
Impact of c-di-AMP Accumulation, L-cysteine, and Oxygen on Catalase Activity and Oxidative Stress Resistance of Listeria monocytogenes 10403S

Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Survival in the Presence of Hydrogen Peroxide
2.3. Catalase Activity Assay
2.4. Measurement of Intracellular c-di-AMP Levels
2.5. RNA Sequencing (RNA-seq) Sample Preparation and Analysis of L. monocytogenes Strains
2.6. Statistical Analysis
3. Results
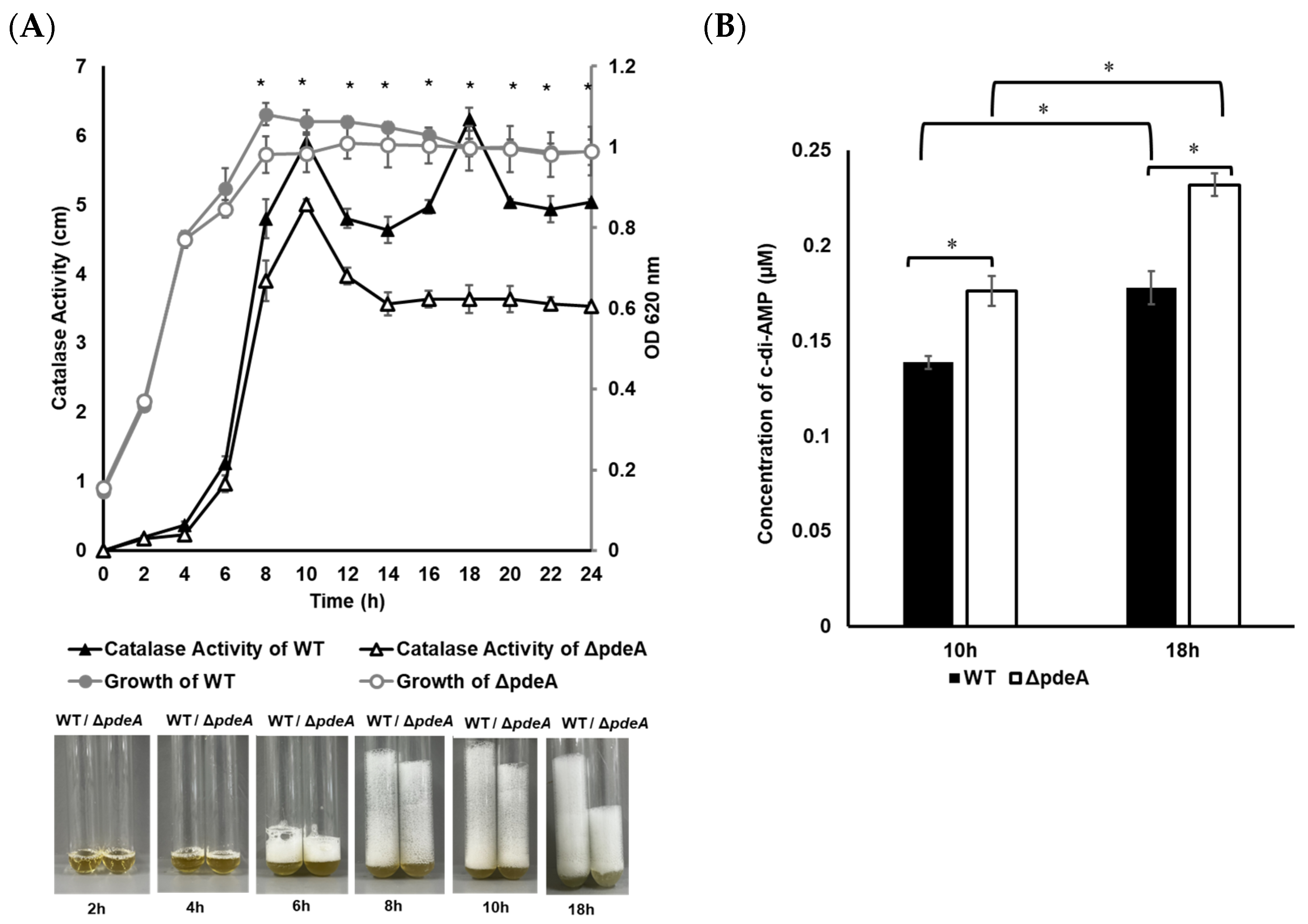
3.1. Catalase Activity Levels During Growth
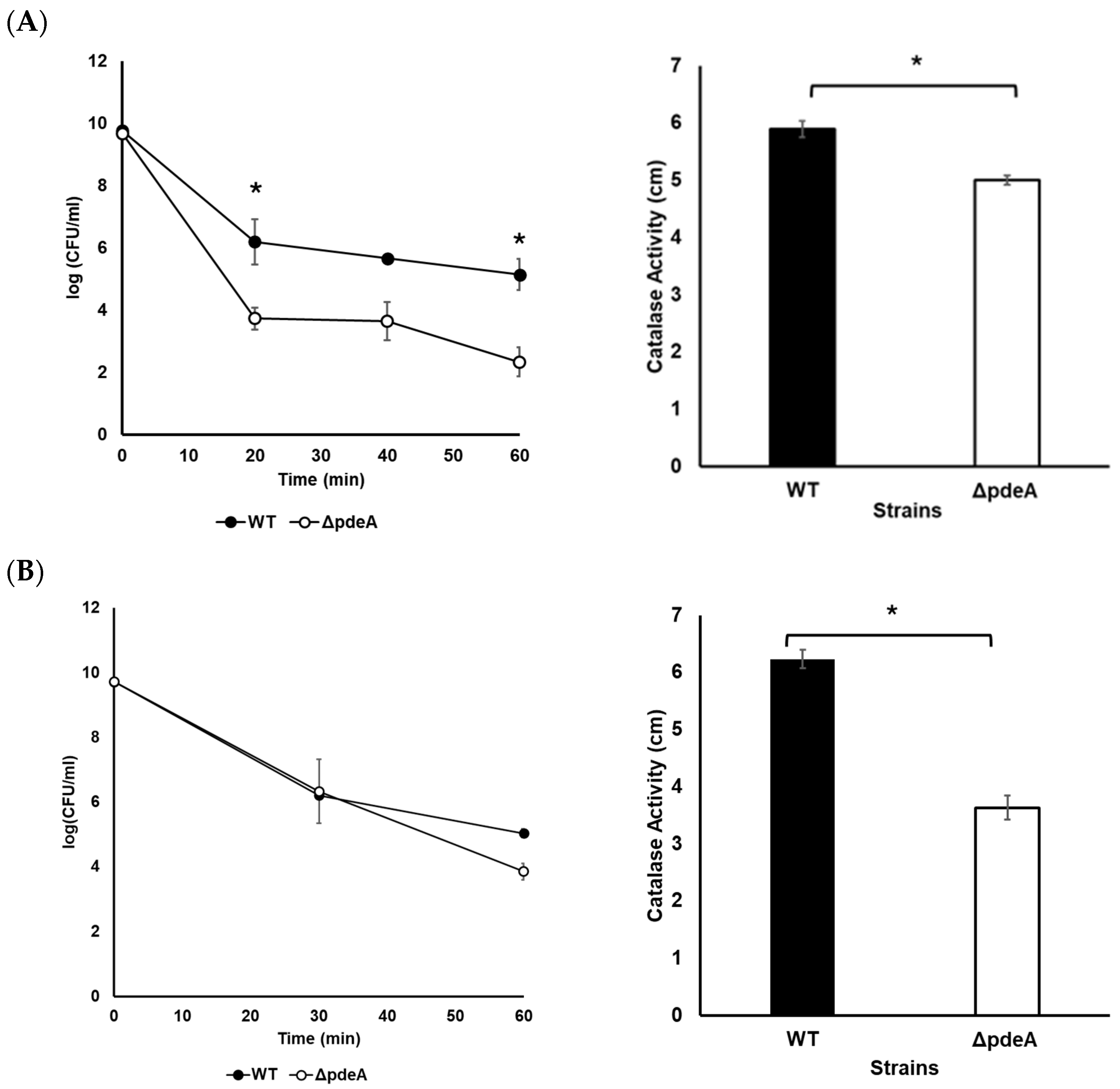
3.2. Survival Against H2O2
3.3. Intracellular c-di-AMP Concentrations During the H2O2 Treatment
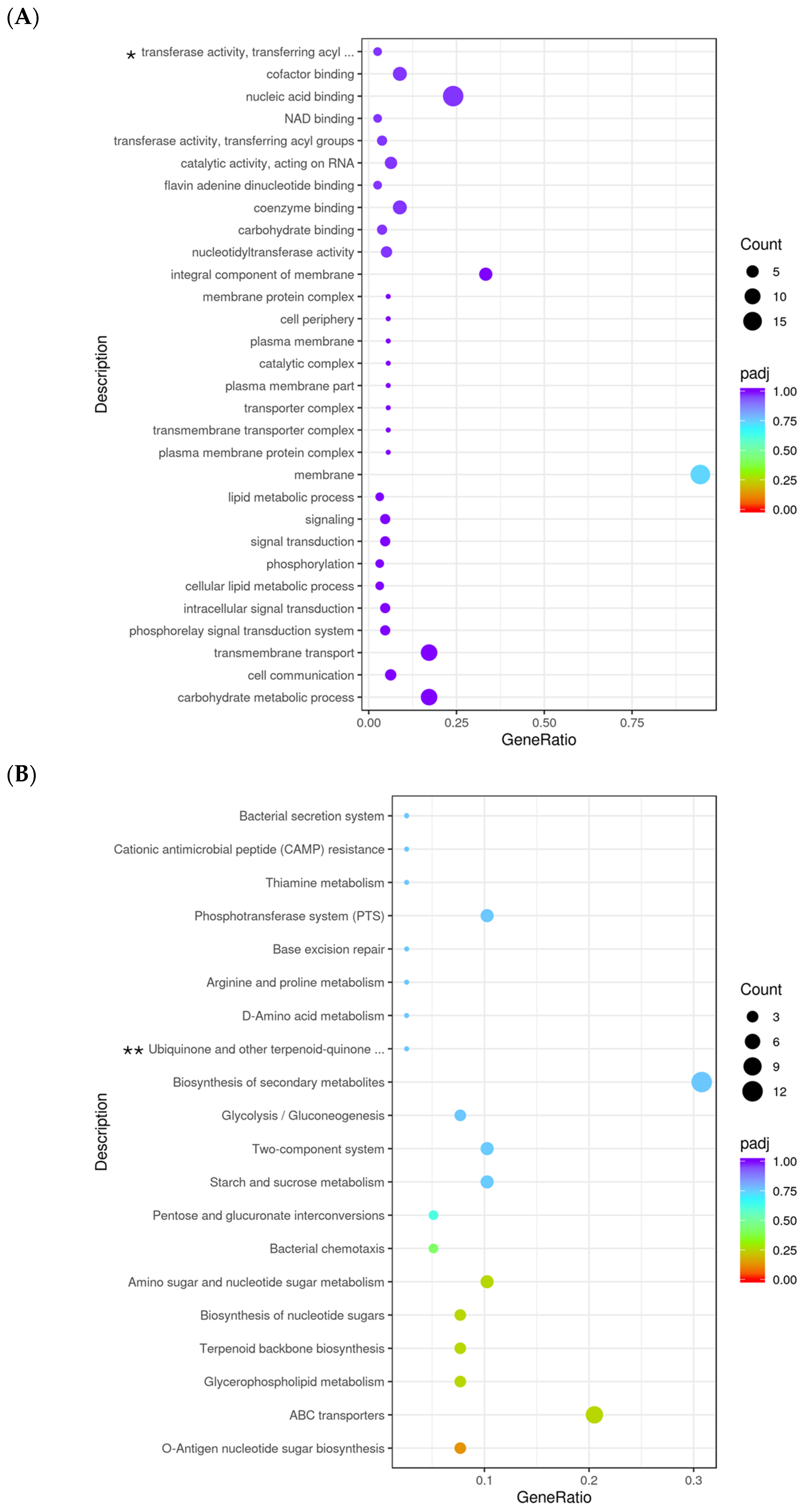
3.4. Transcriptomic Analysis Before the Stress Condition
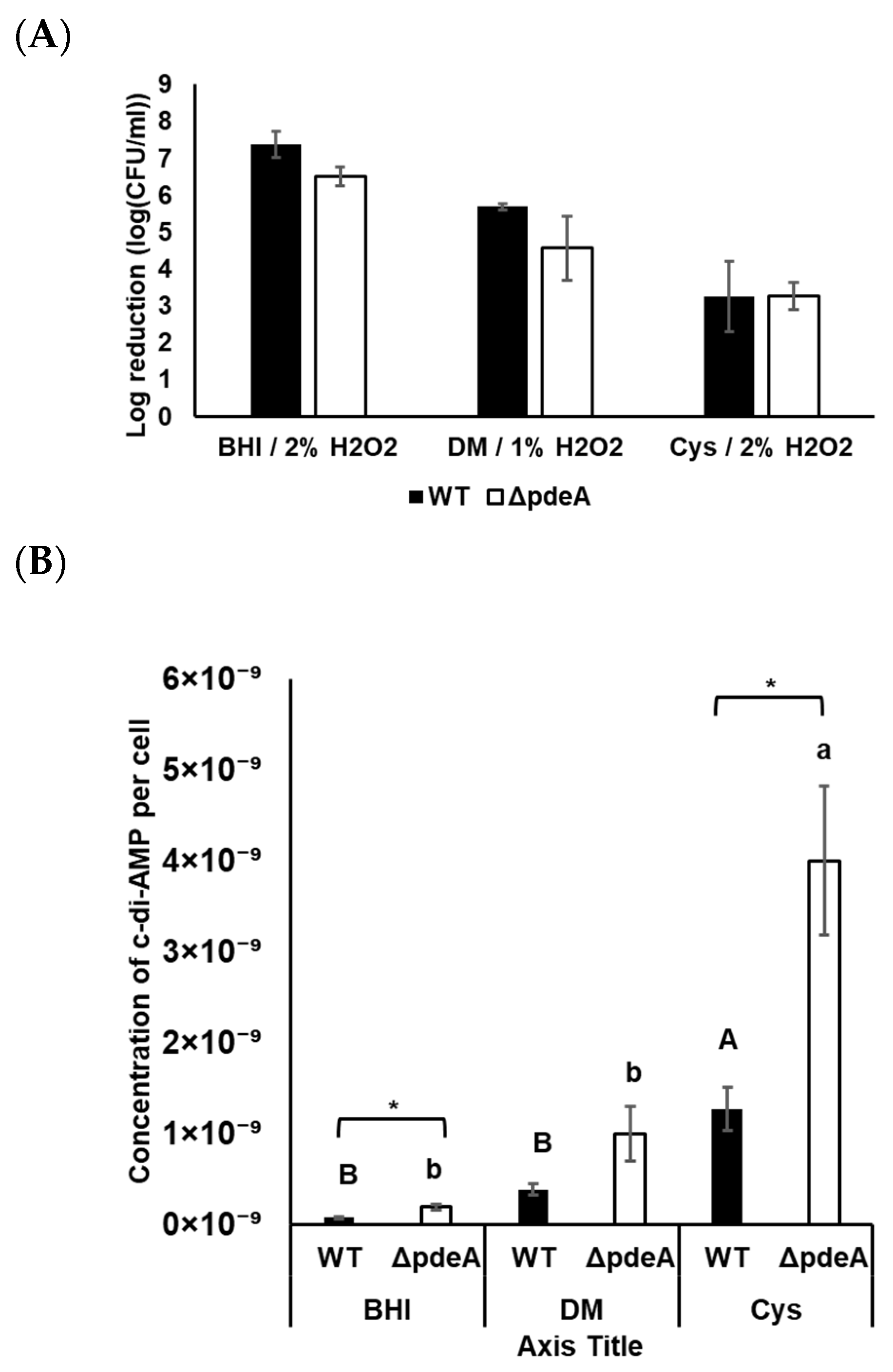
3.5. The Effect of L-cysteine on the Catalase Activity and Survival of WT and ΔpdeA Under Aerobic Conditions
3.6. Intracellular c-di-AMP Concentrations Under Aerobic Conditions
3.7. Survival and Catalase Activity of Anaerobically Grown WT and ΔpdeA Cells
3.8. Intracellular c-di-AMP Concentrations Under Anaerobic Conditions
3.9. Transcriptomic Data Showing the Effect of L-cysteine Supplementation on c-di-AMP Homeostasis Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATP | Adenosine Triphosphate |
| BHI | Brain Heart Infusion |
| BP | Biological Processes |
| c-di-AMP | Cyclic di-AMP |
| CC | Cellular Compound |
| DACs | Diadenylate Cyclases |
| DM | Defined Media |
| ESI | Electrospray Ionisation |
| GO | Gene Ontology |
| HPLC | High-Performance Liquid Chromatography |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| LC-MS | Liquid Chromatography–Mass Spectrometry |
| MF | Molecular Function |
| PDEs | Phosphodiesterases |
| PPP | Pentose Phosphate Pathway |
| ROS | Reactive Oxygen Species |
| SOD | Superoxide Dismutase |
| TCA | Tricarboxylic Acid Cycle |
| WT | Wild Type |
References
- Lungu, B.; Ricke, S.C.; Johnson, M.G. Growth, survival, proliferation and pathogenesis of Listeria monocytogenes under low oxygen or anaerobic conditions: A review. Anaerobe 2009, 15, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Pizarro-Cerdá, J.; Cossart, P. Listeria monocytogenes: Cell biology of invasion and intracellular growth. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Matereke, L.T.; Okoh, A.I. Listeria monocytogenes virulence, antimicrobial resistance and environmental persistence: A review. Pathogens 2020, 9, 528. [Google Scholar] [CrossRef] [PubMed]
- White, S.J.; McClung, D.M.; Wilson, J.G.; Roberts, B.N.; Donaldson, J.R. Influence of pH on bile sensitivity amongst various strains of Listeria monocytogenes under aerobic and anaerobic conditions. J. Med. Microbiol. 2015, 64, 1287–1296. [Google Scholar] [CrossRef] [PubMed]
- Mongkolsuk, S.; Helmann, J.D. Regulation of inducible peroxide stress responses. Mol. Microbiol. 2002, 45, 9–15. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Kettle, A.J. Redox reactions and microbial killing in the neutrophil phagosome. Antioxid. Redox Signal. 2013, 18, 642–660. [Google Scholar] [CrossRef]
- Müller-Herbst, S.; Wüstner, S.; Mühlig, A.; Eder, D.; Fuchs, T.M.; Held, C.; Ehrenreich, A.; Scherer, S. Identification of genes essential for anaerobic growth of Listeria monocytogenes. Microbiology 2014, 160, 752–765. [Google Scholar] [CrossRef]
- Whiteley, A.T.; Pollock, A.J.; Portnoy, D.A. The PAMP c-di-AMP is essential for Listeria monocytogenes growth in rich but not minimal media due to a toxic increase in (p)ppGpp. Cell Host Microbe 2015, 17, 788–798. [Google Scholar] [CrossRef]
- Bo Andersen, J.; Roldgaard, B.B.; Christensen, B.B.; Licht, T.R. Oxygen restriction increases the infective potential of Listeria monocytogenes in vitro in Caco-2 cells and in vivo in guinea pigs. BMC Microbiol. 2007, 7, 55. [Google Scholar] [CrossRef]
- Jydegaard-Axelsen, A.M.; Høiby, P.E.; Holmstrøm, K.; Russell, N.; Knøchel, S. CO2- and anaerobiosis-induced changes in physiology and gene expression of different Listeria monocytogenes strains. Appl. Environ. Microbiol. 2004, 70, 4111–4117. [Google Scholar] [CrossRef]
- Flint, A.; Butcher, J.; Stintzi, A. Stress responses, adaptation, and virulence of bacterial pathogens during host gastrointestinal colonization. Microbiol. Spectrum. 2016, 4, VMBF-0007-2015. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, R.M.; Gründling, A. Cyclic di-AMP: Another second messenger enters the fray. Nat. Rev. Microbiol. 2013, 11, 513–524. [Google Scholar] [CrossRef]
- Fahmi, T.; Port, G.C.; Cho, K.H. c-di-AMP: An essential molecule in the signaling pathways that regulate the viability and virulence of Gram-positive bacteria. Genes 2017, 8, 197. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Z.Q.; Fan, Y.Z.; Song, X.; Liu, X.X.; Xia, Y.J.; Ai, L.Z. The second messenger c-di-AMP mediates bacterial exopolysaccharide biosynthesis: A review. Mol. Biol. Rep. 2020, 47, 7811–7817. [Google Scholar] [CrossRef] [PubMed]
- Huynh, T.A.N.; Luo, S.; Pensinger, D.; Sauer, J.D.; Tong, L.; Woodward, J.J. An HD-domain phosphodiesterase mediates cooperative hydrolysis of c-di-AMP to affect bacterial growth and virulence. Proc. Natl. Acad. Sci. USA 2015, 112, E747–E756. [Google Scholar] [CrossRef]
- Abundiz-Yañez, K.; Leyva-Sánchez, H.C.; Robleto, E.A.; Pedraza-Reyes, M. Stress-associated and growth-dependent mutagenesis are divergently regulated by c-di-AMP levels in Bacillus subtilis. Int. J. Mol. Sci. 2023, 24, 455. [Google Scholar] [CrossRef]
- Rallu, F.; Gruss, A.; Ehrlich, S.D.; Maguin, E. Acid- and multistress-resistant mutants of Lactococcus lactis: Identification of intracellular stress signals. Mol. Microbiol. 2000, 35, 517–528. [Google Scholar] [CrossRef]
- Zarrella, T.M.; Metzger, D.W.; Bai, G. Stress suppressor screening leads to detection of regulation of cyclic di-AMP homeostasis by a Trk family effector protein in Streptococcus pneumoniae. J. Bacteriol. 2018, 200, e00045-18. [Google Scholar] [CrossRef]
- Rørvik, G.H.; Naemi, A.O.; Edvardsen, P.K.T.; Simm, R. The c-di-AMP signaling system influences stress tolerance and biofilm formation of Streptococcus mitis. MicrobiologyOpen 2021, 10, e1203. [Google Scholar] [CrossRef]
- Dengler Haunreiter, V.; Tarnutzer, A.; Bär, J.; von Matt, M.; Hertegonne, S.; Andreoni, F.; Vulin, C.; Künzi, L.; Menzi, C.; Kiefer, P.; et al. c-di-AMP levels modulate Staphylococcus aureus cell wall thickness, response to oxidative stress, and antibiotic resistance and tolerance. Microbiol. Spectr. 2023, 11, e02788-23. [Google Scholar] [CrossRef]
- Gándara, C.; Alonso, J.C. DisA and c-di-AMP act at the intersection between DNA-damage response and stress homeostasis in exponentially growing Bacillus subtilis cells. DNA Repair 2015, 27, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Rubin, B.E.; Huynh, T.N.; Welkie, D.G.; Diamond, S.; Simkovsky, R.; Pierce, E.C.; Taton, A.; Lowe, L.C.; Lee, J.J.; Rifkin, S.A.; et al. High-throughput interaction screens illuminate the role of c-di-AMP in cyanobacterial nighttime survival. PLoS Genet. 2018, 14, e1007301. [Google Scholar] [CrossRef]
- Clauditz, A.; Resch, A.; Wieland, K.P.; Peschel, A.; Götz, F. Staphyloxanthin plays a role in the fitness of Staphylococcus aureus and its ability to cope with oxidative stress. Infect. Immun. 2006, 74, 4950–4953. [Google Scholar] [CrossRef]
- Cheng, X.; Zheng, X.; Zhou, X.; Zeng, J.; Ren, Z.; Xu, X.; Cheng, L.; Li, M.; Li, J.; Li, Y. Regulation of oxidative response and extracellular polysaccharide synthesis by a diadenylate cyclase in Streptococcus mutans. Environ. Microbiol. 2016, 18, 904–922. [Google Scholar] [CrossRef]
- Fahmi, T.; Faozia, S.; Port, G.C.; Cho, K.H. The second messenger c-di-AMP regulates diverse cellular pathways involved in stress response, biofilm formation, cell wall homeostasis, SpeB expression, and virulence in Streptococcus pyogenes. Infect. Immun. 2019, 87, e00571-18. [Google Scholar] [CrossRef] [PubMed]
- Walker, M.C.; van der Donk, W.A. The many roles of glutamate in metabolism. J. Ind. Microbiol. Biotechnol. 2016, 43, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Zeden, M.S.; Kviatkovski, I.; Schuster, C.F.; Thomas, V.C.; Fey, P.D.; Gründling, A. Identification of the main glutamine and glutamate transporters in Staphylococcus aureus and their impact on c-di-AMP production. Mol. Microbiol. 2020, 113, 1085–1100. [Google Scholar] [CrossRef]
- Gundlach, J.; Mehne, F.M.; Herzberg, C.; Kampf, J.; Valerius, O.; Kaever, V.; Stülke, J. An essential poison: Synthesis and degradation of cyclic di-AMP in Bacillus subtilis. J. Bacteriol. 2015, 197, 3265–3274. [Google Scholar] [CrossRef]
- Sato, Y.; Noji, S.; Suzuki, R.; Taniguchi, S. Dual mechanism for stimulation of glutamate transport by potassium ions in Streptococcus mutans. J. Bacteriol. 1989, 171, 4963–4966. [Google Scholar] [CrossRef]
- Krastel, K.; Senadheera, D.B.; Mair, R.; Downey, J.S.; Goodman, S.D.; Cvitkovitch, D.G. Characterization of a glutamate transporter operon, glnQHMP, in Streptococcus mutans and its role in acid tolerance. J. Bacteriol. 2010, 192, 984–993. [Google Scholar] [CrossRef]
- Gibhardt, J.; Hoffmann, G.; Turdiev, A.; Wang, M.; Lee, V.T.; Commichau, F.M. C-di-AMP assists osmoadaptation by regulating the Listeria monocytogenes potassium transporters KimA and KtrCD. J. Biol. Chem. 2019, 294, 16020–16033. [Google Scholar] [CrossRef]
- Sauer, J.D.; Herskovits, A.A.; O’Riordan, M.X. Metabolism of the gram-positive bacterial pathogen Listeria monocytogenes. Microbiol. Spectr. 2019, 7, GPP3-0066-2019. [Google Scholar] [CrossRef] [PubMed]
- Azizoglu, R.O.; Kathariou, S. Temperature-dependent requirement for catalase in aerobic growth of Listeria monocytogenes F2365. Appl. Environ. Microbiol. 2010, 76, 6998–7003. [Google Scholar] [CrossRef] [PubMed]
- Iwase, T.; Tajima, A.; Sugimoto, S.; Okuda, K.I.; Hironaka, I.; Kamata, Y.; Takada, K.; Mizunoe, Y. A simple assay for measuring catalase activity: A visual approach. Sci. Rep. 2013, 3, 3081. [Google Scholar] [CrossRef] [PubMed]
- Amezaga, M.R.; Davidson, I.; McLaggan, D.; Verheul, A.; Abee, T.; Booth, I.R. The role of peptide metabolism in the growth of Listeria monocytogenes ATCC 23074 at high osmolarity. Microbiology 1995, 141, 41–49. [Google Scholar] [CrossRef]
- Karatzas, K.A.G.; Brennan, O.; Heavin, S.; Morrissey, J.; O’Byrne, C.P. Intracellular accumulation of high levels of γ-aminobutyrate by Listeria monocytogenes 10403S in response to low pH: Uncoupling of γ-aminobutyrate synthesis from efflux in a chemically defined medium. Appl. Environ. Microbiol. 2010, 76, 3529–3537. [Google Scholar] [CrossRef]
- Witte, C.E.; Whiteley, A.T.; Burke, T.P.; Sauer, J.D.; Portnoy, D.A.; Woodward, J.J. Cyclic di-AMP is critical for Listeria monocytogenes growth, cell wall homeostasis, and establishment of infection. mBio 2013, 4, e00282-13. [Google Scholar] [CrossRef]
- Oppenheimer-Shaanan, Y.; Wexselblatt, E.; Katzhendler, J.; Yavin, E.; Ben-Yehuda, S. C-di-AMP reports DNA integrity during sporulation in Bacillus subtilis. EMBO Rep. 2011, 12, 594–601. [Google Scholar] [CrossRef]
- Mathews, P.G. Design of Experiments with MINITAB; Quality Press: Milwaukee, WI, USA, 2004. [Google Scholar]
- Ruhland, B.R.; Reniere, M.L. Sense and sensor ability: Redox-responsive regulators in Listeria monocytogenes. Curr. Opin. Microbiol. 2019, 47, 20–25. [Google Scholar] [CrossRef]
- Siletti, C.; Freeman, M.; Tu, Z.; Stevenson, D.M.; Amador-Noguez, D.; Sauer, J.D.; Huynh, T.N. C-di-AMP accumulation disrupts glutathione metabolism and inhibits virulence program expression in Listeria monocytogenes. bioRxiv 2024. [Google Scholar] [CrossRef]
- Muchaamba, F.; Stephan, R.; Tasara, T. Listeria monocytogenes cold shock proteins: Small proteins with a huge impact. Microorganisms 2021, 9, 1061. [Google Scholar] [CrossRef]
- Díaz-Acosta, A.; Sandoval, M.L.; Delgado-Olivares, L.; Membrillo-Hernández, J. Effect of anaerobic and stationary phase growth conditions on the heat shock and oxidative stress responses in Escherichia coli K-12. Arch. Microbiol. 2006, 185, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Boura, M.; Keating, C.; Royet, K.; Paudyal, R.; O’Donoghue, B.; O’Byrne, C.P.; Karatzas, K.A. Loss of SigB in Listeria monocytogenes Strains EGD-e and 10403S Confers Hyperresistance to Hydrogen Peroxide in Stationary Phase under Aerobic Conditions. Appl. Environ. Microbiol. 2016, 82, 4584–4591. [Google Scholar] [CrossRef] [PubMed]
- Yilmaz Topcam, M.M.; Karatzas, K.A.G. Evaluating the Role of Cysteine Transporter ctaP (lmo0135) and Extracellular Cysteine in Stationary Phase-Grown Listeria monocytogenes 10403S by Transcriptional Analysis; University of Reading: Reading, UK, 2025. [Google Scholar]
- Corrigan, R.M.; Bowman, L.; Willis, A.R.; Kaever, V.; Gründling, A. Cross-talk between two nucleotide-signaling pathways in Staphylococcus aureus. J. Biol. Chem. 2015, 290, 5826–5839. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, B.J.; Hinshelwood, C.N. Variations in catalase activity during a bacterial growth cycle. Proc. R. Soc. London. Ser. B-Biol. Sci. 1959, 150, 13–23. [Google Scholar]
- Peterson, B.N.; Young, J.W.; Smith, S.M.; Sham, L.-T.; Winkler, M.E.; Grayczyk, J.P.; Jones-Carson, J.; Slauch, J.M. (P)ppGpp and c-di-AMP homeostasis is controlled by Cbpb in Listeria monocytogenes. mBio 2020, 11, e01625-20. [Google Scholar] [CrossRef]
- Liu, K.; Bittner, A.N.; Wang, J.D. Diversity in (p)ppGpp metabolism and effectors. Curr. Opin. Microbiol. 2015, 24, 72–79. [Google Scholar] [CrossRef]
- Rao, F.; See, R.Y.; Zhang, D.; Toh, D.C.; Ji, Q.; Liang, Z.X. YybT is a signaling protein that contains a cyclic dinucleotide phosphodiesterase domain and a GGDEF domain with ATPase activity. J. Biol. Chem. 2010, 285, 473–482. [Google Scholar] [CrossRef]
- Mains, D.R.; Eallonardo, S.J.; Freitag, N.E. Identification of Listeria monocytogenes genes contributing to oxidative stress resistance under conditions relevant to host infection. Infect. Immun. 2021, 89, e00112-21. [Google Scholar] [CrossRef]
- Portman, J.L.; Dubensky, S.B.; Peterson, B.N.; Whiteley, A.T.; Portnoy, D.A. Activation of the Listeria monocytogenes virulence program by a reducing environment. mBio 2017, 8, e01054-17. [Google Scholar] [CrossRef]
- Sureka, K.; Choi, P.H.; Precit, M.; Delince, M.; Pensinger, D.A.; Huynh, T.N.; Jurado, A.R.; Goo, Y.A.; Sadilek, M.; Iavarone, A.T.; et al. The Cyclic Dinucleotide c-di-AMP Is an Allosteric Regulator of Metabolic Enzyme Function. Cell 2014, 158, 1389–1401. [Google Scholar] [CrossRef]
- Ostrovsky, D.; Diomina, G.; Lysak, E.; Matveeva, E.; Ogrel, O.; Trutko, S. Effect of Oxidative Stress on the Biosynthesis of 2-C-Methyl-D-Erythritol-2, 4-Cyclopyrophosphate and Isoprenoids by Several Bacterial Strains. Arch. Microbiol. 1998, 171, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Frank, A.; Groll, M. The methylerythritol phosphate pathway to isoprenoids. Chem. Rev. 2017, 117, 5675–5703. [Google Scholar] [CrossRef]
- Gray, M.J.; Jakob, U. Oxidative stress protection by polyphosphate—New roles for an old player. Mol. Microbiol. 2015, 95, 497–504. [Google Scholar] [CrossRef]
- Pan, Y.; Cheng, J.H.; Sun, D.W. Metabolomic analyses on microbial primary and secondary oxidative stress responses. Compr. Rev. Food Sci. Food Saf. 2021, 20, 5675–5697. [Google Scholar] [CrossRef] [PubMed]
- Muchaamba, F.; Eshwar, A.K.; Stevens, M.J.A.; von Ah, U.; Tasara, T. Variable carbon source utilization, stress resistance, and virulence profiles among Listeria monocytogenes strains responsible for listeriosis outbreaks in Switzerland. Front. Microbiol. 2019, 10, 957. [Google Scholar] [CrossRef]
- Stincone, A.; Prigione, A.; Cramer, T.; Wamelink, M.M.; Campbell, K.; Cheung, E.; Olin-Sandoval, V.; Grüning, N.M.; Krüger, A.; Tauqeer Alam, M.; et al. The return of metabolism: Biochemistry and physiology of the pentose phosphate pathway. Biol. Rev. 2015, 90, 927–963. [Google Scholar] [CrossRef]
- He, L.; Deng, Q.L.; Chen, M.T.; Wu, Q.P.; Lu, Y.J. Proteomics analysis of Listeria monocytogenes ATCC 19115 in response to simultaneous triple stresses. Arch. Microbiol. 2015, 197, 833–841. [Google Scholar] [CrossRef] [PubMed]
- Abdelhamed, H.; Ramachandran, R.; Narayanan, L.; Islam, S.; Ozan, O.; Freitag, N.; Lawrence, M.L. Role of FruR transcriptional regulator in virulence of Listeria monocytogenes and identification of its regulon. PLoS ONE 2022, 17, e0274005. [Google Scholar] [CrossRef]
- Chandel, N.S. NADPH—The forgotten reducing equivalent. Cold Spring Harb. Perspect. Biol. 2021, 13, a040550. [Google Scholar] [CrossRef]
- Ogunleye, S.C.; Islam, S.; Chowdhury, Q.M.K.; Ozdemir, O.; Lawrence, M.L.; Abdelhamed, H. Catabolite Control Protein C Contributes to Virulence and Hydrogen Peroxide-Induced Oxidative Stress Responses in Listeria monocytogenes. Front. Microbiol. 2024, 15, 1403694. [Google Scholar] [CrossRef] [PubMed]
- Pittman, J.R.; Buntyn, J.O.; Posadas, G.; Nanduri, B.; Pendarvis, K.; Donaldson, J.R. Proteomic analysis of cross protection provided between cold and osmotic stress in Listeria monocytogenes. J. Proteome Res. 2014, 13, 1896–1904. [Google Scholar] [CrossRef]
- Bowman, J.P.; Hages, E.; Nilsson, R.E.; Kocharunchitt, C.; Ross, T. Investigation of the Listeria monocytogenes Scott A acid tolerance response and associated physiological and phenotypic features via whole proteome analysis. J. Proteome Res. 2012, 11, 2409–2426. [Google Scholar] [CrossRef]
- Rosato, R.R.; Fernandez, R.; Paz, L.I.; Singh, C.R.; Rosato, A.E. TCA cycle-mediated generation of ROS is a key mediator for HeR-MRSA survival under β-lactam antibiotic exposure. PLoS ONE 2014, 9, e99605. [Google Scholar] [CrossRef] [PubMed]
- Chueca, B.; Pagán, R.; García-Gonzalo, D. Oxygenated monoterpenes citral and carvacrol cause oxidative damage in Escherichia coli without the involvement of tricarboxylic acid cycle and Fenton reaction. Int. J. Food Microbiol. 2014, 189, 126–131. [Google Scholar] [CrossRef]
- Cortes, B.W.; Naditz, A.L.; Anast, J.M.; Schmitz-Esser, S. Transcriptome sequencing of Listeria monocytogenes reveals major gene expression changes in response to lactic acid stress exposure but a less pronounced response to oxidative stress. Front. Microbiol. 2020, 10, 3110. [Google Scholar] [CrossRef] [PubMed]
- Tu, Z.; Stevenson, D.M.; McCaslin, D.; Amador-Noguez, D.; Huynh, T.N. The Role of Listeria monocytogenes PstA in β-Lactam Resistance Requires the Cytochrome bd Oxidase Activity. J. Bacteriol. 2024, 206, e00130-24. [Google Scholar] [CrossRef]
- Kalynych, S.; Morona, R.; Cygler, M. Progress in understanding the assembly process of bacterial O-antigen. FEMS Microbiol. Rev. 2014, 38, 1048–1065. [Google Scholar] [CrossRef]
- Tian, B.; Xu, D.; Li, W.; Wang, J.; Cheng, J.; Liu, Y. Proteomic analysis of hexahydro-β-acids/hydroxypropyl-β-cyclodextrin inhibit Listeria monocytogenes. Appl. Microbiol. Biotechnol. 2022, 106, 755–771. [Google Scholar] [CrossRef]
- Bhat, S.V.; Booth, S.C.; Vantomme, E.A.; Afroj, S.; Yost, C.K.; Dahms, T.E. Oxidative stress and metabolic perturbations in Escherichia coli exposed to sublethal levels of 2, 4-dichlorophenoxyacetic acid. Chemosphere 2015, 135, 453–461. [Google Scholar] [CrossRef]
- Liao, X.; Liu, D.; Xiang, Q.; Ahn, J.; Chen, S.; Ye, X.; Ding, T. Inactivation Mechanisms of Non-Thermal Plasma on Microbes: A Review. Food Control 2017, 75, 83–91. [Google Scholar] [CrossRef]
- Avalos, M.; Garbeva, P.; Vader, L.; van Wezel, G.P.; Dickschat, J.S.; Ulanova, D. Biosynthesis, evolution and ecology of microbial terpenoids. Nat. Prod. Rep. 2022, 39, 249–272. [Google Scholar] [CrossRef] [PubMed]
- Guidea, A.; Zăgrean-Tuza, C.; Moț, A.C.; Sârbu, C. Comprehensive evaluation of radical scavenging, reducing power and chelating capacity of free proteinogenic amino acids using spectroscopic assays and multivariate exploratory techniques. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2020, 233, 118158. [Google Scholar] [CrossRef] [PubMed]
- Göbbels, L.; Poehlein, A.; Dumnitch, A.; Egelkamp, R.; Kröger, C.; Haerdter, J.; Hackl, T.; Feld, A.; Weller, H.; Daniel, R.; et al. Cysteine: An overlooked energy and carbon source. Sci. Rep. 2021, 11, 2139. [Google Scholar] [CrossRef]
- Cesinger, M.R.; Thomason, M.K.; Edrozo, M.B.; Halsey, C.R.; Reniere, M.L. Listeria monocytogenes SpxA1 Is a Global Regulator Required to Activate Genes Encoding Catalase and Heme Biosynthesis Enzymes for Aerobic Growth. Mol. Microbiol. 2020, 114, 230–243. [Google Scholar] [CrossRef]
- Saleh, E.; Sharkawy, I.S. The Impact of Growth History on Stress Robustness of Listeria monocytogenes. Clin. Investig. 2022, 12, 276–284. [Google Scholar]
- Roberts, B.N.; Chakravarty, D.; Gardner, J.C., III; Ricke, S.C.; Donaldson, J.R. Listeria monocytogenes Response to Anaerobic Environments. Pathogens 2020, 9, 210. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Relevant Properties | Reference Source |
|---|---|---|
| 10403S | Serotype ½ a, wild type, isolated from skin lesion | [36] |
| 10403S ΔpdeA | In-frame deletion (knockout) | [37] |
| Gene Name | Locus Tag | Log2fold Change | p-Value | p-adj | Gene Description |
|---|---|---|---|---|---|
| Novel | NI | 1.12 | 1.14 × 10−5 | 1.58 × 10−5 | cyclic-di-AMP receptor |
| pdeA | lmo0052 | −0.77 | 5.11 × 10−10 | 8.54 × 10−10 | cyclic-di-AMP phosphodiesterase |
| pgpH | lmo1466 | −2.34 | 1.19 × 10−139 | 4.23 × 10−138 | cyclic-di-AMP phosphodiesterase |
| dacA | lmo2120 | −3.16 | 1.48 × 10−109 | 3.13 × 10−108 | Diadenylate cyclase |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yilmaz Topcam, M.M.; Balagiannis, D.P.; Karatzas, K.A.G. Impact of c-di-AMP Accumulation, L-cysteine, and Oxygen on Catalase Activity and Oxidative Stress Resistance of Listeria monocytogenes 10403S. Microorganisms 2025, 13, 1400. https://doi.org/10.3390/microorganisms13061400
Yilmaz Topcam MM, Balagiannis DP, Karatzas KAG. Impact of c-di-AMP Accumulation, L-cysteine, and Oxygen on Catalase Activity and Oxidative Stress Resistance of Listeria monocytogenes 10403S. Microorganisms. 2025; 13(6):1400. https://doi.org/10.3390/microorganisms13061400
Chicago/Turabian StyleYilmaz Topcam, Mahide Muge, Dimitrios P. Balagiannis, and Kimon Andreas G. Karatzas. 2025. "Impact of c-di-AMP Accumulation, L-cysteine, and Oxygen on Catalase Activity and Oxidative Stress Resistance of Listeria monocytogenes 10403S" Microorganisms 13, no. 6: 1400. https://doi.org/10.3390/microorganisms13061400
APA StyleYilmaz Topcam, M. M., Balagiannis, D. P., & Karatzas, K. A. G. (2025). Impact of c-di-AMP Accumulation, L-cysteine, and Oxygen on Catalase Activity and Oxidative Stress Resistance of Listeria monocytogenes 10403S. Microorganisms, 13(6), 1400. https://doi.org/10.3390/microorganisms13061400