

A Novel Neotropical Bacillus siamensis Strain Inhibits Soil-Borne Plant Pathogens and Promotes Soybean Growth

 , , , , , ,
, , , , , ,  , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Soil-Borne Pathogens and Bacillus siamensis
2.2. Genomic Analysis and Sequencing of Bacillus siamensis
2.3. Phylogenetic Analysis and Average Nucleotide Identity of Bacillus siamensis
2.4. In Vitro Antagonistic Activity of Bacillus siamensis Against Soil Pathogens
2.5. In Vivo Antagonistic Activity of Bacillus siamensis Against Soil Pathogens
2.6. Effect of Bacillus siamensis on Soybean Seed Germination and Growth
2.7. In Vivo Antagonist Bioassay of Bacillus siamensis in Soybean
2.8. Statistical Analysis
3. Results
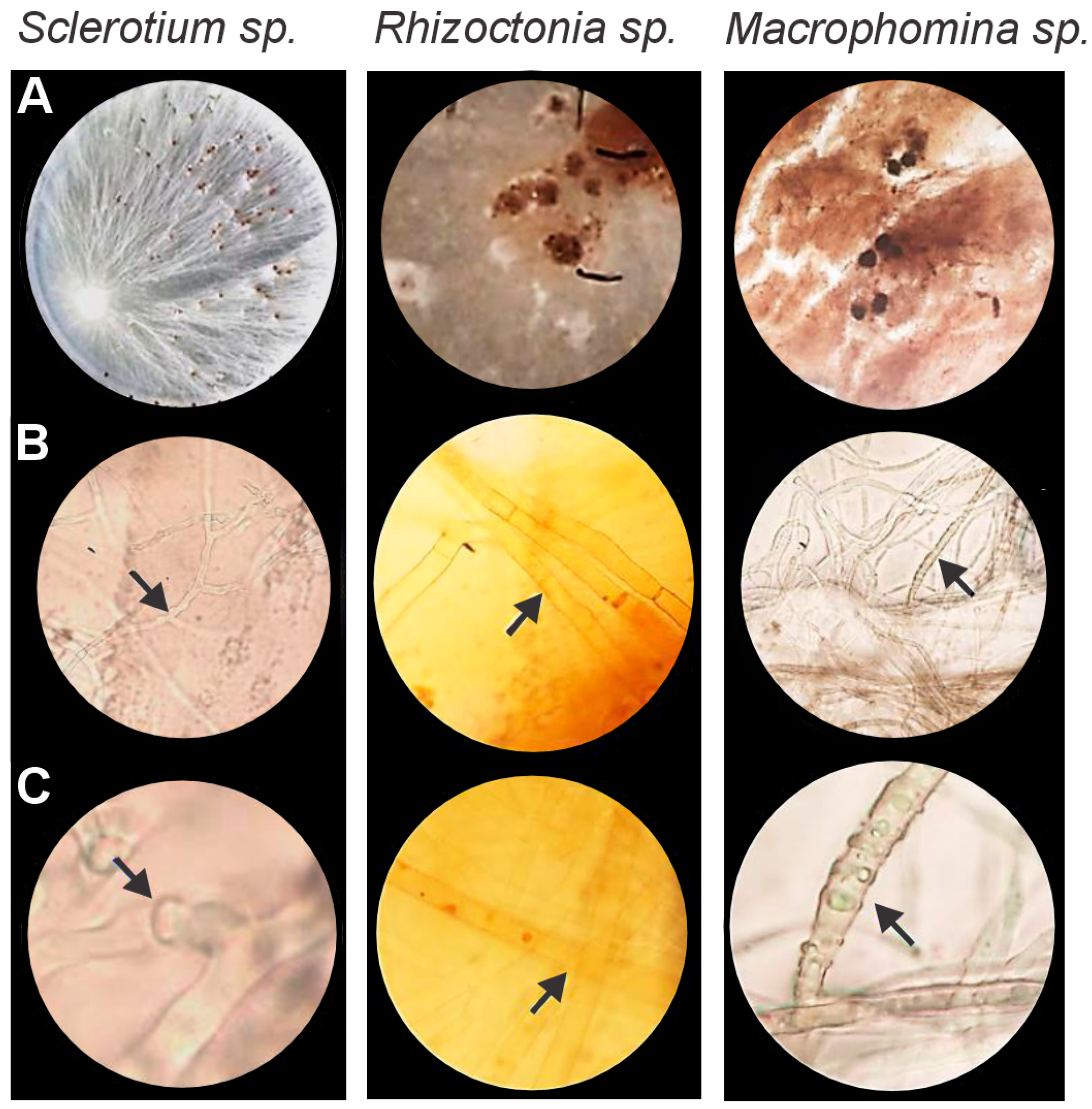
3.1. Identification of Soil-Borne Pathogens
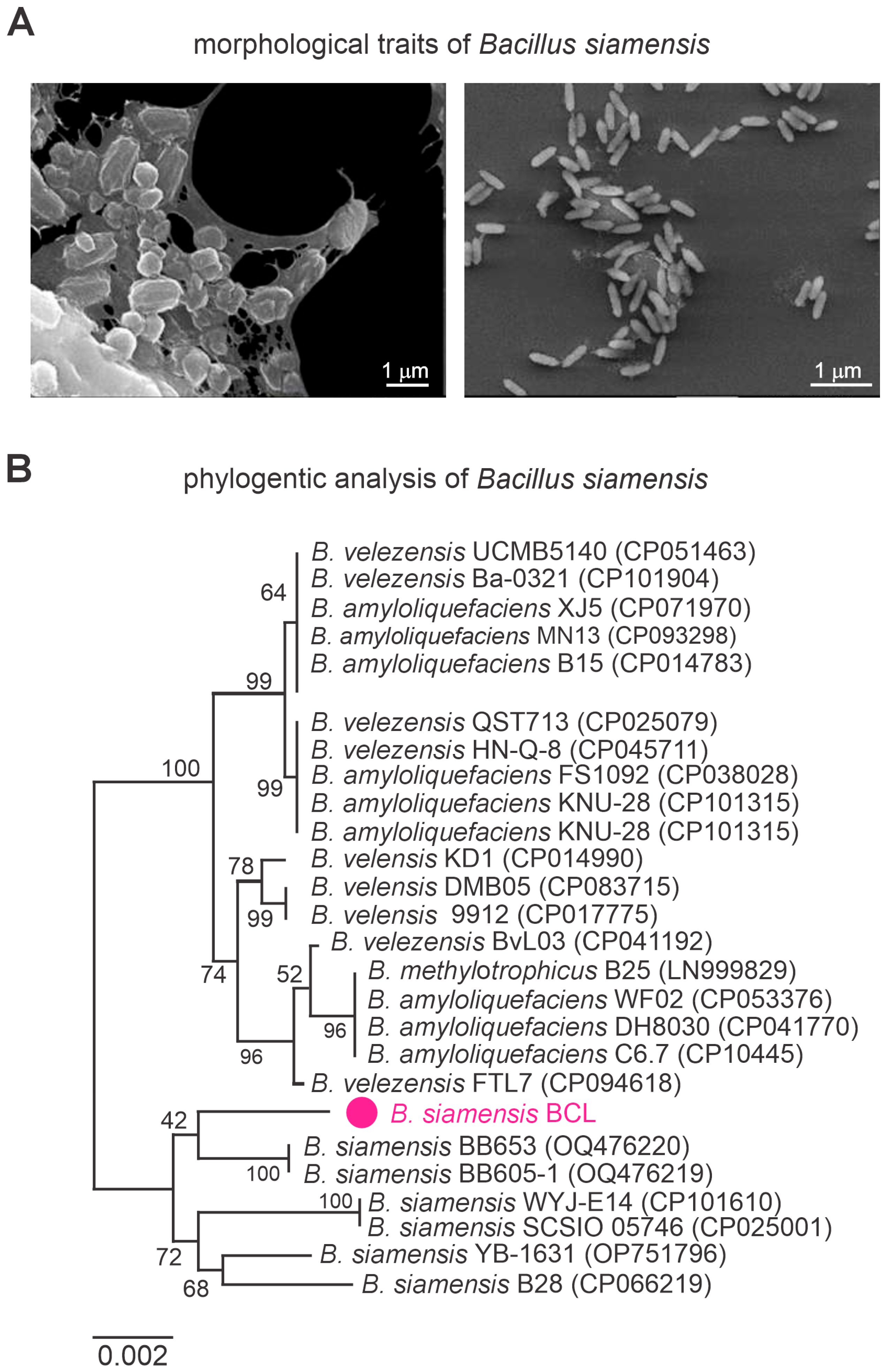
3.2. Characterization of Bacillus siamensis
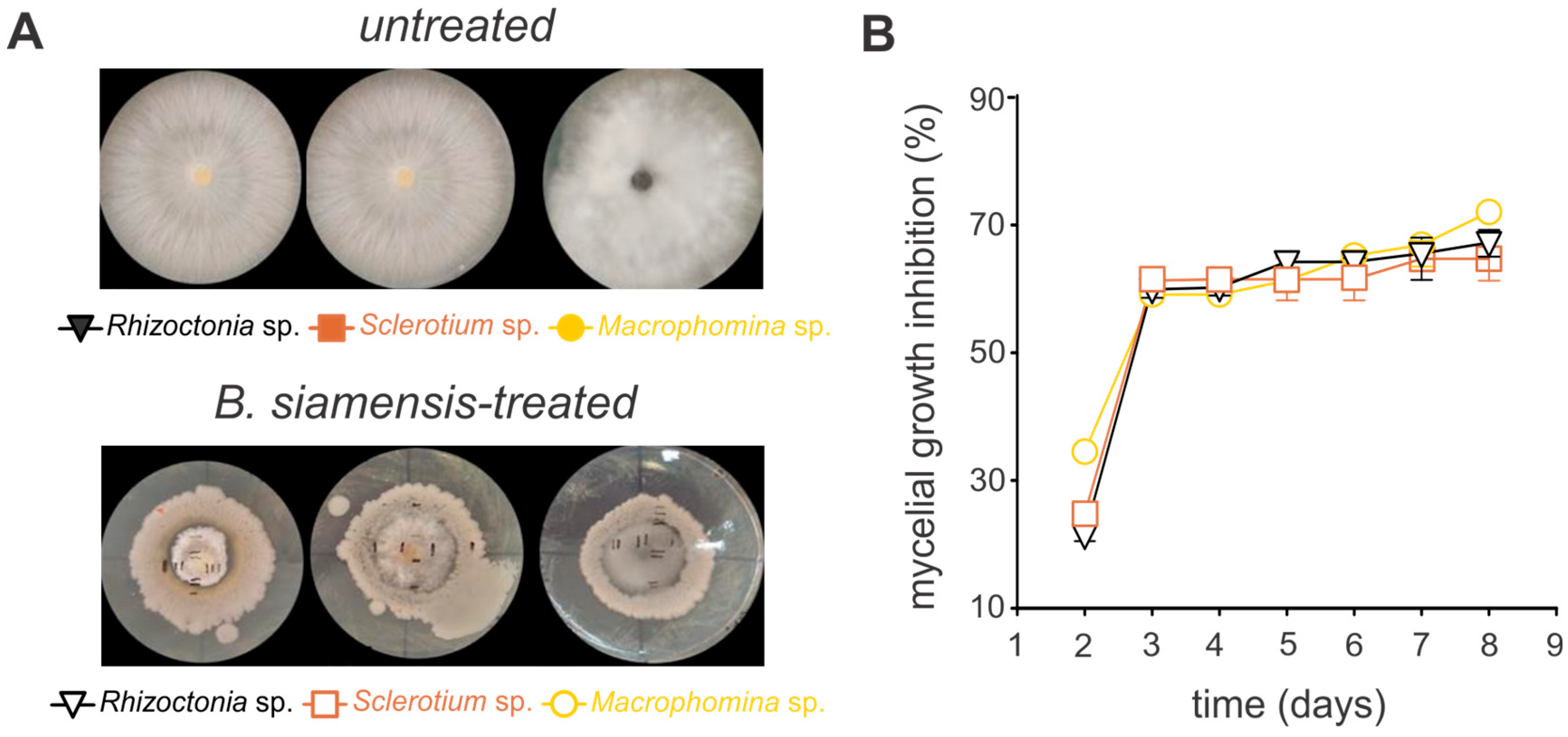
3.3. Antagonistic Activity of Bacillus siamensis
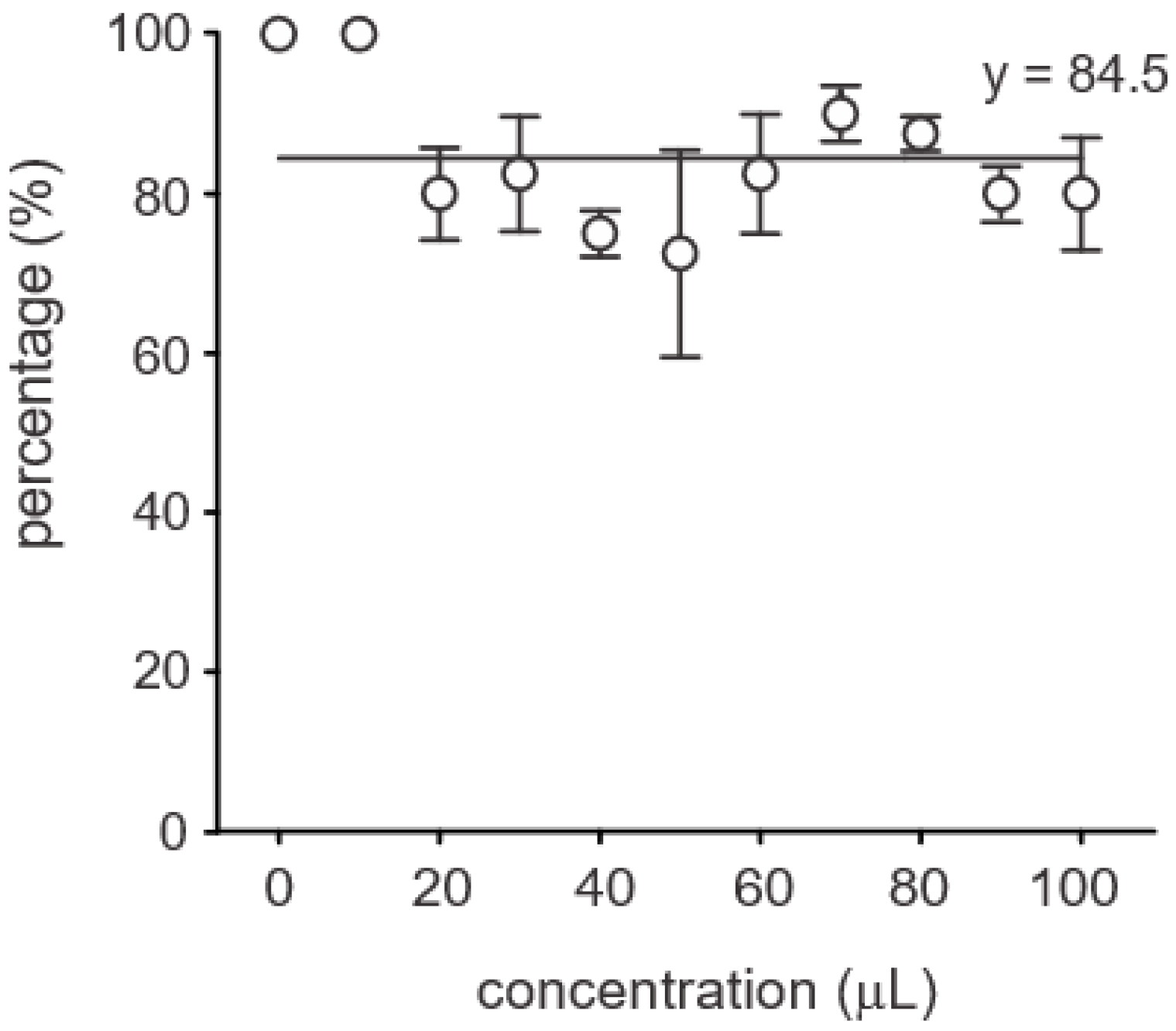
3.4. Effect of Bacillus siamensis on Soybean Growth and Protection
3.5. Genomic Insights into Secondary Metabolites Biosynthesis in Bacillus siamensis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fan, B.; Blom, J.; Klenk, H.-P.; Borriss, R. Bacillus amyloliquefaciens, Bacillus velezensis, and Bacillus siamensis Form an “Operational Group B. amyloliquefaciens” within the B. subtilis Species Complex. Front. Microbiol. 2017, 8, 22. [Google Scholar] [CrossRef] [PubMed]
- Shahniani, A.; Bamzadeh, Z.; Mahmoudnia, F.; Rouhi, L. Evaluation of antibacterial and anticancer properties of secondary metabolites isolated from soil Bacillus spp. focusing on two strains of Bacillus licheniformis and Bacillus siamensis. BMC Mol. Cell Biol. 2024, 25, 21. [Google Scholar] [CrossRef] [PubMed]
- Yassin, S.M.; Aly, A.; Abdel-Kader, D.A.; Morsy, K.; Atallah, O. Antagonistic potential of rhizospheric biocontrol agents against soybean root rot-wilt disease complex syndrome. Zagazig J. Agric. Res. 2019, 46, 1395–1418. [Google Scholar] [CrossRef]
- Pikovskyi, M.; Solomiichuk, M. Identification of mycobiota and diagnosis of soybean seed diseases. Plant Soil Sci. 2022, 13, 44–50. [Google Scholar] [CrossRef]
- Chang, Y.; Dong, Q.; Zhang, L.; Goodwin, P.H.; Xu, W.; Xia, M.; Zhang, J.; Sun, R.; Wu, C.; Wu, K.; et al. Peanut growth promotion and biocontrol of blight by Sclerotium rolfsii with rhizosphere bacterium, Bacillus siamensis YB-1632. Agronomy 2025, 15, 568. [Google Scholar] [CrossRef]
- Hanif, A.; Zhang, F.; Li, P.; Li, C.; Xu, Y.; Zubair, M.; Zhang, M.; Jia, D.; Zhao, X.; Liang, J.; et al. Fengycin produced by Bacillus amyloliquefaciens FZB42 inhibits Fusarium graminearum growth and mycotoxins biosynthesis. Toxins 2019, 11, 295. [Google Scholar] [CrossRef]
- He, H.; Zhai, Q.; Tang, Y.; Gu, X.; Pan, H.; Zhang, H. Effective biocontrol of soybean root rot by a novel bacterial strain Bacillus siamensis HT1. Physiol. Mol. Plant Pathol. 2023, 125, 101984. [Google Scholar] [CrossRef]
- Marquez, N.; Giachero, M.L.; Declerck, S.; Ducasse, D.A. Macrophomina phaseolina: General characteristics of pathogenicity and methods of control. Front. Plant Sci. 2021, 12, 634397. [Google Scholar] [CrossRef]
- Khambhati, V.H.; Abbas, H.K.; Sulyok, M.; Tomaso-Peterson, M.; Chen, J.; Shier, W.T. Mellein: Production in culture by Macrophomina phaseolina isolates from soybean plants exhibiting symptoms of charcoal rot and its role in pathology. Front. Plant Sci. 2023, 14, 1105590. [Google Scholar] [CrossRef]
- Ghag, S.B.; Shekhawat, U.K.; Ganapathi, T.R. Fusarium wilt of banana: Biology, epidemiology and management. Int. J. Pest Manag. 2015, 61, 250–263. [Google Scholar] [CrossRef]
- Hossain, M.M.; Sultana, F.; Li, W.; Tran, L.-S.P.; Mostofa, M.G. Sclerotinia sclerotiorum (Lib.) de Bary: Insights into the pathogenomic features of a global pathogen. Cells 2023, 12, 1063. [Google Scholar] [CrossRef] [PubMed]
- Gorai, P.S.; Ghosh, R.; Ghosh, S.; Samanta, S.; Sen, A.; Panja, S.; Gond, S.K.; Mandal, N.C. Management of black root disease-causing fungus Fusarium solani CRP1 by endophytic Bacillus siamensis CNE6 through its metabolites and activation of plant defense genes. Microbiol. Spectr. 2023, 11, e0308222. [Google Scholar] [CrossRef]
- Afzal, M.; Alghamdi, S.S.; Nawaz, H.; Migdadi, H.H.; Altaf, M.; El-Harty, E.; Al-Fifi, S.A.; Sohaib, M. Genome-wide identification and expression analysis of CC-NB-ARC-LRR (NB-ARC) disease-resistant family members from soybean (Glycine max L.) reveal their response to biotic stress. J. King Saud Univ. Sci. 2022, 34, 101758. [Google Scholar] [CrossRef]
- Díaz-Cruz, G.A.; Cassone, B.J. Changes in the phyllosphere and rhizosphere microbial communities of soybean in the presence of pathogens. FEMS Microbiol. Ecol. 2022, 98, fiac022. [Google Scholar] [CrossRef]
- Matthiesen, R.L.; Robertson, A.E. Effect of infection timing by four Pythium spp. on soybean damping-off symptoms with and without cold stress. Plant Dis. 2023, 107, 3975–3983. [Google Scholar] [CrossRef]
- Wang, X.; Komatsu, S. Subcellular proteomics to elucidate soybean response to abiotic stress. Plants 2023, 12, 2865. [Google Scholar] [CrossRef]
- Chai, M.; Fan, R.; Huang, Y.; Jiang, X.; Wai, M.H.; Yang, Q.; Su, H.; Liu, K.; Ma, S.; Chen, Z.; et al. GmbZIP152, a soybean bZIP transcription factor, confers multiple biotic and abiotic stress responses in plant. Int. J. Mol. Sci. 2022, 23, 10935. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Ma, C.; Fan, Y.; Qiu, Z.; Li, M.; Tian, Y.; Shang, Y.; Liu, C.; Cao, Q.; Peng, Y.; et al. CRISPR-Cas9-mediated editing of GmARM improves resistance to multiple stresses in soybean. Plant Sci. 2024, 346, 112147. [Google Scholar] [CrossRef]
- López-Cardona, N.; Guevara-Castro, A.; Gañán, L. First report of target spot of soybean caused by Corynespora cassiicola in the Colombian eastern plains. Plant Dis. 2021, 105, 490. [Google Scholar] [CrossRef]
- Liu, J.; Cui, W.; Zhao, Q.; Ren, Z.; Li, L.; Li, Y.; Sun, L.; Ding, J. Identification, characterization, and chemical management of Fusarium asiaticum causing soybean root rot in northeast China. Agronomy 2025, 15, 388. [Google Scholar] [CrossRef]
- Bhamra, G.K.; Borah, M.; Borah, P.K. Rhizoctonia aerial blight of soybean, its prevalence and epidemiology: A review. Agric. Rev. 2022, 43, 463–468. [Google Scholar] [CrossRef]
- Kang, I.-J.; Lee, M.; Han, S.Y.; Kim, Y.-H.; Lee, S. First report of soybean root rot caused by Fusarium proliferatum in the republic of Korea. Plant Dis. 2024, 108, 1883. [Google Scholar] [CrossRef]
- Nazarov, P.A.; Baleev, D.N.; Ivanova, M.I.; Sokolova, L.M.; Karakozova, M.V. Infectious Plant Diseases: Etiology, current status, problems and prospects in plant protection. Acta naturae 2020, 12, 46–59. [Google Scholar] [CrossRef]
- Vijayreddy, D. Classification of Plant Diseases, Characteristics and Symptoms on Crop Plants. In Text Book on Practices for Agricultural Sustainability; Golden Leaf Publishers: Lucknow, India, 2024; pp. 171–183. [Google Scholar]
- Lloyd, A.W.; Percival, D.; Langille, M.G.I.; Yurgel, S.N. Changes to soil microbiome resulting from synergetic effects of fungistatic compounds pyrimethanil and fluopyram in lowbush blueberry agriculture, with nine fungicide products tested. Microorganisms 2023, 11, 410. [Google Scholar] [CrossRef]
- Tofan, L.; Niță, V.; Nenciu, M.; Coatu, V.; Lazăr, L.; Damir, N.; Vasile, D.; Popoviciu, D.R.; Brotea, A.-G.; Curtean-Bănăduc, A.M.; et al. Multiple assays on non-target organisms to determine the risk of acute environmental toxicity in tebuconazole-based fungicides widely used in the black sea coastal area. Toxics 2023, 11, 597. [Google Scholar] [CrossRef]
- Baćmaga, M.; Kucharski, J.; Wyszkowska, J. Microbiological and biochemical properties of soil polluted with a mixture of spiroxamine, tebuconazole, and triadimenol under the cultivation of Triticum aestivum L. Environ. Monit. Assess. 2019, 191, 416. [Google Scholar] [CrossRef]
- Duré, L.M.M.; Galeano, R.M.S.; Viana, T.F.C.; Roque, C.G.; Matias, R.; Paggi, G.M.; Corrêa, B.O.; da Silva Brasil, M. Bacillus strains with potential for growth promotion and control of white mold in soybean. Biologia 2022, 77, 3305–3317. [Google Scholar] [CrossRef]
- Conrad, A.M.; Telenko, D.E.P. Efficacy of Biocontrol Agents Coniothyrium minitans and Bacillus amyloliquefaciens for Managing Sclerotinia sclerotiorum in Indiana Soybean. PhytoFrontiers™ 2023, 3, 518–524. [Google Scholar] [CrossRef]
- Hussain, S.; Tai, B.; Ali, M.; Jahan, I.; Sakina, S.; Wang, G.; Zhang, X.; Yin, Y.; Xing, F. Antifungal potential of lipopeptides produced by the Bacillus siamensis Sh420 strain against Fusarium graminearum. Microbiol. Spectr. 2024, 12, e0400823. [Google Scholar] [CrossRef]
- Zhang, M.; Li, X.; Pan, Y.; Qi, D.; Zhou, D.; Chen, Y.; Feng, J.; Wei, Y.; Zhao, Y.; Li, K.; et al. Biocontrol mechanism of Bacillus siamensis sp. QN2MO-1 against tomato fusarium wilt disease during fruit postharvest and planting. Microbiol. Res. 2024, 283, 127694. [Google Scholar] [CrossRef]
- Dimopoulou, A.; Theologidis, I.; Benaki, D.; Koukounia, M.; Zervakou, A.; Tzima, A.; Diallinas, G.; Hatzinikolaou, D.G.; Skandalis, N. Direct antibiotic activity of bacillibactin broadens the biocontrol range of Bacillus amyloliquefaciens MBI600. mSphere 2021, 6, e0037621. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Estebanez, M.; Sanmartín, P.; Camacho-Chab, J.C.; De la Rosa-García, S.C.; Chan-Bacab, M.J.; Águila-Ramírez, R.N.; Carrillo-Villanueva, F.; De la Rosa-Escalante, E.; Arteaga-Garma, J.L.; Serrano, M.; et al. Characterization of a native Bacillus velezensis-like strain for the potential biocontrol of tropical fruit pathogens. Biol. Control 2020, 141, 104127. [Google Scholar] [CrossRef]
- Cui, W.; Zhang, J.; Wang, W.; Wu, X.; Luo, X.; Zou, Y.; Chen, K.; He, P. Screening native Bacillus strains as potential biological control agents against ginger bacterial wilt and for promoting plant growth. Biol. Control 2024, 192, 105510. [Google Scholar] [CrossRef]
- Ikeda, A.C.; Bassani, L.L.; Adamoski, D.; Stringari, D.; Cordeiro, V.K.; Glienke, C.; Steffens, M.B.R.; Hungria, M.; Galli-Terasawa, L.V. Morphological and genetic characterization of endophytic bacteria isolated from roots of different maize genotypes. Microb. Ecol. 2013, 65, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Soares, A.S.; Nascimento, V.L.; de Oliveira, E.E.; Jumbo, L.V.; dos Santos, G.R.; Queiroz, L.L.; da Silva, R.R.; Filho, R.N.A.; Romero, M.A.; de Souza Aguiar, R.W. Pseudomonas aeruginosa and Bacillus cereus Isolated from Brazilian Cerrado Soil Act as Phosphate-Solubilizing Bacteria. Curr. Microbiol. 2023, 80, 146. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Gurevich, A.; Saveliev, V.; Vyahhi, N.; Tesler, G. QUAST: Quality assessment tool for genome assemblies. Bioinformatics 2013, 29, 1072–1075. [Google Scholar] [CrossRef]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Kautsar, S.A.; Medema, M.H.; Weber, T. The antiSMASH database version 3: Increased taxonomic coverage and new query features for modular enzymes. Nucleic Acids. Res. 2020, 49, D639–D643. [Google Scholar] [CrossRef]
- Grant, J.R.; Enns, E.; Marinier, E.; Mandal, A.; Herman, E.K.; Chen, C.-y.; Graham, M.; Van Domselaar, G.; Stothard, P. Proksee: In-depth characterization and visualization of bacterial genomes. Nucleic Acids. Res. 2023, 51, W484–W492. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2015, 32, 929–931. [Google Scholar] [CrossRef]
- Riungu, G.; Muthomi, J.; Narla, R.; Wagacha, J.; Gathumbi, J. Management of Fusarium head blight of wheat and deoxynivalenol accumulation using antagonistic microorganisms. Plant Pathol J. 2008, 7, 13–19. [Google Scholar] [CrossRef]
- Gilbert, G.S.; Diaz, A.; Bregoff, H.A. Seed disinfestation practices to control seed-borne fungi and bacteria in home production of sprouts. Foods 2023, 12, 747. [Google Scholar] [CrossRef]
- Fehr, W.; Caviness, C. Stages of soybean development; Iowa State University of Science and Technology: Ames, IA, USA, 1977; p. 11. [Google Scholar]
- John, E.; Singh, K.B.; Oliver, R.P.; Tan, K.-C. Transcription factor control of virulence in phytopathogenic fungi. Mol. Plant Pathol. 2021, 22, 858–881. [Google Scholar] [CrossRef]
- Santos, A.M.; Soares, A.; Luz, J.; Cordeiro, C.; Sousa Silva, M.; Dias, T.; Melo, J.; Cruz, C.; Carvalho, L. Microbial interactions as a sustainable tool for enhancing pgpr antagonism against phytopathogenic fungi. Sustainability 2024, 16, 2006. [Google Scholar] [CrossRef]
- Shafique, H.A.; Sultana, V.; Ehteshamul-Haque, S.; Athar, M. Management of soil-borne diseases of organic vegetables. J. Plant Prot. Res. 2016, 56, 221–230. [Google Scholar] [CrossRef]
- Botcazon, C.; Bergia, T.; Lecouturier, D.; Dupuis, C.; Rochex, A.; Acket, S.; Nicot, P.; Leclère, V.; Sarazin, C.; Rippa, S. Rhamnolipids and fengycins, very promising amphiphilic antifungal compounds from bacteria secretomes, act on Sclerotiniaceae fungi through different mechanisms. Front. Microbiol. 2022, 13, 977633. [Google Scholar] [CrossRef]
- Zanatta, T.P.; Kulczynski, S.M.; Guterres, C.W.; Fontana, D.C.; Meira, D.; Ceolin, E.L.; Balem, E.; Trevisan, M.; Paraginski, J.A.; Buffon, P.A. Morphological and patogenic characterization of Sclerotinia sclerotiorum. J. Agric. Sci. 2019, 11, 302–313. [Google Scholar] [CrossRef]
- Nasimi, Z.; Barriuso, J.; Keshavarz, T.; Zheng, A. Molecular, physiological, and biochemical properties of sclerotia metamorphosis in Rhizoctonia solani. Fungal Biol. Rev. 2024, 48, 100351. [Google Scholar] [CrossRef]
- Almeida-Silva, F.; Venancio, T.M. Integration of genome-wide association studies and gene coexpression networks unveils promising soybean resistance genes against five common fungal pathogens. Sci. Rep. 2021, 11, 24453. [Google Scholar] [CrossRef]
- Zhao, X.; Ni, Y.; Liu, X.; Zhao, H.; Wang, J.; Chen, Y.-C.; Chen, W.; Liu, H. A simple and effective technique for production of pycnidia and pycnidiospores by Macrophomina phaseolina. Plant Dis. 2020, 104, 1183–1187. [Google Scholar] [CrossRef] [PubMed]
- Coque, J.J.R.; Álvarez-Pérez, J.M.; Cobos, R.; González-García, S.; Ibáñez, A.M.; Diez Galán, A.; Calvo-Peña, C. Chapter Four—Advances in the control of phytopathogenic fungi that infect crops through their root system. In Advances in Applied Microbiology; Gadd, G.M., Sariaslani, S., Eds.; Academic Press: Cambridge, MA, USA, 2020; Volume 111, pp. 123–170. [Google Scholar]
- Hung, S.-H.W.; Yeh, P.-H.; Huang, T.-C.; Huang, S.-Y.; Wu, I.-C.; Liu, C.-H.; Lin, Y.-H.; Chien, P.-R.; Huang, F.-C.; Ho, Y.-N.; et al. A cyclic dipeptide for salinity stress alleviation and the trophic flexibility of endophyte provide insights into saltmarsh plant–microbe interactions. ISME Communications 2024, 4, ycae041. [Google Scholar] [CrossRef]
- Abd-Elsalam, K.A.; Alghuthaymi, M.A.; Abdel-Momen, S.M. Biofungicides: Eco-Safety and Future Trends: Novel Sources and Mechanisms; CRC Press: Boca Raton, FL, USA, 2023; Volume 2. [Google Scholar]
- Ajuna, H.B.; Hyo-In, L.; Jae-Hyun, M.; Sang-Jae, W.; Vantha, C.; Su-In, C.; Ju-Yeol, Y.; Ahn, Y.S. The prospect of antimicrobial peptides from Bacillus species with biological control potential against insect pests and diseases of economic importance in agriculture, forestry and fruit tree production. Biotechnol. Biotechnol. Equip. 2024, 38, 2312115. [Google Scholar] [CrossRef]
- Woo, J.M.; Kim, H.S.; Lee, I.K.; Byeon, E.J.; Chang, W.J.; Lee, Y.S. Potentiality of beneficial microbe Bacillus siamensis GP-P8 for the suppression of anthracnose pathogens and pepper plant growth promotion. Plant Pathol. J. 2024, 40, 346–357. [Google Scholar] [CrossRef]
- Osorio, P.R.A.; Dias, F.R.; Mourão, D.S.C.; Araujo, S.H.C.; Toledo, P.F.S.; Silva, A.C.F.; Viera, W.A.S.; Câmara, M.P.S.; Moura, W.S.; Aguiar, R.W.A.; et al. Essential oil of Noni, Morinda citrifolia L., fruits controls the rice stem-rot disease without detrimentally affect beneficial fungi and ladybeetles. Ind. Crop. Prod. 2021, 170, 113728. [Google Scholar] [CrossRef]
- Barros, A.M.; Ferreira, T.P.d.S.; Mourão, D.d.S.C.; Ferreira, T.P.d.S.; Lima, C.S.L.; Barbosa, R.d.S.; Battistelli, L.C.; Santos, G.R.d. Utilização da farinha e óleo de noni no controle alternativo de Rhizoctonia solani em soja/ Use of flour and noni oil in the alternative control of Rhizoctonia solani in soy. Braz. J. Dev. 2021, 7, 36069–36079. [Google Scholar] [CrossRef]
- Santos Santos, G.R.; Mourão, D.d.S.C.; Han, J.S.; Osório, P.R.A.; da Cruz Araújo, S.H.; Lima, L.R. Potencial fungitóxico dos óleos essenciais de plantas do cerrado no controle dos fitopatógenos Curvularia lunata e Rhizoctonia solani. Peer Rev. 2023, 5, 112–127. [Google Scholar] [CrossRef]
- Souza, W.; Oliveira, L.; Aguiar, R.W.; Falcão, D.; Machado, I.E.; Fidelis, R. Microbiolization with Bacillus sp. in irrigated rice seed positively impacts physiological and phytosanitary quality. Rev. Cienc. Agríc. 2023, 40, e3221. [Google Scholar] [CrossRef]
- Chen, W.; Wu, Z.; Liu, C.; Zhang, Z.; Liu, X. Biochar combined with Bacillus subtilis SL-44 as an eco-friendly strategy to improve soil fertility, reduce Fusarium wilt, and promote radish growth. Ecotoxicol. Environ. Saf. 2023, 251, 114509. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Bishnoi, S.K.; Chandra, S. Assessment of antibiosis potential of Bacillus sp. against the soil-borne fungal pathogen Sclerotium rolfsii Sacc. (Athelia rolfsii (Curzi) Tu & Kimbrough). Egypt. J. Biol. Pest Control 2021, 31, 17. [Google Scholar] [CrossRef]
- Gholami, M.; Reza, K.; Niknam, G. Introduction of some new endophytic bacteria from Bacillus and Streptomyces genera as successful biocontrol agents against Sclerotium rolfsii. Arch. Phytopathol. Plant Prot. 2014, 47, 122–130. [Google Scholar] [CrossRef]
- Castaldi, S.; Petrillo, C.; Donadio, G.; Piaz, F.D.; Cimmino, A.; Masi, M.; Evidente, A.; Isticato, R. Plant growth promotion function of Bacillus sp. strains isolated from salt-pan rhizosphere and their biocontrol potential against Macrophomina phaseolina. Int. J. Mol. Sci. 2021, 22, 3324. [Google Scholar] [CrossRef]
- Marroni, I.V. Screening of bacteria of the genus Bacillus for the control of the plant-pathogenic fungus Macrophomina phaseolina. Biocontrol Sci. Technol. 2015, 25, 302–315. [Google Scholar] [CrossRef]
- Shen, N.; Li, S.; Li, S.; Zhang, H.; Jiang, M. The siderophore-producing bacterium, Bacillus siamensis Gxun-6, has an antifungal activity against Fusarium oxysporum and promotes the growth of banana. Egypt. J. Biol. Pest Control 2022, 32, 34. [Google Scholar] [CrossRef]
- Dong, Q.; Liu, Q.; Goodwin, P.H.; Deng, X.; Xu, W.; Xia, M.; Zhang, J.; Sun, R.; Wu, C.; Wang, Q.; et al. Isolation and Genome-Based Characterization of Biocontrol Potential of Bacillus siamensis YB-1631 against Wheat Crown Rot Caused by Fusarium pseudograminearum. J. Fungi 2023, 9, 547. [Google Scholar] [CrossRef]
- Xie, Z.; Li, M.; Wang, D.; Wang, F.; Shen, H.; Sun, G.; Feng, C.; Wang, X.; Chen, D.; Sun, X. Biocontrol efficacy of Bacillus siamensis LZ88 against brown spot disease of tobacco caused by Alternaria alternata. Biol. Control 2021, 154, 104508. [Google Scholar] [CrossRef]
- Zehra, A.; Raytekar, N.A.; Meena, M.; Swapnil, P. Efficiency of microbial bio-agents as elicitors in plant defense mechanism under biotic stress: A review. Curr. Res. Microb. Sci. 2021, 2, 100054. [Google Scholar] [CrossRef]
- Alemneh, A.A.; Zhou, Y.; Ryder, M.H.; Denton, M.D. Soil environment influences plant growth-promotion traits of isolated rhizobacteria. Pedobiologia 2022, 90, 150785. [Google Scholar] [CrossRef]
- Fanai, A.; Bohia, B.; Lalremruati, F.; Lalhriatpuii, N.; Lalrokimi; Lalmuanpuii, R.; Singh, P.K.; Zothanpuia. Plant growth promoting bacteria (PGPB)-induced plant adaptations to stresses: An updated review. PeerJ 2024, 12, e17882. [Google Scholar] [CrossRef] [PubMed]
- Kunkel, B.N.; Harper, C.P. The roles of auxin during interactions between bacterial plant pathogens and their hosts. J. Exp. Bot. 2017, 69, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Rangel-Montoya, E.A.; Delgado-Ramírez, C.S.; Sepulveda, E.; Hernández-Martínez, R. Biocontrol of Macrophomina phaseolina using Bacillus amyloliquefaciens strains in cowpea (Vigna unguiculata L.). Agronomy 2022, 12, 676. [Google Scholar] [CrossRef]
- Tan, W.; Yin, Y.; Wen, J. Increasing fengycin production by strengthening the fatty acid synthesis pathway and optimizing fermentation conditions. Biochem. Eng. J. 2022, 177, 108235. [Google Scholar] [CrossRef]
- Khan, M.; Muhammad, S.; Abdullah. High-performance liquid chromatography-based characterization of fengycin produced by Bacillus amyloliquefaciens against Fusarium graminearum and Rhizoctonia solani. Kuwait J. Sci. 2022, 49, 13917. [Google Scholar] [CrossRef]
- Vasait, R.D.; Gore, U.B.; Ahire, K.S. Trichoderma species: Screening and in vitro evaluation of its plant growth promoting potential. J. Mycopathol. Res. 2023, 61, 201–206. [Google Scholar] [CrossRef]
- Sanjaya, A.; Praseptiangga, D.; Zaman, M.; Umiati, V.; Baraja, S. Effect of pH, temperature, and salt concentration on the growth of Bacillus subtilis T9-05 isolated from fish sauce. IOP Conf. Ser. Earth Environ. Sci. 2023, 1200, 012050. [Google Scholar] [CrossRef]
- Elazzazy, A.M.; Mobarki, M.O.; Baghdadi, A.M.; Bataweel, N.M.; Al-Hejin, A.M. Optimization of culture conditions and batch process control for the augmented production of bacteriocin by Bacillus Species. Microorganisms 2024, 12, 651. [Google Scholar] [CrossRef]
- Navarro, M.O.P.; Piva, A.C.M.; Simionato, A.S.; Spago, F.R.; Modolon, F.; Emiliano, J.; Azul, A.M.; Chryssafidis, A.L.; Andrade, G. Bioactive compounds produced by biocontrol agents driving plant health. In Microbiome in Plant Health and Disease: Challenges and Opportunities; Kumar, V., Prasad, R., Kumar, M., Choudhary, D.K., Eds.; Springer Singapore: Singapore, 2019; pp. 337–374. [Google Scholar]
- Llorens, E.; Agustí-Brisach, C. Biocontrol of plant diseases by means of antagonist microorganisms, biostimulants and induced resistance as alternatives to chemicals. Plants 2022, 11, 3521. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, C. Fengycins, cyclic lipopeptides from marine Bacillus subtilis strains, Kill the plant-pathogenic fungus Magnaporthe grisea by inducing reactive oxygen species production and chromatin condensation. Appl. Environ. Microbiol. 2018, 84, e00445-18. [Google Scholar] [CrossRef]
- Xiao, P.; Tian, X.; Zhu, P.; Xu, Y.; Zhou, C. The use of surfactin in inhibiting Botrytis cinerea and in protecting winter jujube from the gray mold. AMB Express 2023, 13, 37. [Google Scholar] [CrossRef] [PubMed]
- Kisil, O.; Trefilov, V.; Sadykova, V.; Zvereva, M.; Kubareva, E. SURFACTIN: Biological activity and the possibility of agriculture application. Prikl. Biohim. I Mikrobiol. 2023, 59, 3–16. [Google Scholar] [CrossRef]
- Fischer, S.; López-Ramírez, V.; Asconapé, J. Historical advancements in understanding bacteriocins produced by rhizobacteria for their application in agriculture. Rhizosphere 2024, 31, 100908. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreira, R.F.; Pires, E.B.E.; Sousa, O.F.; Alves, G.B.; Viteri Jumbo, L.O.; Santos, G.R.; Maia, L.J.; Ribeiro, B.M.; Smagghe, G.; Perino, E.H.B.; et al. A Novel Neotropical Bacillus siamensis Strain Inhibits Soil-Borne Plant Pathogens and Promotes Soybean Growth. Microorganisms 2025, 13, 1366. https://doi.org/10.3390/microorganisms13061366
Moreira RF, Pires EBE, Sousa OF, Alves GB, Viteri Jumbo LO, Santos GR, Maia LJ, Ribeiro BM, Smagghe G, Perino EHB, et al. A Novel Neotropical Bacillus siamensis Strain Inhibits Soil-Borne Plant Pathogens and Promotes Soybean Growth. Microorganisms. 2025; 13(6):1366. https://doi.org/10.3390/microorganisms13061366
Chicago/Turabian StyleMoreira, Rodrigo F., Elizabeth B. E. Pires, Odaiza F. Sousa, Giselly B. Alves, Luis O. Viteri Jumbo, Gil R. Santos, Luís J. Maia, Bergmann M. Ribeiro, Guy Smagghe, Elvio H. B. Perino, and et al. 2025. "A Novel Neotropical Bacillus siamensis Strain Inhibits Soil-Borne Plant Pathogens and Promotes Soybean Growth" Microorganisms 13, no. 6: 1366. https://doi.org/10.3390/microorganisms13061366
APA StyleMoreira, R. F., Pires, E. B. E., Sousa, O. F., Alves, G. B., Viteri Jumbo, L. O., Santos, G. R., Maia, L. J., Ribeiro, B. M., Smagghe, G., Perino, E. H. B., Hausmann, R., Oliveira, E. E., & Aguiar, R. W. S. (2025). A Novel Neotropical Bacillus siamensis Strain Inhibits Soil-Borne Plant Pathogens and Promotes Soybean Growth. Microorganisms, 13(6), 1366. https://doi.org/10.3390/microorganisms13061366