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.; Li, W.; Pan, Y.; Zheng, X.; Fu, X.; Wang, M.; Shi, W.; Wang, Z.; He, X.; He, C.; et al. Response of Endophytic Microbial Communities and Quality of Salvia miltiorrhiza to Fertilization Treatments. Microorganisms 2025, 13, 1429. https://doi.org/10.3390/microorganisms13061429
Chen W, Li W, Pan Y, Zheng X, Fu X, Wang M, Shi W, Wang Z, He X, He C, et al. Response of Endophytic Microbial Communities and Quality of Salvia miltiorrhiza to Fertilization Treatments. Microorganisms. 2025; 13(6):1429. https://doi.org/10.3390/microorganisms13061429
Chicago/Turabian StyleChen, Wenjing, Wanyun Li, Yangyang Pan, Xin Zheng, Xinxin Fu, Menghui Wang, Wenyi Shi, Zhenzhou Wang, Xueli He, Chao He, and et al. 2025. "Response of Endophytic Microbial Communities and Quality of Salvia miltiorrhiza to Fertilization Treatments" Microorganisms 13, no. 6: 1429. https://doi.org/10.3390/microorganisms13061429
APA StyleChen, W., Li, W., Pan, Y., Zheng, X., Fu, X., Wang, M., Shi, W., Wang, Z., He, X., He, C., & Li, X. (2025). Response of Endophytic Microbial Communities and Quality of Salvia miltiorrhiza to Fertilization Treatments. Microorganisms, 13(6), 1429. https://doi.org/10.3390/microorganisms13061429