Abstract
The genome of Geobacillus sp. G4, a thermophilic bacterium isolated from a geothermal field in Peru, was sequenced and analyzed to evaluate its taxonomic and biotechnological potential. This strain exhibits optimal growth at temperatures between 50 and 70 °C and at a pH range of 6.0–7.5. Phenotypic assays demonstrated extracellular enzymatic activities, including amylases, cellulases, pectinases, and xylanases, highlighting its potential for efficient polysaccharide degradation. The assembled genome comprises approximately 3.4 Mb with a G+C content of 52.59%, containing 3,490 genes, including coding sequences, rRNAs, and tRNAs. Functional annotation revealed genes associated with key metabolic pathways such as glycogen and trehalose biosynthesis, indicating adaptation to carbohydrate-rich environments. Phylogenetic analyses based on ANI and dDDH values identified Geobacillus thermoleovorans KCTC3570 as its closest relative, suggesting a strong evolutionary relationship. Additionally, the genome harbors gene clusters for secondary metabolites such as betalactone and fengycin, suggesting potential industrial and pharmaceutical applications, including bioremediation. The identification of antibiotic resistance genes, specifically those conferring glycopeptide resistance, underscores their relevance for antimicrobial resistance studies. The presence of enzymes like amylases and pullulanase further emphasizes its biotechnological potential, particularly in starch hydrolysis and biofuel production. Overall, this research highlights the significant potential of Geobacillus species as valuable sources of thermostable enzymes and biosynthetic pathways for industrial applications.
1. Introduction
The genus Geobacillus comprises thermophilic, Gram-positive, spore-forming, rod-shaped bacteria that are either aerobic or facultatively anaerobic, constituting a phylogenetically coherent clade within the family Bacillaceae [1]. These microorganisms inhabit a wide range of environments, including hot springs, cold regions, and ocean sediments [2]. The genus Geobacillus was proposed by Nazina et al. (2001) [3] to differentiate it from the genus Bacillus based on distinct physiological traits, variations in growth temperatures, and phenotypic characteristics among strains. Geobacillus species exhibit broad catabolic versatility, facilitating their widespread occurrence across diverse environments. These bacteria are recognized for secreting commercially important enzymes, including lipases, xylanases, and amylases, widely utilized in various industrial and biotechnological processes.
According to the List of Prokaryotic Names with Standing in Nomenclature (LPSN, 2025), the genus Geobacillus currently includes 30 valid species, including Geobacillus thermodenitrificans, Geobacillus uzenensis, Geobacillus thermocatenulatus, Geobacillus kaustophillus, Geobacillus thermoglucosidasius, Geobacillus thermoleovorans, Geobacillus subterraneus, Geobacillus stearothermophilus, Geobacillus toebii, Geobacillus jurassicus, Geobacillus thermoglucosidans, Geobacillus stearothermophilus, and Geobacillus thermodenitrificans. Geobacillus stearothermophilus serves as the type species, with DSM 22 designated as its type strain [1,4].
Analysis based on the 16S rRNA gene revealed a cohesive grouping of 56 Geobacillus strains, exhibiting intra-species similarities between 99.3% and 100%, and inter-species similarities ranging from 96.0% to 99.4%, facilitating reliable distinctions [5]. However, limitations in taxonomic resolution using the 16S rRNA gene led researchers to adopt additional genetic markers, such as recN, infB, rpoB, and spo0A, providing enhanced phylogenetic clarity [6,7].
Distinctions between Geobacillus species in previous studies have been based on small chemotaxonomic differences and phenotypic characteristics [5]. Growth temperatures range from 35 °C to 80 °C, with most isolated strains growing between 45 °C and 70 °C [3]. The core genome of Geobacillus comprises approximately 1665 genes, 85% of which are shared with the genome of Bacillus subtilis 168, suggesting a common survival strategy centered around rapid growth and sporulation under nutrient-limited conditions [8].
Traditionally, bacterial classification has relied on methods such as 16S rRNA sequencing, DNA-DNA hybridization (DDH), and phenotypic characterization. Nonetheless, high sequence similarity within the 16S rRNA gene among Geobacillus species complicates precise identification and has resulted in misidentifications [9]. The growing number of studies classifying new species using only single-pathway markers has led to the rise of unreliable and confusing classifications within the genus [3,10,11,12]. To overcome these challenges, integrated approaches combining genomic and phenotypic analyses, including whole-genome sequencing, have been proposed to achieve higher phylogenetic resolution.
The Calientes geothermal field in Tacna, Peru, represents an underexplored source of microbial biodiversity with significant biotechnological potential [13,14]. Characterized by elevated temperatures and unique mineral compositions, these geothermal springs provide optimal conditions for thermophilic bacteria like Geobacillus [15,16]. These microorganisms have attracted considerable interest due to their ability to produce thermostable enzymes applicable in biotechnology, particularly in industrial processes involving the bioconversion and biodegradation of complex organic polymers such as lignocellulosic biomass (e.g., cellulose, hemicellulose, and lignin) [17].
Geobacillus species are highly valuable biotechnological resources due to their capability of producing stable and active enzymes under extreme environmental conditions, such as high temperatures and variable pH levels. These enzymes have found extensive industrial applications, notably in biofuel production and the bioconversion of industrial waste [18]. Moreover, certain Geobacillus strains efficiently degrade complex organic polymers like lignin and cellulose, offering promising opportunities for bioethanol production from renewable biomass [19]. Ongoing genomic and metabolic engineering research efforts further enhance the biotechnological potential of these bacteria, optimizing their utilization in industrial applications [20].
Geobacillus species have attracted considerable attention not only due to their capacity to produce thermostable enzymes but also because of their potential to biosynthesize secondary metabolites with diverse industrial and pharmaceutical applications. These metabolites include compounds like fengycin, known for its antimicrobial and antifungal properties, valuable in bioremediation and biological control applications, and lanthipeptides or betalactones, which exhibit potential therapeutic properties, including antibiotic and anticancer activities, relevant to pharmaceutical development [21]. The identification and characterization of secondary metabolite biosynthetic gene clusters (BGCs) in thermophilic bacteria can reveal novel compounds beneficial for biotechnological and biomedical use [17,18].
The study of genomic diversity and characterization of new Geobacillus strains is essential from both a taxonomic and biotechnological perspective. Genomic analysis of new strains allows for the identification of genes involved in specific metabolic processes, such as the degradation of starch and hemicellulose, which are crucial for biomass conversion into biofuels and other value-added products [22,23].
In this study, we aimed to characterize Geobacillus sp. G4, a thermophilic strain isolated from a geothermal environment in southern Peru, through phenotypic and enzymatic analyses. Furthermore, we sought to sequence, assemble, and annotate its genome; determine its taxonomic position; and identify genes and metabolic pathways with potential biotechnological relevance, particularly those involved in the production of thermostable enzymes and the degradation of complex carbohydrates for industrial applications.
2. Materials and Methods
2.1. Bacterium Isolation and Characterization of Geobacillus sp. G4
Sediment samples were obtained from a geothermal spring in the Calientes geothermal field, Candarave, Tacna, Peru. The spring, designated as G8, was selected based on the presence of sediment, elevated temperatures (>60 °C), and accessibility. The G8 spring, with a water temperature of 65.3 °C and a pH of 7.35, is located at an altitude of 4162 m above sea level (UTM coordinates: 0.379908 E, 81.09185 N). The area surrounding the spring was characterized by sparse vegetation and the accumulation of brownish sediments (Figure 1). Approximately 250 g of sediment was collected from the upper 3 cm layer of the sediment bed, at a depth of up to 15 cm from the water surface, at the edge of the geothermal spring using a sterile plastic bottle. A suspension of sediment and spring water was transferred into a sterile thermos bottle to maintain a temperature close to that of the sample during both sampling and transport to preserve microbial viability.

Figure 1.
Map of the geothermal sampling area in Calientes, Candarave, using ArcGIS version 10.5. Image of the geothermal spring sampled in this study, located in the Calientes geothermal field, Candarave, Tacna, Peru.
A sediment sample (10 g) was initially enriched in Luria–Bertani (LB) liquid medium (consisting of 10 g/L peptone, 5 g/L yeast extract, and 10 g/L sodium chloride) at a 10% (v/v) concentration. The enrichment culture was incubated at 60 °C for 48 h. After incubation, 100 μL of the enriched culture (OD 600 nm = 0.2) was spread onto LB agar plates supplemented with 1% (w/v) starch as the sole carbon source to selectively isolate amylolytic bacteria. The plates were incubated at 60 °C until bacterial colonies appeared. After two days, bacterial colonies were individually collected and streaked three times on the same LB agar to ensure purity. Five amylolytic colonies (G1, G2, G4, G16, and G23) were isolated, and the strain G4, which exhibited the highest amylase activity levels, was chosen for further characterization and genomic analysis. Amylase activity was quantified using the Miller method [24]: 100 μL of starch solution (0.5% w/v) and 100 μL of G4 culture supernatant were incubated at 60 °C for 15 min. Then, 300 μL of DNS reagent was added, and absorbance was measured at 540 nm.
The morphology of the G4 strain was evaluated by analyzing colony shape, pigmentation, and microscopic characteristics using Gram staining, endospore formation assays, and motility tests. The effect of temperature on the growth of strain G4 was assessed by culturing it in LB liquid medium (pH 7) at a range of temperatures (37, 45, 50, 55, 60, 65, 70, 75, and 80 °C) for 48 h. Similarly, the effect of pH on growth was evaluated by incubating G4 in LB medium buffered at pH values ranging from 4 to 8 (100 mM sodium citrate for pH 4 and 5, 100 mM sodium phosphate for pH 6 and 7, and 100 mM Tris-HCl for pH 8) at a fixed temperature of 60 °C for 48 h. For acid production assays, substrates such as lactose, cellobiose, glycerol, sucrose, and xylose were used, following the methodology described by Mac Faddin (2000) [25].
Carbon utilization of strain G4 was determined by culturing it in LB liquid medium (pH 7) supplemented with various substrates, including carboxymethylcellulose (CMC), starch, pectin, microcrystalline cellulose, casein, lactose, xylose, glucose, sucrose, cellobiose, and maltose, each at 1% (w/v) as a carbon source (Sigma Aldrich, St. Louis, MO, USA). Cultures were incubated at 60 °C for 48 h. Bacterial growth was monitored by measuring optical density at 600 nm.
The extracellular hydrolytic enzyme activities, including cellulase, pectinase, xylanase, amylase, protease, and lipase, were assessed. The G4 isolate was cultivated on LB agar plates supplemented individually with specific substrates: carboxymethylcellulose (CMC) 1% (w/v), pectin 0.5% (w/v), oat spelt xylan 1% (w/v), potato starch 1% (w/v), casein 1% (w/v), and tributyrin 0.5% (v/v), following the protocol described by Guta et al. (2024) with minor modifications [26]. Plates were incubated at 60 °C for 24–48 h. The presence of clear zones surrounding colonies indicated positive enzymatic activity.
To analyze enzyme production by isolate G4, time-course experiments were performed. G4 was inoculated into 250 mL Erlenmeyer flasks containing 50 mL of LB liquid medium at pH 7.0, individually supplemented with the enzyme-specific substrates mentioned above. Cultures were incubated at 60 °C, and 2 mL samples were withdrawn at intervals of 3, 6, 12, 24, 30, and 48 h. Samples were centrifuged at 10,000× g for 10 min at 4 °C, and the resulting supernatants were used for enzyme assays.
The enzymatic activities of cellulase, pectinase, xylanase, and amylase were determined using their respective substrates buffered with 100 mM sodium phosphate (pH 7.0), quantifying the reducing sugars released according to the Ticona method [27]. One unit of enzymatic activity was defined as the amount of enzyme required to release 1 µmol of reducing sugar per minute.
Protease activity was assessed by mixing 25 µL of enzyme solution with 130 µL of 0.5% (w/v) casein buffered in 100 mM Tris-acetate (pH 7.5), following the methodology described by Oluoch et al. (2018) [28]. One unit of protease activity was defined as the amount of enzyme that released 1 µmol of tyrosine per minute. Lipase activity was measured titrimetrically using an olive oil emulsion as the substrate, as described by Sharma et al. (2017) [29]. One unit of lipase activity was defined as the enzyme amount releasing 1 μmol of fatty acid per minute under assay conditions.
2.2. DNA Extraction and Sequencing
Genomic DNA was extracted from bacterial strain G4 for whole-genome sequencing. Strain G4 was first cultured in 10 mL of Luria–Bertani (LB) broth and incubated at 60 °C for 48 h. The culture was centrifuged at 8000× g for 15 min at 4 °C, and the supernatant was discarded. The resulting cell pellet was resuspended in 5 mL of Tris-HCl buffer (pH 8.0). Genomic DNA was isolated using the Bacterial Genomic DNA Isolation Kit (Norgen Biotek Corp., Thorold, ON, Canada) following the manufacturer’s protocol. The concentration and purity of the DNA were measured using a BioTek Epoch 2T spectrophotometer (Agilent Technologies, Inc., Santa Clara, CA, USA).
DNA libraries were prepared using the Illumina TruSeq Nano DNA Library Prep Kit (350), and sequencing was performed using the Illumina NovaSeq platform with paired-end 150 bp reads at Macrogen Inc. (Seoul, Republic of Korea).
2.3. Quality Analysis, Genome Assembly, and Annotation of Geobacillus sp. G4
The raw fastq files (paired-end) comprising 13,778,078 sequences with an average length of 151 bp were imported into the FastQC v0.12.1 tool for quality assessment. The quality filtering was performed using Trimmomatic v0.38 to remove sequences with low-quality scores, applying the following parameters: SLIDINGWINDOW (4:30) and PHRED 30. The remaining sequences were then imported into the SPAdes v3.15.4 tool for genome assembly using the parameters k-mer (21, 33, 55, and 77) and a Phred score of 64 (Illumina). Short contigs (<200 bp) were removed from the assembly using seqtk v.1.4 (https://github.com/lh3/seqtk, accessed on 13 November 2024). The completeness and contamination of the genome were analyzed using the CheckM v1.2.2 tool and CoverM v0.7.0 (https://github.com/wwood/CoverM, accessed on 13 November 2024), while the size and quality of the assembled genome were assessed using Quast v5.2.0 and Busco v5.5.0 [30,31,32].
Functional genome annotation was performed in the Galaxy (https://usegalaxy.org/) environment using Prokka v1.14.6 with default parameters. Prokka predicted CDSs, rRNAs, and tRNAs, assigning functions initially through BLAST+ searches against core databases (ISfinder, NCBI AMR, and UniProtKB/SwissProt), followed by refinement using HMMER3 [33,34,35,36,37]. After annotation, the genome was mapped according to its functional categories and enzyme classes using the online mapper eggNOG-mapper v2 (eggNOG-mapper) with the eggNOG 5.0 database [38]. Type strains from the most similar species, based on the results from the Digital DNA-DNA hybridization (dDDH) analysis, were used for comparison with the literature.
To analyze and highlight the biotechnological potential of the Geobacillus sp. G4 strain, three metabolic pathways were highlighted in the KEGG annotation: glycogen biosynthesis (M00854), glycogen degradation (M00855), and trehalose biosynthesis (M00565) (Supplementary Figure S1). The visualization of the pathways of interest was performed using the KEGG mapper (www.genome.jp/kegg/mapper, accessed on 20 November 2024) via the KEGG database.
To identify gene clusters related to secondary metabolism (BCGs) in microorganisms, AntiSMASH v7.1.0 software was used [39]. The Antibiotic Resistance Genes Database (ARDB) was used to select and annotate genes potentially responsible for antibiotic resistance [40].
The genome of Geobacillus sp. G4, along with three reference genomes (G. kaustophilus HTA426, G. kaustophilus NBRC12445, and G. thermoleovorans KCTC3570), was functionally annotated using the eggNOG-mapper online tool (http://eggnog-mapper.embl.de/, accessed on 6 January 2025). Genes related to four functional categories of interest—carbon starvation, DNA repair and supercoiling, heat-shock response, and oxidative stress—were selected based on their presence in the genomes. Gene presence/absence analyses and graphical visualization were performed using R statistical software (https://www.r-project.org/).
2.4. Taxonomic Classification of Geobacillus sp. G4
2.4.1. Digital DNA Hybridization (dDDH) and Average Nucleotide Identity (ANI)
Digital DNA hybridization (dDDH) was performed on the TYGS platform—DSMZ (tygs.dsmz.de) for the taxonomic classification of the strain and identity confirmation [41]. For phylogenomic inference to compare complete genomes in dDDH, the GBDP method (Genome BLAST Distance Phylogeny) was used, employing the trimming algorithm, formula d5, and bootstrap 100 [42]. To calculate genomic similarity using DDQ (Digital DNA-DNA Hybridization), GGDC v4.0 software integrated into the TYGS platform was used. The average nucleotide identity (ANI) analysis was calculated using the JSpeciesWS platform (https://jspecies.ribohost.com/), which provides ANIb, representing the average nucleotide identity based on BLAST calculations [43], and ANIm, representing the average nucleotide identity based on MUMmer calculations [43].
2.4.2. 16S rRNA Gene-Based Phylogenetic Tree Reconstruction
16S rRNA gene sequences from the genomes were extracted using barnnap v0.9 (https://github.com/tseemann/barrnap, accessed on 11 January 2025). All information regarding the sequences used can be found in Supplementary Table S1. Sequence alignment was performed using MAFFT v7.526 in default mode [44]. After alignment, all sequences were trimmed using TrimAl v1.4.1 [45]. The selection of the most appropriate model for tree reconstruction was performed using ModelTest-NG v0.1.7 [46], suggesting the TIM1+I model. The phylogenetic tree was constructed using the Maximum Likelihood method [47], utilizing RAxML-NG X v1.2.2 [48] with the following parameters: 10 random initial parsimony trees, nucleotide substitution matrix model (TIM1+I), and 1000 bootstrap replications. The Bacillus subtilis DSM10 strain was used as an outgroup.
2.4.3. Phylogenetic Tree Reconstruction
The genomes used were obtained from complete genomes deposited in GenBank. The selection of type strains for valid Geobacillus and Bacillus species used was based on the LPSN (List of Prokaryotic names with Standing in Nomenclature—lpsn.dsmz.de) [41,49]. All information regarding the complete genomes used can be found in Supplementary Table S2.
Two phylogenetic trees were constructed using the studied genome. The first tree was reconstructed using molecular markers extracted from the genomes using AMPHORA2 [50], and data on the extracted markers are available in Supplementary Table S3. Sequence alignment was performed using the MAFFT v7.526 aligner in default mode [44]. After alignment, all sequences were trimmed using TrimAl v1.4.1 [45]. The selection of the most appropriate model for tree reconstruction was performed using ModelTest-NG v0.1 [46], with the LG+I+G4+F model suggested. The phylogenetic tree was constructed using the Maximum Likelihood method [47], utilizing RAxML-NG X v1.2.2 [48] with the following parameters: 10 random initial parsimony trees, nucleotide substitution matrix model (LG+I+G4+F), and 1000 bootstrap replications. The Bacillus subtilis DSM10 strain was used as an outgroup.
The second tree was reconstructed using the FastME v2.1.6.1 tool [51] with the GBDP method (Genome BLAST Distance Phylogeny) integrated into the TYGS platform (tygs.dsmz.de). The clade lengths are scaled in terms of GBDP distance formula d5, and 100 pseudo-bootstrap replicates were performed. Only pseudo-bootstrap support values above 60% were accepted. The tree was rooted at the midpoint [52]. The Bacillus subtilis DSM10 strain was used as an outgroup.
For genome comparison, the online platform Proksee (proksee.ca) was used [53]. Comparisons were made using the genome of the type strains G. thermoleovorans KCTC3570 and G. kaustophilus NBRC102445 in comparison with the Geobacillus sp. G4 strain. BLAST v1.3.1 and BLAST Formatter v1.0.3 tools were used to check identity and similarity profiles between the genomes [42].
3. Results
3.1. Characterization of Geobacillus sp. G4
The Geobacillus sp. G4 strain is a Gram-positive, rod-shaped bacterium forming round, white-yellow colonies with an average diameter of 1.4 mm (Supplementary Table S4). Characterization revealed that Geobacillus sp. G4 is aerobic, non-motile, and forms endospores. Optimal growth conditions (50 °C to 70 °C and pH 6.0 to 7.5) were determined in LB liquid medium based on maximal bacterial growth, as measured by optical density at 600 nm. Additionally, acid production from lactose, cellobiose, glycerol, sucrose, and xylose was observed (Table 1).

Table 1.
Phenotypic characteristics for differentiation among G. kaustophilus, G. thermoleovorans, and Geobacillus sp. G4.
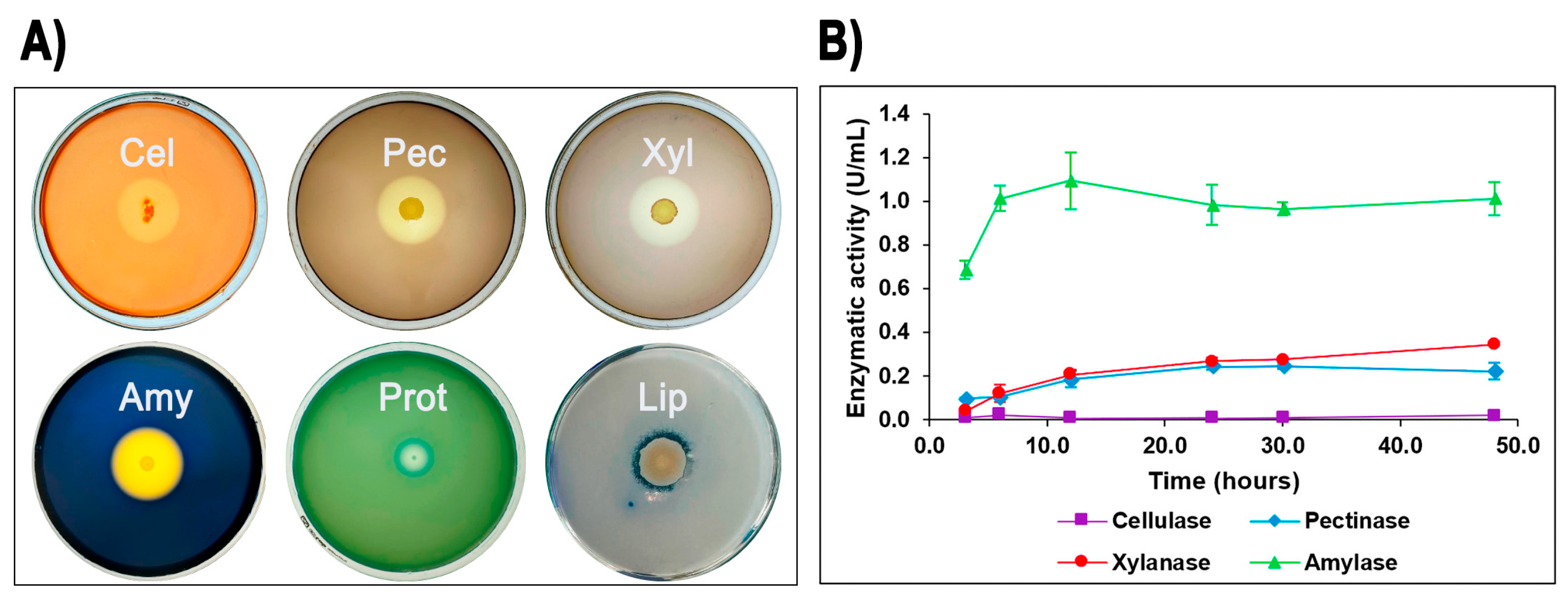
The ability of isolate G4 to produce extracellular hydrolytic enzymes was evaluated on agar plates. After incubation at 60 °C for 24–48 h, isolate G4 demonstrated positive enzymatic activities for cellulase, pectinase, xylanase, amylase, protease, and lipase, indicated by distinct clear zones around colonies. Among these, Geobacillus sp. G4 showed the largest halo formation for amylase (starch), followed by pectinase, xylanase, and cellulase (Figure 2A).
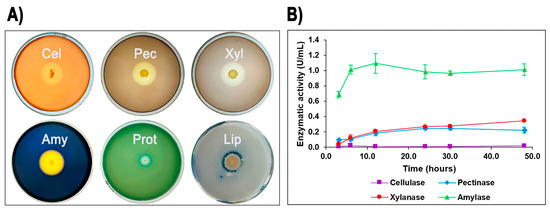
Figure 2.
Quantitative and qualitative enzymatic activity of Geobacillus sp. G4. Extracellular enzymatic activities of Geobacillus sp. G4. (A) Formation of clear halos on agar plates indicating positive extracellular enzyme production: Amy (amylase), Pec (pectinase), Xyl (xylanase), Cel (cellulase), Prot (protease), and Lip (lipase). (B) Enzyme production profiles measured in LB liquid media over time.
To further quantify these enzymatic activities, isolate G4 was cultured in liquid media containing potato starch, oat spelt xylan, pectin, carboxymethylcellulose (CMC), olive oil, and casein as carbon sources. Enzyme assays revealed measurable production of amylase (maximum activity: 1.09 U/mL at 12 h), xylanase (0.34 U/mL at 48 h), and pectinase (0.24 U/mL at 30 h), as well as low but detectable cellulase activity (0.02 U/mL at 48 h) (Figure 2B). In contrast, protease and lipase activities, although clearly evident in agar plate assays, were below detection limits in liquid media under the tested conditions.
3.2. Genome Description and Taxonomic Classification of Geobacillus sp. G4
The results regarding the preliminary genome draft of Geobacillus sp. G4 is presented in Table 2. The strain genome has a size of approximately 3.4 Mb with a coverage of ~1225x. The assembly comprised 164 contigs, with a total G+C content of 52.59%. The genome completeness and contamination were 99.45% and 0.95%, respectively. The N50 and L50 values for the assembly were 236,119 bp and 5 contigs, respectively. The annotation identified a total of 3490 genes: 2047 coding sequences (CDS), 5 rRNAs, 88 tRNAs, 1 transfer-messenger RNA, and 1349 putative proteins. Regarding the gene clusters related to secondary metabolism (BCGs), seven clusters were identified: betalactone, T3PKS, Lanthipeptide class-i, NI-siderophore, terpene, fengycin, and RiPP-like.
Table 2.
Genome assembly data for Geobacillus sp. G4 was performed using SPAdes v.3.15.4. The annotation was conducted using Prokka v.1.14.6, and quality parameters were assessed using Busco v.5.5.0 and Quast v.5.2.0. All processes were carried out on the Galaxy v24.1.2 server.
The nucleotide average identity (ANI) values between the Geobacillus sp. G4 genome and other species of the Geobacillus genus show similarity with Geobacillus kaustophilus and Geobacillus thermoleovorans, with ANI values above 97% (Table 3). Thus, the obtained results suggest that Geobacillus thermoleovorans KCTC3570 and Geobacillus sp. G4 are sufficiently similar to be considered as belonging to the same species.
Table 3.
Average nucleotide identity (ANI) calculated for Geobacillus sp. G4 using type strains of each species of the Geobacillus genus, calculated on the JSpeciesWS platform (ANIb and ANIm). Information is available at the following link: (https://jspecies.ribohost.com/jspeciesws/, accessed on 11 January 2025).
In the dDDH analysis, the Geobacillus sp. G4 strain showed dDDH values above 70% with Geobacillus kaustophilus HTA426 (79.1%) and Geobacillus kaustophilus NBRC102445 (78.5%) (Table 4). However, the Geobacillus thermoleovorans KCTC 3570 strain exhibited the highest similarity with Geobacillus sp. G4, with a dDDH value of 83.2% and a GC content difference of only 0.31%, shows a high genomic proximity. This suggests that both organisms are closely related, meaning Geobacillus sp. G4 could be a variant of G. thermoleovorans. Other species showed dDDH values below 70% and a larger GC content difference. These data indicate that, among the species compared, Geobacillus thermoleovorans is the closest to Geobacillus sp. G4 in terms of genomic similarity.
Table 4.
Digital DNA hybridization (dDDH) generated by the TYGS-DSMZ platform between the Geobacillus sp. G4 genome and the species described for the genus up to August 2024. All type strains of each species according to the List of Prokaryotic Names with Standing in Nomenclature (https://lpsn.dsmz.de/, accessed on 9 January 2025).
The 16S rRNA gene sequence of the Geobacillus sp. G4 strain showed 99.87% identity with Geobacillus kaustophilus strain BGSC 90A1 and 99.80% identity with Geobacillus thermoleovorans strain LEH-1 (Supplementary Table S5). The Geobacillus thermoleovorans BGSC 96A1 lineage also had 100% query coverage and 99.61% identity. Other Geobacillus and Bacillus strains showed identity percentages ranging from 99.08% to 99.87%, with query coverages ranging from 92% to 100%.
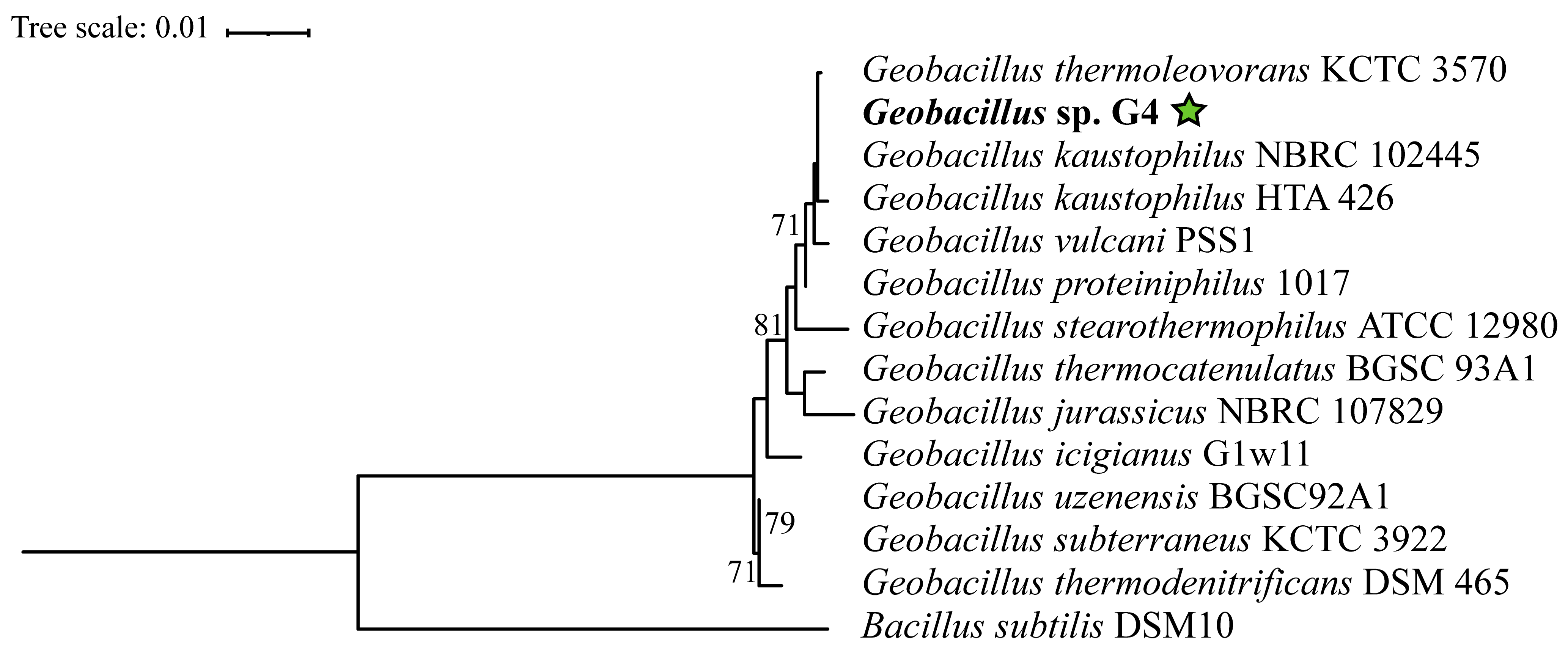
In the phylogenetic tree constructed based on the 16S rRNA gene, the Geobacillus strains are distributed across several clades without statistical support (Figure 3), indicating significant diversification within this genus. However, Geobacillus sp. G4 is positioned within a clade supported by a bootstrap value of 71%, containing the strains Geobacillus thermoleovorans KCTC3570, Geobacillus kaustophilus HTA426, Geobacillus kaustophilus NBRC102445, Geobacillus proteiniphilus 1017, and Geobacillus vulcani PSS1.
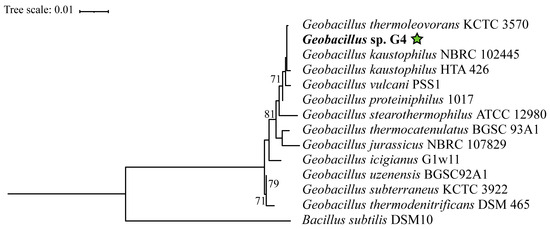
Figure 3.
Phylogenetic tree based on the 16S rRNA gene of 13 type strains of Geobacillus and Bacillus, extracted from the complete genome using the (barrnap) tool. Accession numbers and references are available in Supplementary Table S1. Evolutionary analyses were conducted using the RAxML tool. The strain Geobacillus sp. G4, in bold, corresponds to the 16S gene from the genome assembled in this study. The green star indicates the position of the strain isolated in this study.
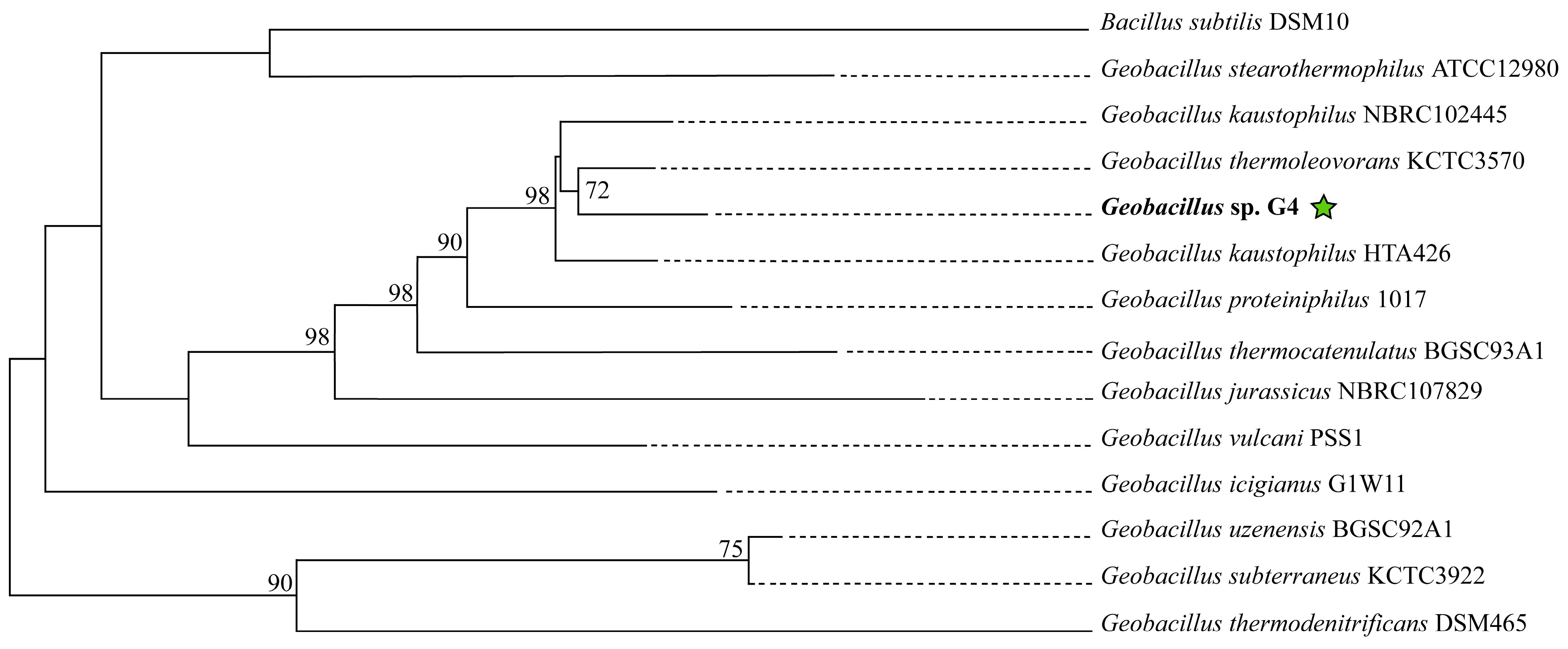
In the phylogenomic tree reconstructed using molecular markers extracted with AMPHORA2, the strain Geobacillus sp. G4 is grouped with Geobacillus kaustophilus HTA426 and Geobacillus thermoleovorans KCTC3570, forming a clade supported by a bootstrap value of 95% (Supplementary Figure S1). In the phylogenomic tree reconstructed on the TYGS platform (Figure 4), the strain Geobacillus sp. G4 is in the clade with the strain Geobacillus thermoleovorans KCTC3570, supported by a bootstrap of 72%.
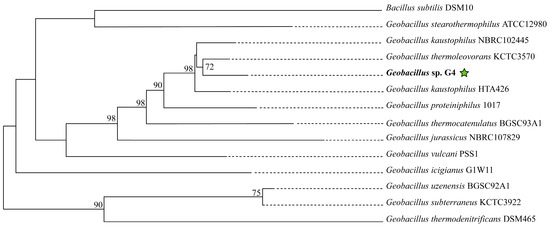
Figure 4.
Phylogenetic tree of 13 type strains of Geobacillus and Bacillus, constructed on the online platform TYGS available at (https://tygs.dsmz.de). The green star indicates the position of the strain isolated in this study.
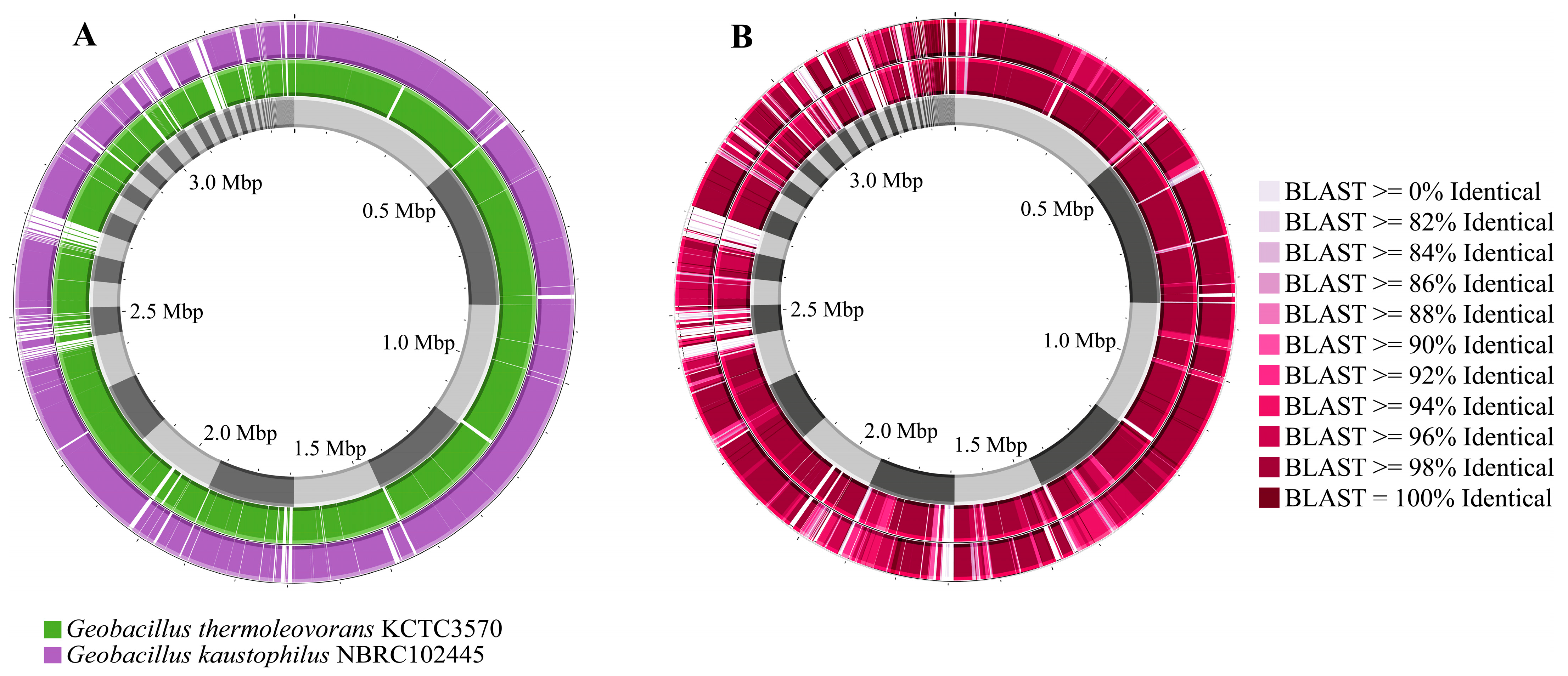
Comparing the genomes of Geobacillus sp. G4 with the genomes of G. kaustophilus NBRC102445 and G. thermoleovorans KCTC3570 reveals several gaps between them, with a higher frequency observed against G. kaustophilus NBRC102445 (Figure 5A). It was also noted that Geobacillus sp. G4 showed greater similarity across several regions of its genome compared to G. thermoleovorans KCTC3570 (Figure 5B).
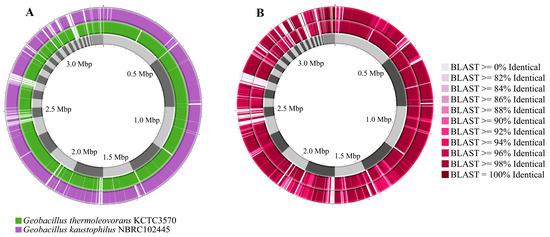
Figure 5.
Comparison of the genomes of G. thermoleovorans KCTC3570 and G. kaustophilus NBRC102445 with the genome of the isolated strain Geobacillus sp. G4. (A) Geobacillus sp. G4 genome (gray), G. thermoleovorans KCTC3570 genome (green), and G. kaustophilus NBRC102445 genome (purple). (B) Similarity comparison between the genomes of Geobacillus sp. G4 (gray), G. thermoleovorans KCTC3570 (inner circle), and G. kaustophilus NBRC102445 (outer circle).
3.3. Functional Analysis of Geobacillus thermoleovorans G4 Strain
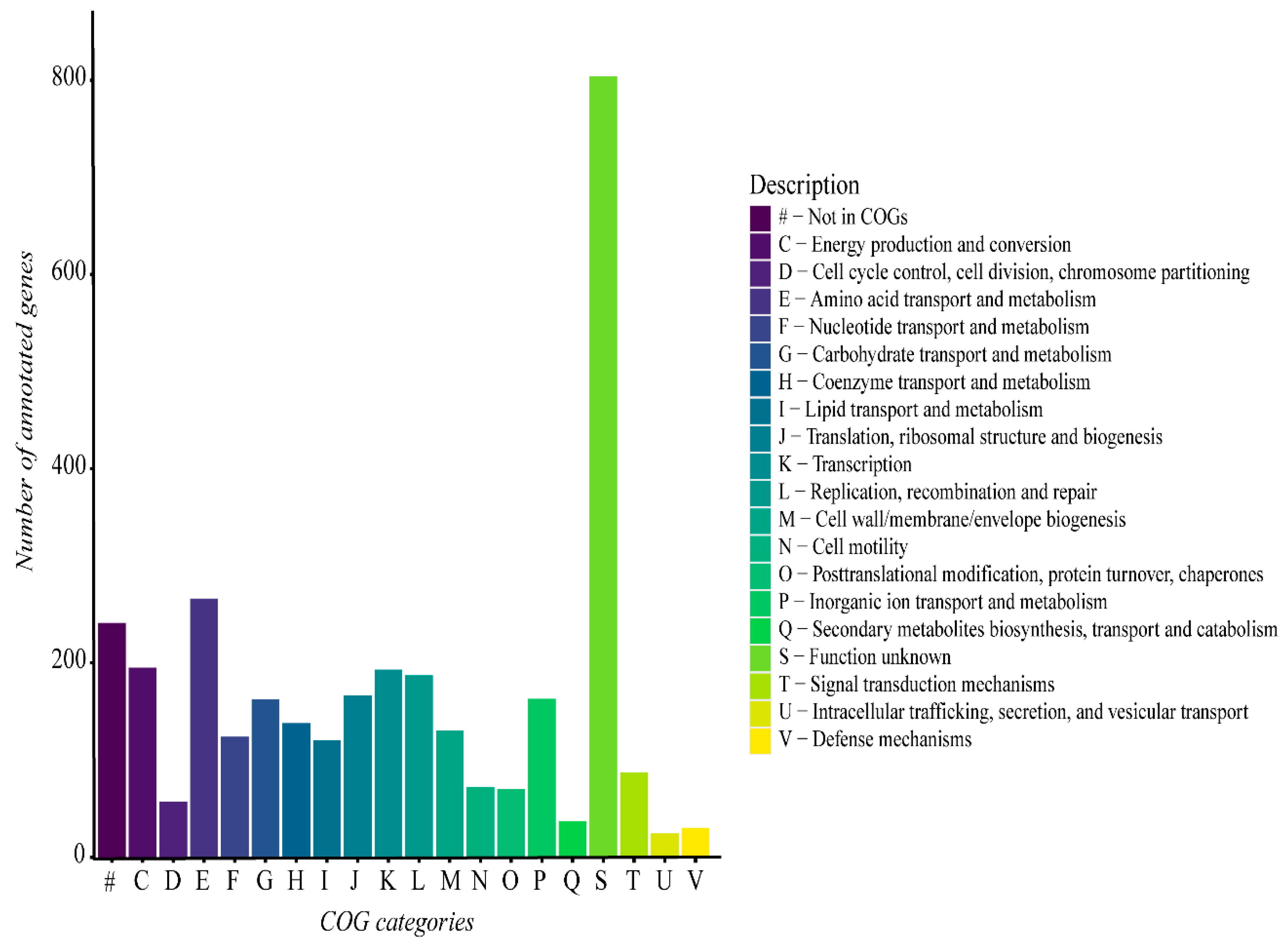
The functional analysis of the G. thermoleovorans G4 genome revealed a predominance of genes involved in different various metabolic categories, including metabolism, transport, transcription, replication, and repair, among others (Figure 6). Additionally, a large portion of genes were not classified into any known COG category, highlighting the presence of genes with functions that remain unclear within the context of the G. thermoleovorans G4 genome.
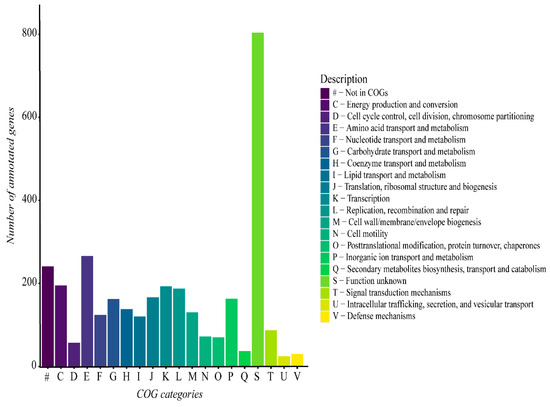
Figure 6.
Functional gene categories derived from the functional annotation of the G. thermoleovorans G4 genome using Prokka software and the UniProt database on the Galaxy server. Categories were classified using the eggNOG-mapper online tool with the eggNOG5 database.
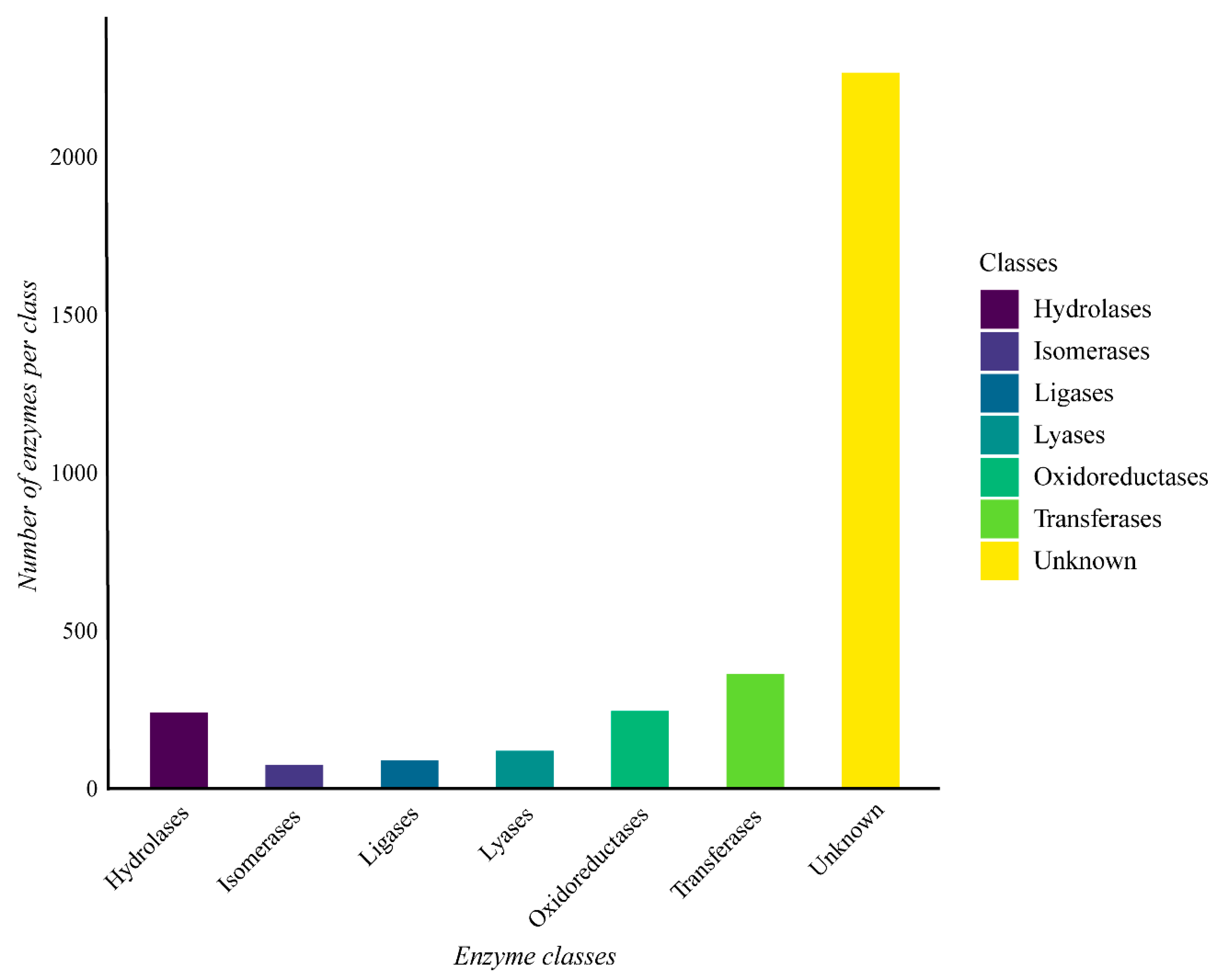
The genome of G. thermoleovorans G4 shows a predominance of enzymes belonging to the various classes, with hydrolases, transferases, and oxidoreductases being the most abundant (Figure 7). Additionally, many sequences lacked known annotations for any of the classes.
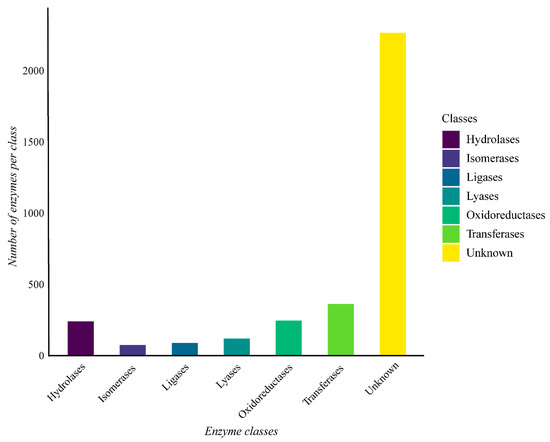
Figure 7.
Enzyme classes were obtained through functional annotation of the assembled genome of Geobacillus thermoleovorans G4 using Prokka software on the Galaxy server (https://usegalaxy.org/) and classified using the eggNOG-mapper online tool with the eggNOG 5 database (http://eggnog-mapper.embl.de/). The “Unknown” category refers to proteins encoded in the genome that have been annotated as possible enzymes, but do not have an assigned EC number.
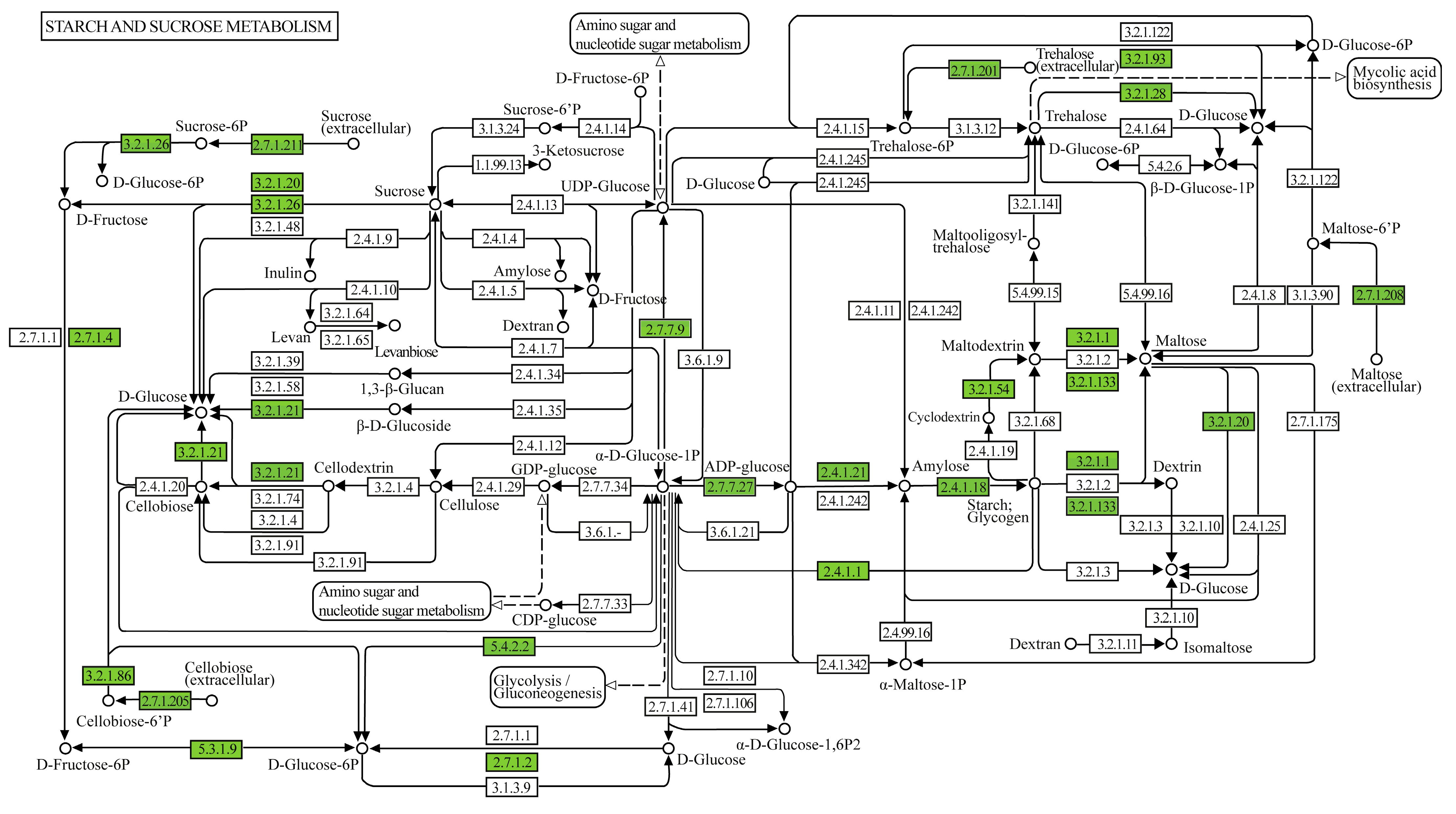
In the set of metabolic pathways involved in the degradation of starch and sucrose present in the genome of Geobacillus thermoleovorans G4, enzymes involved in the degradation, conversion, and biosynthesis of sugars such as glucose, fructose, and their phosphorylated derivatives were observed (Figure 8). In the glycogen biosynthesis pathway (Supplementary Figure S2A), the enzymes annotated were 1,4-alpha-glucan branching enzyme (GBE1, glgB), glucose-1-phosphate adenylyltransferase (glgC), UTP-glucose-1-phosphate uridylyltransferase (UGP2, galU, galF), and starch synthase (glgA), all classified under transferases, related to starch synthesis. In the glycogen degradation pathway (Supplementary Figure S2B), the annotated enzymes were glycogen phosphorylase (PYG, glgP), pullulanase (pulA), phosphoglucomutase (pgm), and phosphomannomutase/phosphoglucomutase (pmm-pgm), classified under transferases, isomerases, and hydrolases, related to the mobilization and breakdown of glycogen and starch. In the trehalose biosynthesis pathway (Supplementary Figure S2C), the annotated enzymes were glucose-1-phosphate adenylyltransferase (glgC), starch synthase (glgC), and 1,4-alpha-glucan branching enzyme (GBE1, glgB), classified under transferases, related to starch synthesis.
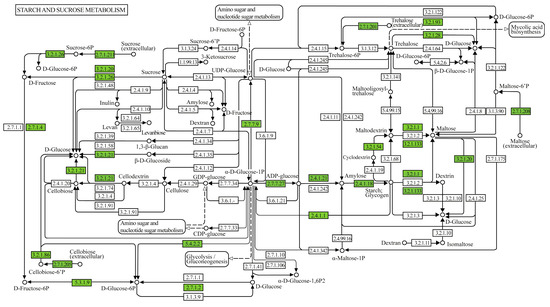
Figure 8.
Metabolic pathway map of starch and sucrose metabolism displaying genes and enzymes of interest annotated in the genome of G. thermoleovorans G4. Annotations were performed using the KEGG Mapper tool (www.genome.jp/kegg/mapper, accessed on 20 November 2024), identifying key enzymes involved in the metabolism of sugars and complex carbohydrates.
The analysis of antibiotic resistance genes in the genome of G. thermoleovorans G4 identified two clusters related to glycopeptide resistance: vanY/vanB_cl and vanT/vanG_cl (Supplementary Table S6). The similarity of these genes between Geobacillus sp. G4 and other species ranged from 34.15% to 36.41% for vanY/vanB_cl and from 35.9% to 34.15% for vanT/vanG_cl, with both species compared showing high similarity.
For the G. thermoleovorans G4 strain, clusters for seven different types of secondary metabolites were identified: betalactone, T3PKS, Lanthipeptide class-i, NI-siderophore, terpene, fengycin, and RiPP-like (Supplementary Table S7). Comparatively, the strains Geobacillus kaustophilus HTA425 (G.K HTA425), Geobacillus kaustophilus NBRC102445 (G.K NBRC102445), and Geobacillus thermoleovorans KCTC3570 (G.T KCTC3570) also presented clusters for some of the analyzed secondary metabolites, except for NI-siderophore, fengycin, and RiPP-like, where no clusters were found. These observations suggest a shared biosynthetic capability among the strains, with minimal variations in the types of secondary metabolites detected.
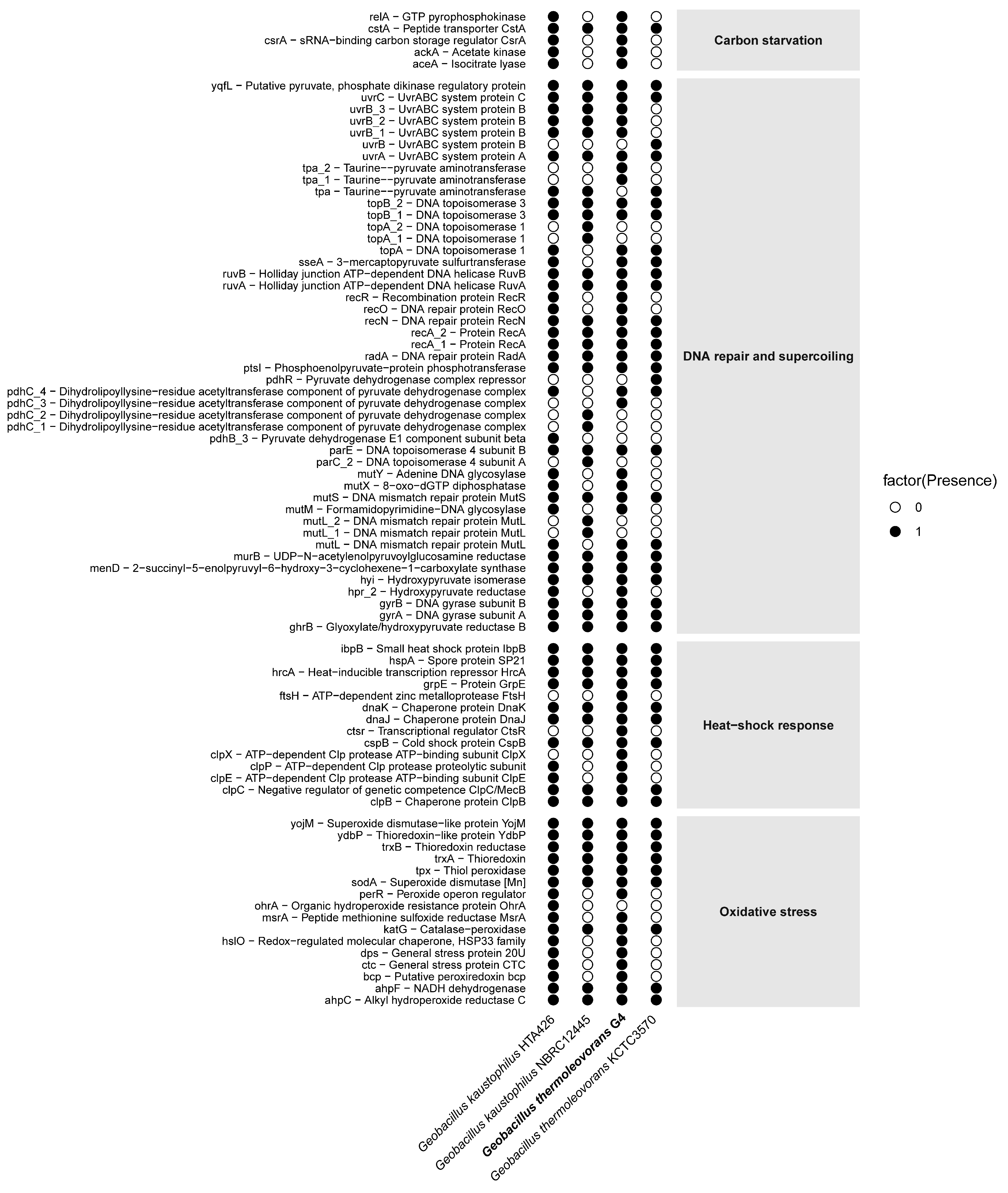
The Geobacillus thermoleovorans G4 strain presents a complete repertoire of genes (Figure 9) associated with carbon starvation, including relA, cstA, ackA, and accA. Complementarily, in the DNA repair and supercoiling category, a diverse set of genes was observed, such as uvrABC, recA, recO, recR, mutS, mutL, and gyrAB. In addition, classical heat-shock response genes were detected, including hrcA, dnaK, groEL, clpB, and ibpB. Finally, the presence of oxidative stress-related genes, such as sodA, katA, ahpC, trxB, and msrA, was also confirmed. The complete list of selected genes, along with their corresponding COG reference numbers, can be found in Supplementary Table S8.
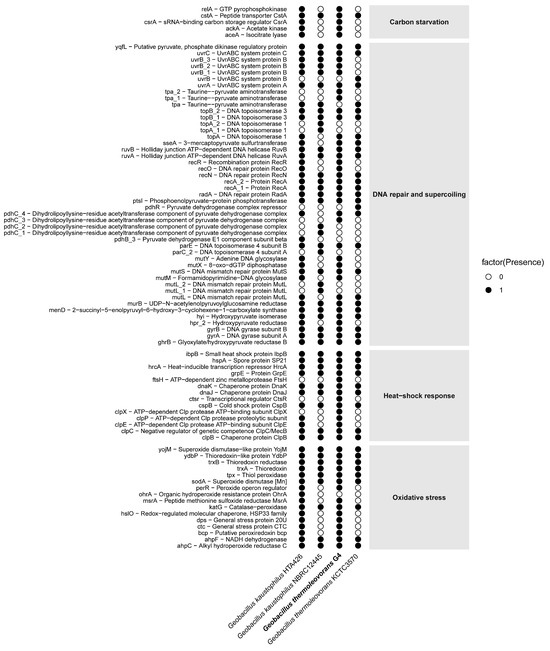
Figure 9.
Distribution of presence/absence of stress-related genes in the genome of G. thermoleovorans G4 and in reference genomes of other Geobacillus strains. Each row represents an annotated gene, while each column corresponds to a strain. Black circles indicate the presence of the gene, and white circles indicate its absence. Genes are grouped into four main functional categories: carbon starvation, DNA repair and supercoiling, heat-shock response, and oxidative stress.
4. Discussion
The genomic characterization and functional analysis of Geobacillus sp. G4, isolated from the geothermal field of Calientes in Tacna, Peru, provided valuable insights into its biotechnological potential. Geobacillus sp. G4 exhibited morphological and physiological characteristics typical of the Geobacillus genus. Initially, a direct comparison with Geobacillus kaustophilus and Geobacillus thermoleovorans was established, given that preliminary phylogenetic analyses and previous studies identified these species as the closest phylogenetic relatives to strain G4, sharing similar physiological profiles and temperature ranges [1,46].
Morphologically, strain G4 is characterized as a Gram-positive, endospore-forming rod, producing yellowish-white colonies approximately 1.4 mm in diameter, features that are commonly observed among thermophilic Geobacillus strains [3,5]. Its growth within a temperature range of 50–70 °C and an optimal pH of 6.0–7.5 further confirms its adaptation to thermophilic environments.
Table 1 specifically compares the properties of G4 with G. kaustophilus and G. thermoleovorans, highlighting relevant similarities and differences. All three strains are aerobic, Gram-positive, non-motile, and endospore-forming; however, notable differences were observed regarding acid production from various substrates. For instance, G. kaustophilus does not produce acid from lactose, whereas G4 does, along with cellobiose, glycerol, sucrose, and xylose, indicating broader or more efficient metabolic pathways for sugar degradation. The shared ability of G4 and G. thermoleovorans to metabolize lactose and glycerol supports their close relationship [1].
Regarding the optimal pH range, Geobacillus sp. G4 exhibits growth between pH 6.0 and 7.5, slightly narrower compared to some G. thermoleovorans strains that can tolerate conditions up to approximately pH 9.0. Additionally, the tolerance of strain G4 to NaCl concentrations up to 2.5% (w/v) indicates adaptations to moderately saline environments, a trait shared by many thermophilic Geobacillus species inhabiting geothermal springs [19,54,55].
Geobacillus sp. G4 exhibited extracellular enzymatic activities for cellulase, pectinase, xylanase, amylase, protease, and lipase on agar plates, with notably prominent hydrolysis halos observed particularly on starch-containing media. Similar enzyme profiles were previously reported for other thermophilic strains, such as G. stearothermophilus, Geobacillus sp. GS33, Geobacillus sp. WSUCF1, and Geobacillus sp. DUSELR13, particularly regarding the stability of amylase and xylanase at elevated temperatures [56,57,58,59]. Although protease and lipase activities were detectable in solid media assays, they were not quantifiable in liquid cultures, potentially due to differences in gene regulation or specific culture conditions [60,61].
Moreover, the diverse enzymatic profile identified in Geobacillus sp. G4 highlights a promising opportunity for developing enzyme cocktails tailored for efficient lignocellulosic biomass conversion into bioethanol and other valuable bioproducts. Despite their recognized biotechnological potential, relatively few studies have focused on formulating enzyme cocktails based on multiple thermostable enzymes derived from Geobacillus species [2,18].
Genome sequencing revealed a 3.4 Mb genome with a G+C content of 52.59%, consistent with genomic attributes previously reported for the Geobacillus genus, including strains such as Geobacillus stearothermophilus EF60045 and SJEF4-2 and Geobacillus sp. E55-1, and Geobacillus sp. 56T2 and 56T3 [22,62,63]. Studies demonstrate that the pan-genome of these bacteria reveals great genetic diversity, facilitated by horizontal gene transfer (HGT), which enables adaptation to various ecological niches [64]. However, classification based solely on 16S rRNA sequences has proven to be limited due to the high similarity between species [9]. To address this issue, ANI and dDDH analyses, combined with complete genome sequences, provide more precise phylogenetic resolution [65].
In our study, ANI and dDDH analyses revealed that Geobacillus sp. G4 is closely related to G. thermoleovorans KCTC3570, with ANI values above 97% and dDDH values exceeding 83%. These results are consistent with previous studies indicating that species such as G. kaustophilus and G. thermoleovorans could be conspecific, suggesting ongoing taxonomic revisions [66]. Phylogenetic analysis corroborates this relationship, showing that Geobacillus sp. G4 aligns within the G. thermoleovorans clade, indicating an evolutionary proximity.
In the genome of G. thermoleovorans G4, key enzymes for the degradation of complex carbohydrates, such as amylases and pullulanase, were identified. These enzymes play essential roles in breaking down starch and pullulan into simpler sugars. These enzymes are widely reported in other Geobacillus strains [57], reflecting the metabolic versatility of this bacterium in starch and sucrose degradation.
The enzymatic classes of hydrolases, transferases, and oxidoreductases are essential for the metabolism and adaptation of Geobacillus to extreme environments. Hydrolases enable the breakdown of biomolecules and the acquisition of nutrients under limited conditions. Transferases regulate biosynthesis and metabolic pathways, facilitating environmental adaptation. Oxidoreductases are crucial for energy production and defense against oxidative stress. The abundance of these enzymes in the G. thermoleovorans sp. G4 genome highlights its ability to adapt and survive in harsh environments.
Concerning the biotechnological potential of G. thermoleovorans, extensive research has focused on identifying enzymes capable of withstanding extreme environmental conditions, such as elevated temperatures and the presence of inhibitory substances [67,68]. Several enzymes from extremophiles have been identified as interesting tools for industrial processes, including biomass deconstruction in biorefinery [69,70].
Here, we describe a new strain of G. thermoleovorans with the potential to produce several industrially relevant enzymes, including more specific endoglucanases (evaluated against carboxymethylcellulose) and pectinases. Cellulases derived from organisms capable of surviving extreme temperatures are extensively documented targets for biotechnological applications because of their ability to withstand prolonged exposure to high temperatures while maintaining catalytic activity [71,72]. The capacity of G. thermoleovorans to grow in the presence of pectin highlights its potential to degrade other complex carbohydrates present in plant cell walls.
Despite an increasing number of reports describing thermophiles that can grow in the presence of cellulose-based polymers, there are few studies describing such organisms as capable of growing in the presence of other complex carbohydrates, such as pectin. The ability of G. thermoleovorans to grow in the presence of pectin may indicate its diverse potential to degrade such complex polymers, which require enzymes with different modes of action, including esterases, lyases, and glycosyl hydrolases [73,74]. To date, the potential of this bacterium to degrade complex carbohydrates has not been explored; however, closely related species have been described as potential producers of pectin lyases [75].
The strain explored in the present study could grow in the presence of starch, which may be a direct consequence of the diversity of starch-degrading enzymes produced by this organism. G. thermoleovorans has been reported to be a good producer of hyperthermostable amylases, including maltogenic α-amylase [76]. The results shown in the present study and previous literature call attention to further heterologous expression and characterization of such enzymes with untapped biotechnological potential.
Genes involved in metabolic categories such as nutrient metabolism, molecule transport, transcription, DNA replication, and repair are fundamental to ensuring the survival and adaptation of Geobacillus sp. G4 in extreme environments [18]. Nutrient metabolism and molecule transport ensure that the bacterium can absorb and process a wide variety of carbon and energy sources, which is essential in habitats with limited resources or adverse conditions. Genes related to transcription and replication ensure the regulation and efficient reproduction of genetic material, while DNA repair genes are crucial for maintaining genomic integrity, especially in environments that expose the organism to stress factors such as heat, radiation, or oxidative compounds [77].
The presence of pathways for glucose-6-phosphate and fructose-6-phosphate metabolism highlights the adaptation of G. thermoleovorans G4 to carbohydrate-rich environments. Furthermore, enzymes associated with cellulose and cellobiose processing indicate a robust capacity to degrade lignocellulosic materials, an important resource for biofuel production, as well as efficient adaptation for energy production via glycolysis, reflecting the metabolic versatility and adaptation of Geobacillus to various ecological niches, including extreme environments [78].
The G. thermoleovorans G4 strain also harbors genes related to antibiotic resistance, such as the vanY/vanB_cl and vanT/vanG_cl clusters, associated with glycopeptide resistance [79], with similarity ranging from 34% to 36% with other Geobacillus species. These findings suggest potential for studies on antimicrobial resistance, which could have important implications for understanding the evolution of bacterial resistance.
This study highlights the identification of seven biosynthetic gene clusters associated with secondary metabolites, including betalactones, terpenes, and, notably, fengycin. Fengycin is a well-known antimicrobial lipopeptide typically produced by mesophilic Bacillus species; however, its occurrence in thermophilic Geobacillus species has not been previously reported [80,81]. Studying fengycin from G. thermoleovorans G4 could uncover novel structural features, enhanced stability, or unique biological activities, highlighting its potential for innovative industrial and pharmaceutical applications.
During carbon starvation, bacteria activate specific mechanisms, including the expression of genes such as cstA, relA, and spoT, to maintain cellular viability. The cstA gene encodes a peptide transporter that is upregulated during nutrient limitation and regulated by CsrA, while relA and spoT mediate the stringent response through the synthesis of (p)ppGpp, which modulates global gene expression and protein synthesis to adapt to stress conditions [82,83,84].
To preserve genomic integrity, G. thermoleovorans G4 possesses conserved DNA repair systems such as nucleotide excision repair (NER) and mismatch repair (MMR). NER, involving uvrA, uvrB, and uvrC, is critical for correcting UV-induced lesions and bulky adducts [85]. Likewise, MMR, mediated by mutS and mutL, ensures replication fidelity by removing base mismatches [86]. Extremophilic organisms have adapted these systems with functional versatility to cope with high-stress environments.
Oxidative stress responses in G. thermoleovorans G4 include enzymes such as superoxide dismutase (sodA), catalase (katA), and alkyl hydroperoxide reductase (ahpC), which detoxify reactive oxygen species (ROS). The overexpression of ahpC, for instance, enhances tolerance to oxidative damage, as shown in other bacterial models [87].
Under heat stress, genes such as dnaK, groEL, and clpXP are upregulated, functioning as molecular chaperones and proteases that maintain protein folding and quality control. These genes are regulated by the transcriptional repressor hrcA, which functions as a protein thermosensor in thermophilic organisms [88].
Finally, these genomic adaptations spanning nutrient limitation, DNA repair, and oxidative and thermal stress responses emphasize the ecological resilience of G. thermoleovorans G4 and its promising value for industrial applications requiring stability under extreme environmental conditions.
5. Conclusions
The genomic and functional data of Geobacillus thermoleovorans G4 demonstrate its value as a highly promising strain for biotechnological applications. Its thermostable enzymes, along with biosynthetic diversity and antibiotic resistance genes, make this strain an important target for biofuel production, industrial biotechnology, and antimicrobial resistance studies.
The detailed analysis of the G. thermoleovorans G4 genome reinforces the importance of combining modern molecular tools, such as ANI, dDDH, and phylogenomics, for accurate bacterial taxonomy and understanding of phylogenetic relationships within the Geobacillus genus. This not only improves the classification of new strains but also facilitates the development of innovative biotechnological solutions based on the adaptive and metabolic capabilities of these extremophilic microorganisms.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/microorganisms13061374/s1, Table S1: The type strains of the genera Bacillus and Geobacillus used to build the phylogenetic tree. Table S2: The complete genomes of type strains from the genera Bacillus and Geobacillus used to construct the phylogenetic tree. Table S3: A list of genes and markers extracted from complete genomes using the AMPHORA2 tool for phylogenetic tree reconstruction. Table S4: The phenotypic characteristics of Geobacillus sp. G4. Table S5: A comparison of the 16S rRNA gene of Geobacillus sp. G4 with the NCBI database. Table S6: An analysis of the annotation of possible antibiotic resistance genes. Table S7: An analysis of biosynthetic gene clusters for secondary metabolites. Table S8: Genes associated with the functional categories: heat-shock response, DNA repair and supercoiling, oxidative stress, and Carbon starvation identified in the genomes of Geobacillus sp. G4, G. kaustophilus HTA426, G. kaustophilus NBRC12445, and G. thermoleovorans KCTC3570. Figure S1: Phylogenomic tree constructed from type strains of Geobacillus and Bacillus species, retrieved from NCBI (GenBank). Figure S2: Metabolic pathways displaying genes and enzymes of interest annotated in the genome of G. thermoleovorans G4.
Author Contributions
Conceptualization, A.R.P.T., F.A.C.L. and R.C.; methodology, A.R.P.T., H.E.R.-A. and K.C.R.S.; software, validation, formal analysis, K.C.R.S., A.R.P.T., F.A.C.L. and R.C.; investigation, H.E.R.-A., A.R.P.T., J.P.S. and P.R.V.H.; resources, A.R.P.T., R.C., P.R.V.H. and E.F.N.; data curation, A.R.P.T. and K.C.R.S.; writing—original draft preparation, K.C.R.S. and A.R.P.T.; writing—review and editing, A.R.P.T., E.F.N., R.C., P.R.V.H. and F.A.C.L.; visualization, H.E.R.-A. and J.P.S.; supervision, E.F.N. and F.A.C.L.; project administration, R.C. and A.R.P.T.; funding acquisition, A.R.P.T., E.F.N. and F.A.C.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research and APC was funded by the Jorge Basadre Grohmann National University through “Fondos del canon sobrecanon y regalias mineras”, approved by Rectoral Resolution N°11174-2023-UNJBG, under the project “Aplicación de tecnología del ADN recombinante en la generación de cocteles enzimáticos para la biodegradación de residuo orgánico vegetal”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding authors.
Acknowledgments
Alonso R. Poma Ticona acknowledges the financial support of CONCYTEC through the PROCIENCIA Program, within the framework of Contest E067-2022-042 (contract No. PE501080334-2022-PROCIENCIA). Kárita C. R. Santos was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES). Fabyano A. C. Lopes was supported by Fundação de Amparo à Pesquisa do Tocantins (FAPT). E.F Noronha is a recipient of CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico) research fellowship. Pedro R.V Hamann is a recipient of the CAPES-PIPD postdoctoral fellowship process number 88887.084629/2024-00. Jéssica P. Silva is a recipient of CNPq postdoctoral fellowship process number 381641/2025-02.
Conflicts of Interest
The authors mutually agree on this submission as it is in the present format; there are no conflicts of interest, and neither human nor animal samples were used during the study. Credit is given to the authors who contributed, and the funding agencies are acknowledged.
References
- Najar, I.N.; Thakur, N. A Systematic Review of the Genera Geobacillus and Parageobacillus: Their Evolution, Current Taxonomic Status and Major Applications. Microbiology 2020, 166, 800–816. [Google Scholar] [CrossRef] [PubMed]
- Dumorné, K.; Córdova, D.C.; Astorga-Eló, M.; Renganathan, P. Extremozymes: A Potential Source for Industrial Applications. J. Microbiol. Biotechnol. 2017, 27, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Nazina, T.N.; Tourova, T.P.; Poltaraus, A.B.; Novikova, E.V.; Grigoryan, A.A.; Ivanova, A.E.; Lysenko, A.M.; Petrunyaka, V.V.; Osipov, G.A.; Belyaev, S.S.; et al. Taxonomic Study of Aerobic Thermophilic Bacilli: Descriptions of Geobacillus subterraneus Gen. Nov., Sp. Nov. and Geobacillus uzenensis Sp. Nov. from Petroleum Reservoirs and Transfer of Bacillus stearothermophilus, Bacillus thermocatenulatus, Bacillus thermoleovorans, Bacillus kaustophilus, Bacillus thermodenitrificans to Geobacillus as the new combinations G. stearothermophilus, G. th. Int. J. Syst. Evol. Microbiol. 2001, 51, 433–446. [Google Scholar] [CrossRef]
- LPSN—List of Prokaryotic Names with Standing in Nomenclature. Available online: https://lpsn.dsmz.de/ (accessed on 30 March 2025).
- Coorevits, A.; Dinsdale, A.E.; Halket, G.; Lebbe, L.; de Vos, P.; van Landschoot, A.; Logan, N.A. Taxonomic Revision of the Genus Geobacillus: Emendation of Geobacillus, G. stearothermophilus, G. jurassicus, G. toebii, G. thermodenitrificans and G. Thermoglucosidans (Nom. Corrig., Formerly ‘Thermoglucosidasius’); Transfer of Bacillus thermantarcticus to the genus as G. thermantarcticus comb. nov.; proposal of Caldibacillus debilis gen. nov., comb. nov.; transfer of G. tepidamans to Anoxybacillus as A. tepidamans comb. nov.; and proposal of Anoxybacillus caldiproteolyticus sp. nov. Int. J. Syst. Evol. Microbiol. 2012, 62, 1470–1485. [Google Scholar] [CrossRef]
- Kuisiene, N.; Raugalas, J.; Chitavichius, D. Geobacillus lituanicus Sp. Nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 1991–1995. [Google Scholar] [CrossRef]
- Meintanis, C.; Chalkou, K.I.; Kormas, K.A.; Lymperopoulou, D.S.; Katsifas, E.A.; Hatzinikolaou, D.G.; Karagouni, A.D. Application of RpoB Sequence Similarity Analysis, REP-PCR and BOX-PCR for the Differentiation of Species within the Genus Geobacillus. Lett. Appl. Microbiol. 2008, 46, 395–401. [Google Scholar] [CrossRef]
- Zeigler, D.R. The Geobacillus Paradox: Why Is a Thermophilic Bacterial Genus so Prevalent on a Mesophilic Planet? Microbiology 2014, 160, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Studholme, D.J. Some (Bacilli) like It Hot: Genomics of Geobacillus Species. Microb. Biotechnol. 2015, 8, 40–48. [Google Scholar] [CrossRef]
- Nazina, T.N.; Lebedeva, E.V.; Poltaraus, A.B.; Tourova, T.P.; Grigoryan, A.A.; Sokolova, D.S.; Lysenko, A.M.; Osipov, G.A. Geobacillus gargensis sp. Nov., a Novel Thermophile from a Hot Spring, and the Reclassification of Bacillus vulcani as Geobacillus vulcani Comb. Nov. Int. J. Syst. Evol. Microbiol. 2004, 54, 2019–2024. [Google Scholar] [CrossRef]
- Nazina, T.N.; Sokolova, D.S.; Grigoryan, A.A.; Shestakova, N.M.; Mikhailova, E.M.; Poltaraus, A.B.; Tourova, T.P.; Lysenko, A.M.; Osipov, G.A.; Belyaev, S.S. Geobacillus jurassicus sp. Nov., a New Thermophilic bacterium Isolated from a High-Temperature Petroleum Reservoir, and the Validation of the Geobacillus Species. Syst. Appl. Microbiol. 2005, 28, 43–53. [Google Scholar] [CrossRef]
- Miñana-Galbis, D.; Pinzón, D.L.; Lorén, J.G.; Manresa, À.; Oliart-Ros, R.M. Reclassification of Geobacillus pallidus (Scholz et al. 1988) Banat et al. 2004 as Aeribacillus pallidus Gen. Nov., Comb. Nov. Int. J. Syst. Evol. Microbiol. 2010, 60, 1600–1604. [Google Scholar] [CrossRef] [PubMed]
- Valdez, S.; de la Vega, F.V.; Pairazaman, O.; Castellanos, R.; Esparza, M. Hyperthermophile Diversity Microbes in the Calientes Geothermal Field, Tacna, Peru. Braz. J. Microbiol. 2023, 54, 2927–2937. [Google Scholar] [CrossRef] [PubMed]
- Castellanos, R.; Santos, K.C.R.; Ticona, A.R.P.; Ramirez-arua, H.E.; Bornás-acosta, S.A.; Silva, P.; Hamann, P.R.V.; Lopes, F.A.C. Draft Genome Sequence of Geobacillus sp. Strain G4 Isolated from the Calientes Geothermal Field in Tacna, Peru. Microbiol. Resour. Announc. 2024, 13, e0071724. [Google Scholar] [CrossRef] [PubMed]
- Cruz, V.; Vargas, V.; Matsuda, K. Geochemical Characterization of Thermal Waters in the Calientes Geothermal Field, Tacna, South of Peru. Proc. World Geotherm. Congr. 2010, 1, 1–7. [Google Scholar]
- Vargas, V.; Cruz Pauccara, V.; Antayhua Vera, Y.; Rivera Porras, M.; Cacya Dueñas, L. Estudio Geotérmico Del Campo Calientes; Instituto Geológico, Minero y Metalúrgico–INGEMMET: Lima, Peru, 2012; Volume 48. [Google Scholar]
- Cortez, Y.; Paul, S.; Villena, G.K.; Gutiérrez-Correa, M.; Rodríguez-Herrera, R.; Kharat, A.S. Isolation and Characterization of Cellulase Producing Bacterial Strains from an Amazonian Geothermal Spring in Peru. Microbiol. Res. J. Int. 2016, 15, 1–8. [Google Scholar] [CrossRef]
- Khaswal, A.; Chaturvedi, N.; Mishra, S.K.; Kumar, P.R.; Paul, P.K. Current Status and Applications of Genus Geobacillus in the Production of Industrially Important Products—A Review. Folia Microbiol. 2022, 67, 389–404. [Google Scholar] [CrossRef]
- Suzuki, H. Peculiarities and Biotechnological Potential of Environmental Adaptation by Geobacillus Species. Appl. Microbiol. Biotechnol. 2018, 102, 10425–10437. [Google Scholar] [CrossRef]
- Mol, M.; de Maayer, P. Elucidating the Biotechnological Potential of the Genera Parageobacillus and Saccharococcus through Comparative Genomic and Pan-Genome Analysis. BMC Genom. 2024, 25, 723. [Google Scholar] [CrossRef]
- van Staden, A.D.P.; van Zyl, W.F.; Trindade, M.; Dicks, L.M.T.; Smith, C. Therapeutic Application of Lantibiotics and Other Lanthipeptides: Old and New Findings. Appl. Environ. Microbiol. 2021, 87, e00186-21. [Google Scholar] [CrossRef]
- Brumm, P.J.; De Maayer, P.; Mead, D.A.; Cowan, D.A. Genomic Analysis of Six New Geobacillus Strains Reveals Highly Conserved Carbohydrate Degradation Architectures and Strategies. Front. Microbiol. 2015, 6, 137987. [Google Scholar] [CrossRef]
- De Maayer, P.; Brumm, P.J.; Mead, D.A.; Cowan, D.A. Comparative Analysis of the Geobacillus Hemicellulose Utilization Locus Reveals a Highly Variable Target for Improved Hemicellulolysis. BMC Genom. 2014, 15, 836. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.L. Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Mac Faddin, J.F. Biochemical Tests for Identification of Medical Bacteria, 3rd ed.; Williams and Wilkins: Baltimore, MD, USA, 2000; ISBN 0683053183. [Google Scholar]
- Guta, M.; Abebe, G.; Bacha, K.; Cools, P. Screening and Characterization of Thermostable Enzyme-Producing Bacteria from Selected Hot Springs of Ethiopia. Microbiol. Spectr. 2024, 12, e0371023. [Google Scholar] [CrossRef]
- Ticona, A.R.P.; Ullah, S.F.; Hamann, P.R.V.; Lopes, F.A.C.; Noronha, E.F. Paenibacillus barengoltzii A1_50L2 as a Source of Plant Cell Wall Degrading Enzymes and Its Use on Lignocellulosic Biomass Hydrolysis. Waste Biomass Valorization 2021, 12, 393–405. [Google Scholar] [CrossRef]
- Oluoch, K.R.; Okanta, P.W.; Hatti-Kaul, R.; Mattiasson, B.; Mulaa, F.J. Protease-, Pectinase- and Amylase- Producing Bacteria from a Kenyan Soda Lake. Open Biotechnol. J. 2018, 12, 33–45. [Google Scholar] [CrossRef]
- Sharma, P.; Sharma, N.; Pathania, S.; Handa, S. Purification and Characterization of Lipase by Bacillus methylotrophicus PS3 under Submerged Fermentation and Its Application in Detergent Industry. J. Genet. Eng. Biotechnol. 2017, 15, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the Quality of Microbial Genomes Recovered from Isolates, Single Cells, and Metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality Assessment Tool for Genome Assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Seppey, M.; Manni, M.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness. Methods Mol. Biol. 2019, 1962, 227–245. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Eddy, S.R. Accelerated Profile HMM Searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [PubMed]
- Feldgarden, M.; Brover, V.; Gonzalez-Escalona, N.; Frye, J.G.; Haendiges, J.; Haft, D.H.; Hoffmann, M.; Pettengill, J.B.; Prasad, A.B.; Tillman, G.E.; et al. AMRFinderPlus and the Reference Gene Catalog Facilitate Examination of the Genomic Links among Antimicrobial Resistance, Stress Response, and Virulence. Sci. Rep. 2021, 11, 12728. [Google Scholar] [CrossRef] [PubMed]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The Reference Centre for Bacterial Insertion Sequences. Nucleic Acids Res. 2006, 34, 32–36. [Google Scholar] [CrossRef]
- Bateman, A.; Martin, M.J.; Orchard, S.; Magrane, M.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bye-A-Jee, H.; Cukura, A.; et al. The UniProt Consortium. UniProt: The Universal Protein Knowledgebase in 2023. Nucleic Acids Res. 2023, 51, D523–D531. [Google Scholar] [CrossRef]
- Huerta-Cepas, J.; Szklarczyk, D.; Heller, D.; Hernández-Plaza, A.; Forslund, S.K.; Cook, H.; Mende, D.R.; Letunic, I.; Rattei, T.; Jensen, L.J.; et al. EggNOG 5.0: A Hierarchical, Functionally and Phylogenetically Annotated Orthology Resource Based on 5090 Organisms and 2502 Viruses. Nucleic Acids Res. 2019, 47, D309–D314. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Augustijn, H.E.; Reitz, Z.L.; Biermann, F.; Alanjary, M.; Fetter, A.; Terlouw, B.R.; Metcalf, W.W.; Helfrich, E.J.N.; et al. AntiSMASH 7.0: New and Improved Predictions for Detection, Regulation, Chemical Structures and Visualisation. Nucleic Acids Res. 2023, 51, W46–W50. [Google Scholar] [CrossRef]
- Liu, B.; Pop, M. ARDB—Antibiotic Resistance Genes Database. Nucleic Acids Res. 2009, 37, D443–D447. [Google Scholar] [CrossRef] [PubMed]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A Database Tandem for Fast and Reliable Genome-Based Classification and Nomenclature of Prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A Web Server for Prokaryotic Species Circumscription Based on Pairwise Genome Comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.I.; Miyata, T. MAFFT: A Novel Method for Rapid Multiple Sequence Alignment Based on Fast Fourier Transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. TrimAl: A Tool for Automated Alignment Trimming in Large-Scale Phylogenetic Analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, H.; Saitou, N. On the Maximum Likelihood Method in Molecular Phylogenetics. J. Mol. Evol. 1991, 32, 443–445. [Google Scholar] [CrossRef]
- Kozlov, A.M.; Darriba, D.; Flouri, T.; Morel, B.; Stamatakis, A. RAxML-NG: A Fast, Scalable and User-Friendly Tool for Maximum Likelihood Phylogenetic Inference. Bioinformatics 2019, 35, 4453–4455. [Google Scholar] [CrossRef] [PubMed]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) Moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef]
- Wu, M.; Scott, A.J. Phylogenomic Analysis of Bacterial and Archaeal Sequences with AMPHORA2. Bioinformatics 2012, 28, 1033–1034. [Google Scholar] [CrossRef]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A Comprehensive, Accurate, and Fast Distance-Based Phylogeny Inference Program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef]
- Farris, J.S. Estimating Phylogenetic Trees from Distance Matrices. Am. Nat. 1972, 106, 645–668. [Google Scholar] [CrossRef]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.Y.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-Depth Characterization and Visualization of Bacterial Genomes. Nucleic Acids Res. 2023, 51, W484–W492. [Google Scholar] [CrossRef]
- Hussein, A.H.; Lisowska, B.K.; Leak, D.J. The Genus Geobacillus and Their Biotechnological Potential. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 92, pp. 1–48. [Google Scholar] [CrossRef]
- Meslé, M.M.; Mueller, R.C.; Peach, J.; Eilers, B.; Tripet, B.P. Isolation and Characterization of Lignocellulose-Degrading Geobacillus thermoleovorans from Yellowstone National Park. Appl. Environ. Microbiol. 2022, 88, e00958-21. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, S.; Salleh, A.B.; Rahman, R.N.Z.R.A.; Chor Leow, T.; Oslan, S.N. Expression and Characterization of Geobacillus stearothermophilus Sr74 Recombinant α-Amylase in Pichia Pastoris. Biomed. Res. Int. 2015, 2015, 529059. [Google Scholar] [CrossRef] [PubMed]
- Burhanoğlu, T.; Sürmeli, Y.; Şanlı-Mohamed, G. Identification and Characterization of Novel Thermostable α-Amylase from Geobacillus sp. GS33. Int. J. Biol. Macromol. 2020, 164, 578–585. [Google Scholar] [CrossRef] [PubMed]
- Bhalla, A.; Bischoff, K.M.; Sani, R.K. Highly Thermostable Xylanase Production from A Thermophilic Geobacillus sp. Strain WSUCF1 Utilizing Lignocellulosic Biomass. Front. Bioeng. Biotechnol. 2015, 3, 84. [Google Scholar] [CrossRef]
- Bibra, M.; Kunreddy, V.R.; Sani, R.K. Thermostable Xylanase Production by Geobacillus sp. Strain Duselr13, and Its Application in Ethanol Production with Lignocellulosic Biomass. Microorganisms 2018, 6, 93. [Google Scholar] [CrossRef]
- Shaheen, M.; Shah, A.A.; Hameed, A.; Hasan, F. Influence of Culture Conditions on Production and Activity of Protease from Bacillus subtilis Bs1. Pak. J. Bot. 2008, 40, 2161–2169. [Google Scholar]
- Morabandza, C.J.; Dibangou, V.; Mabika, F.A.S.; Gatse, E.V.; Ngoulou, T.B.; Itsouhou, N.; Nguimbi, E. Optimization of Culture Conditions for Protease Production Using Three Strains of Bacillus. J. Pure Appl. Microbiol. 2021, 15, 621–629. [Google Scholar] [CrossRef]
- Sung, J.; Ganbat, D.; Kim, S.B.; Lee, S.; Lee, D. Complete Genome Sequences of Geobacillus stearothermophilus Strains EF60045 and SJEF4-2 from Korean Hot Springs. Microbiol. Resour. Announc. 2024, 13, e0057324. [Google Scholar] [CrossRef]
- Miyazaki, K.; Hase, E.; Tokito, N. Complete Genome Sequence of Geobacillus sp. Strain E55-1, Isolated from Mine Geyser in Japan. Microbiol. Resour. Announc. 2020, 9, 10–1128. [Google Scholar] [CrossRef]
- Esin, A.; Ellis, T.; Warnecke, T. Horizontal Gene Flow into Geobacillus Is Constrained by the Chromosomal Organization of Growth and Sporulation. bioRxiv 2018, 381442. [Google Scholar] [CrossRef]
- Aliyu, H.; Lebre, P.; Blom, J.; Cowan, D.; De Maayer, P. Phylogenomic Re-Assessment of the Thermophilic Genus Geobacillus. Syst. Appl. Microbiol. 2016, 39, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Sunna, A.; Tokajian, S.; Burghardt, J.; Rainey, F.; Antranikian, G.; Hashwa, F. Identification of Bacillus kaustophilus, Bacillus thermocatenulatus and Bacillus Strain HSR as Members of Bacillus thermoleovorans. Syst. Appl. Microbiol. 1997, 20, 232–237. [Google Scholar] [CrossRef]
- GArcía-Aparicio, M.P.; Ballesteros, I.; González, A.; Oliva, J.; Ballesteros, M.; Negro, M.J. Effect of Inhibitors Released During Steam-Explosion Pretreatment of Barley Straw on Enzymatic Hydrolysis. Appl. Biochem. Biotechnol. 2006, 129, 278–288. [Google Scholar] [CrossRef]
- Duarte, G.C.; Moreira, L.R.S.; Jaramillo, P.M.D.; Filho, E.X.F. Biomass-Derived Inhibitors of Holocellulases. Bioenergy Res. 2012, 5, 768–777. [Google Scholar] [CrossRef]
- Haki, G.D.; Rakshit, S.K. Developments in Industrially Important Thermostable Enzymes: A Review. Bioresour. Technol. 2003, 89, 17–34. [Google Scholar] [CrossRef]
- Dutta, T.; Sahoo, R.; Sengupta, R.; Ray, S.S.; Bhattacharjee, A.; Ghosh, S. Novel Cellulases from an Extremophilic Filamentous Fungi Penicillium citrinum: Production and Characterization. J. Ind. Microbiol. Biotechnol. 2008, 35, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, R.; Yang, X.; Wu, H.; Xu, D.; Tang, Z.; Shen, Q. Thermostable Cellulase Production of Aspergillus fumigatus Z5 under Solid-State Fermentation and Its Application in Degradation of Agricultural Wastes. Int. Biodeterior. Biodegrad. 2011, 65, 717–725. [Google Scholar] [CrossRef]
- Waheeb, M.S.; Elkhatib, W.F.; Yassien, M.A.; Hassouna, N.A. Optimized Production and Characterization of a Thermostable Cellulase from Streptomyces thermodiastaticus Strain. AMB Express 2024, 14, 129. [Google Scholar] [CrossRef]
- Jayani, R.S.; Saxena, S.; Gupta, R. Microbial Pectinolytic Enzymes: A Review. Process Biochem. 2005, 40, 2931–2944. [Google Scholar] [CrossRef]
- Gummadi, S.N.; Panda, T. Purification and Biochemical Properties of Microbial Pectinases—A Review. Process Biochem. 2003, 38, 987–996. [Google Scholar] [CrossRef]
- Demir, N.; Nadaroglu, H.; Tasgin, E.; Adiguzel, A.; Gulluce, M. Purification and Characterization of a Pectin Lyase Produced by Geobacillus stearothermophilus Ah22 and Its Application in Fruit Juice Production. Ann. Microbiol. 2011, 61, 939–946. [Google Scholar] [CrossRef]
- Uma, J.L.; Rao, M.; Satyanarayana, T. Purification and Characterization of a Hyperthermostable and High Maltogenic α-Amylase of an Extreme Thermophile Geobacillus thermoleovorans. Appl. Biochem. Biotechnol. 2007, 142, 179–193. [Google Scholar] [CrossRef]
- Wang, M.; Zhu, H.; Kong, Z.; Li, T.; Ma, L.; Liu, D.; Shen, Q. Pan-Genome Analyses of Geobacillus spp. Reveal Genetic Characteristics and Composting Potential. Int. J. Mol. Sci. 2020, 21, 3393. [Google Scholar] [CrossRef]
- Najar, I.N.; Sharma, P.; Das, R.; Mondal, K.; Singh, A.K.; Radha, A.; Sharma, V.; Sharma, S.; Thakur, N.; Gandhi, S.G.; et al. Exploring Probiotic Potential: A Comparative Genomics and In Silico Assessment of Genes within the Genus Geobacillus. bioRxiv 2024, 1, 2024.05.15.594408. [Google Scholar] [CrossRef]
- Evers, S.; Courvalin, P. Regulation of VanB-Type Vancomycin Resistance Gene Expression by the VanS B-VanR B Two-Component Regulatory System in Enterococcus Faecalis V583. J. Bacteriol. 1996, 178, 1302–1309. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.Z.; Zheng, Q.W.; Wei, T.; Zhang, Z.Q.; Zhao, C.F.; Zhong, H.; Xu, Q.Y.; Lin, J.F.; Guo, L.Q. Isolation and Characterization of Fengycins Produced by Bacillus amyloliquefaciens JFL21 and Its Broad-Spectrum Antimicrobial Potential Against Multidrug-Resistant Foodborne Pathogens. Front. Microbiol. 2020, 11, 579621. [Google Scholar] [CrossRef] [PubMed]
- Markelova, N.; Chumak, A. Antimicrobial Activity of Bacillus Cyclic Lipopeptides and Their Role in the Host Adaptive Response to Changes in Environmental Conditions. Int. J. Mol. Sci. 2025, 26, 336. [Google Scholar] [CrossRef]
- Matin, A. The Molecular Basis of Carbon-Starvation-Induced General Resistance in Escherichia coli. Mol. Microbiol. 1991, 5, 5–10. [Google Scholar] [CrossRef]
- Dubey, A.K.; Baker, C.S.; Suzuki, K.; Jones, A.D.; Pandit, P.; Romeo, T.; Babitzke, P. CsrA Regulates Translation of the Escherichia coli Carbon Starvation Gene, CstA, by Blocking Ribosome Access to the CstA Transcript. J. Bacteriol. 2003, 185, 4450–4460. [Google Scholar] [CrossRef]
- Flardh, K.; Axberg, T.; Albertson, N.H.; Kjelleberg, S. Stringent Control during Carbon Starvation of Marine Vibrio sp. Strain S14: Molecular Cloning, Nucleotide Sequence, and Deletion of the RelA Gene. J. Bacteriol. 1994, 176, 5949–5957. [Google Scholar] [CrossRef]
- Yeung, A.T.; Mattes, W.B.; Oh, E.Y.; Grossman, L. Enzymatic Properties of Purified Escherichia coli UvrABC Proteins. Proc. Natl. Acad. Sci. USA 1983, 80, 6157–6161. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Zhang, L.; Xu, Q.; Zhang, L.; Yu, Y.; Hua, X. The Mismatch Repair System (MutS and MutL) in Acinetobacter baylyi ADP1. BMC Microbiol. 2020, 20, 40. [Google Scholar] [CrossRef] [PubMed]
- Zuo, F.L.; Yu, R.; Khaskheli, G.B.; Ma, H.Q.; Chen, L.L.; Zeng, Z.; Mao, A.J.; Chen, S.W. Homologous Overexpression of Alkyl Hydroperoxide Reductase Subunit C (AhpC) Protects Bifidobacterium Longum Strain NCC2705 from Oxidative Stress. Res. Microbiol. 2014, 165, 581–589. [Google Scholar] [CrossRef]
- Versace, G.; Palombo, M.; Menon, A.; Scarlato, V.; Roncarati, D. Feeling the Heat: The Campylobacter jejuni HrcA Transcriptional Repressor Is an Intrinsic Protein Thermosensor. Biomolecules 2021, 11, 1413. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).