Abstract
The quorum-quenching activity of essential oils (EOs) from Corsican aromatic plants was evaluated using the marine bacterium Aliivibrio fischeri as a model system. Among the eleven EOs screened, Myrtus communis EO showed significant interference with QS-regulated phenotypes (swimming motility, bioluminescence, and biofilm formation). Its activity was compared to Origanum vulgaris EO, known for its high carvacrol content and potent QS inhibition. The fractionation of M. communis EO revealed that its most polar fractions exhibited comparable levels of QS-disrupting activity. These chromatographic fractions significantly affected QS-controlled traits, indicating that minor or less volatile compounds may contribute to, or enhance, the overall bioactivity. Furthermore, M. communis EO and its polar fractions displayed stronger anti-QS effects against A. fischeri than O. vulgaris EO. These results highlight M. communis EO as a promising source of natural QS inhibitors and underscore the importance of exploring both complete EOs and their active fractions. This study supports the valorization of Mediterranean endemic flora as a reservoir of bioactive compounds, tested on a model system A. fischeri, and encourages future research on the potential of Myrtus communis against clinical bacterial isolates and the development of novel anti-virulence strategies.
1. Introduction
The emergence and accumulation of bacterial resistance limit the effectiveness of certain antibiotic treatments. The selection pressure exerted by antibiotic molecules to kill bacteria not only increases resistance but also promotes bacterial exchange. To fight bacterial resistance, one area of research focuses on disrupting bacterial communication. This mechanism, known as quorum-sensing (QS), plays a crucial role in bacterial organization, resistance and virulence. A key element of QS is a family of specific molecules called autoinducers. By competing with or inhibiting autoinducers, the QS mechanism can be disrupted through quorum-quenching activity [1,2]. To study this activity in vitro, specific bacterial models are used, such as Chromobacterium violaceum (a purple-pigmented bacterium) or Aliivibrio fischeri (a bioluminescent bacterium), the latter is the first bacteria in which the QS mechanism was described [3,4,5,6]. This marine symbiotic bacterium of the squid Euprymna scolopes is a Gram-negative bacteria capable of producing lophotrichous flagellar tufts, biofilm, and blue-green luminescence through QS regulation [7,8].
The Vibrionaceae family includes another luminescent bacterium, Vibrio harveyi, the toxin-producing pathogenic bacterium Vibrio cholerae, and a group of opportunistic pathogens that affect both shellfish and humans, including Vibrio parahaemolyticus, Vibrio alginolyticus, Vibrio anguillarum, and Vibrio vulnificus [9,10,11]. All of these bacteria are implicated in gastroenteric infections following shellfish consumption, with Vibrio vulnificus also known as the flesh-eating bacterium. Vibrio vulnificus, like many species of Vibrio, can survive in harsh environments or within the human body, with flagellar development and biofilm formation providing dispersal and protective abilities.
Aliivibrio fischeri is a model to test potentials quorum-quenching activities of natural products such as essential oils, aiming to limit bacterial proliferation, such as biofilm formation on coastal surfaces, ship hull, or abiotic surface (biofouling) and mobility to development sites, which can lead to infections all without increasing antibacterial resistance [7,8,12].
Natural extracts are well known for their biological activities across various fields of application such as cosmetics and pharmaceuticals. Highly concentrated active molecules found in essential oils (EOs) from leaves and flowers, or in honey from bee pollen [13], are also used as preservatives in cosmetic and food products [14,15,16]. Essential oils are complex mixtures of volatile chemical compounds that have been extensively studied for their wide range of biological activities, including antibacterial, antifungal, and antioxidant effects. The antimicrobial activity of EOs results from additive or synergistic interactions between different molecules, which are expressed or extracted through steam distillation, hydrodistillation, or cold pressing. The type of plant, cultivation method, period, and geography directly influence the complexity of essential oil composition [17,18,19]. EOs consist of organic volatile compounds, generally mono- and sesquiterpenes as well as phenylpropanoids, with various functional groups (alcohols, ketones, aldehydes, esters, oxides, etc.) [20]. Monoterpenes are widely represented in EOs and include different functional groups. For example, geraniol, linalool, and carvacrol are monoterpene alcohols, while citral (a combination of geranial and neral) is a monoterpene aldehyde. Other compounds, like limonene, p-cymene, and α-pinene are monoterpene hydrocarbons. The abundance of these compounds varies depending on the plant family, such as monoterpenes from evergreens [21]. The recurring composition of certain plant species, such as Origanum spp. or Thymus spp., with carvacrol or thymol as active molecules, differs only in the ratio of these molecules [22,23,24].
The direct antibacterial activity of EOs, such as Origanum vulgaris or Melaleuca alternifolia, including their ability to disrupt membrane integrity or deplete adenosine triphosphate concentration, is well documented [25,26,27]. EOs are among the preferred mixtures used to study synergistic interactions, such as carvacrol and p-cymene, which demonstrate activity against the foodborne pathogen Vibrio cholerae [28]. EOs from the Mediterranean, particularly those from Corsica such as Inula graveolens or Cistus ladaniferus, have demonstrated antibacterial activity against Staphylococcus or Enterobacter [29,30]. Some, like Mentha suaveolens ssp. insularis, have shown strong quorum-quenching activity due to cis-cis-p-menthenolide, one of the main compounds in the oil, which inhibits biofilm formation and violacein production in Chromobacterium violaceum [31]. Some others, like oregano essential oil and its carvacrol-concentrated fractions, have demonstrated strong potential to reduce the motility of several bacteria associated with urinary tract infections [32]. In most cases, the major compounds of EOs are responsible for the antibacterial activities due to their proportion in the entire extract, but sometimes minor compounds, through synergistic interactions or strong individual activities, may play a crucial role [33].
In this study, the effectiveness of eleven Corsican EOs was evaluated on three mechanisms regulated by the QS system, namely bioluminescence, motility, and biofilm formation, in Aliivibrio fischeri and compared to a carvacrol chemotype Origanum vulgaris EO used as internal control. The EO showing the highest activity for each of the three tested parameters, Myrtus communis, was selected and then fractionated by chromatography to investigate the chemical families’ molecules involved in the anti-QS activity observed.
2. Materials and Methods
2.1. Bioactive Substances
Twelve essential oils were evaluated, including eleven sourced from Corsica and one non-island reference oil, Origanum vulgaris, which is highly concentrated in carvacrol. This reference oil was purchased from the French producer Lumiflor (Aubagne, France) and distilled from the aerial parts of the plant. Two Corsican essential oil producers were selected: Astratella (Lumio, France) and Amuredda (Prunelli-di-Fiumorbu, France). Five of the eleven Corsican essential oils were provided by Astratella, namely Eucalyptus polybractea, Eucalyptus globulus, Rosmarinus officinalis, and Pistacia lentiscus, all distilled from leafy branches. Astratella also supplies the essential oil extracted from the aerial parts of Inula graveolens. The remaining six essential oils were obtained from Amuredda, namely Lippia citriodora, Myrtus communis, Pelargonium asperum, Mentha piperita, and Lavandula hybrida, which were all distilled from leaves, except for Cymbopogon winterianus, which was distilled from the plant’s aerial parts.
2.2. Column Chromatography of Myrtus communis EO
The Myrtus communis leaves essential oil (M. communis EO; 2.563 g) were chromatographed on silica gel (63–200 µm, 60 Å, 53 g) with a gradient of solvent (pentane/diethyl ether from 100/0 to 0/100), yielding four fractions (F1–F4): F1 (100/0; 1.246 g), F2 (98/2; 664.2 mg), F3 (90/10; 53.0 mg), and F4 (0/100; 453.1 mg).
2.3. GC-FID Analysis
GC-FID analyses were carried out using a Clarus 500 Perkin Elmer (Perkin Elmer, Courtaboeuf, France) system equipped with an FID and two fused-silica capillary columns (50 m × 0.22 mm, film thickness 0.25 µm), BP-1 (polydimethylsiloxane), and BP-20 (polyethylene glycol). The oven temperature was programmed from 60 °C to 220 °C at 2 °C/min and then held isothermal at 220 °C for 20 min; injector temperature: 250 °C; detector temperature: 250 °C; carrier gas: H2 (0.8 mL/min); split: 1/60; injected volume: 0.5 µL. The relative proportions of the essential oil constituents were expressed as percentages obtained by peak-area normalization, without using correction factors. Retention indices (RI) were determined relative to the retention times of a series of n-alkanes with linear interpolation (Target Compounds software from Perkin Elmer).
2.4. GC-MS Analysis
GC-MS analyses were performed on a Clarus SQ8S Perkin Elmer TurboMass detector (quadrupole), directly coupled to a Clarus 580 Perkin-Elmer Autosystem XL, equipped with a BP-1 (polydimethylsiloxane) fused-silica capillary column (60 m × 0.22 mm i.d., film thickness 0.25 µm). The oven temperature was programmed from 60 °C to 230 °C at 2°/min and then held isothermally at 230 °C for 45 min; injector temp., 250 °C; ion-source temp., 150 °C; carrier gas, He (1 mL/min); split ratio, 1:80; injection volume, 0.2 µL; ionization energy, 70 eV. The electron ionization (EI) mass spectra were acquired from within the mass range of 35–350 Da.
2.5. Identification of Individual Components
Essential oil samples and fractions of chromatography were submitted to GC, in combination with retention indices (RI) and GC-MS (50 mg of essential oil or fraction CC diluted in 500 µL of CHCl3). Identification of the individual components was carried out as follows: (i) by the comparison of their GC retention indices (RI) on polar and apolar columns with those of reference compounds compiled in a laboratory-built library and with literature data [34,35,36]; (ii) via computer matching against commercial mass spectral libraries [37].
2.6. Bacterial Strains and Growth Conditions
The bacterial strain Aliivibrio fischeri (formerly Vibrio fischeri) was obtained from the Collection Institut Pasteur (CIP 103206T) and cultivated according to an adaptation of the Christensen and Visick protocol [23] at temperatures between 25 and 28 °C in SWTO broth.
2.7. Minimal Inhibitory Concentration Test
The MIC determination was performed in microplates following the Guinoiseau et al. protocol with some adaptations [30]. All essential oils were solubilized in absolute ethanol (VWR, Radnor, PA, USA). Positive growth controls were performed using absolute ethanol in bacterial culture broth, while negative controls consisted of essential oil diluted in sterile culture medium without bacteria. All test solutions were prepared at a final concentration of 1% v/v, corresponding to 2 µL of absolute ethanol or essential oil diluted in 200 µL of culture broth, with or without bacteria. This yielded a final concentration range from 10,000 ppm to 5 ppm. The minimum inhibitory concentration (MIC) determination enabled the identification of a common sub-inhibitory concentration suitable for all essential oils, based on optical density measurements at 600 nm after 24 h of incubation with shaking.
2.8. Swimming Mobility Test
According to the Christensen and Visick protocol [38], SWTO broth was supplemented with 1.25% agar to evaluate swimming and luminescence in semi-liquid media. In accordance with the MIC testing protocol, each well of the microplate was inoculated with 2 µL of essential oil diluted in 200 µL of bacterial broth culture at concentrations between 1 and 2 × 108 CFU·mL−1, yielding a final essential oil concentration of 50 ppm. A 5 µL mixture of bacteria and essential oil was plated in the center of the plate using a center shot. The swimming interval was 6 h (±1 h), with four measurements taken starting approximately 6 h post-inoculation. Bacterial swimming was assessed from inoculation to 24 h of incubation at 28 °C, every 6 h.
2.9. Bioluminescence Perturbation Assay
Luminescence quantification was performed using a Tecan Infinite M Plex (Tecan, Männedorf, Austria) luminometer. The inoculum used for the swimming motility assay was also used for the luminescence kinetics study. The luminescence intervals were set at 10 min throughout a period of 2 h and 30 min at 28 °C. A total of 200 µL of bacterial inoculum were inoculated into each well with 2 µL of diluted essential oils. All tests were performed using white (Nunc, Roskilde, Denmark) and transparent (Greiner, Kremsmünster, Austria) plates to control for OD600 measurements.
2.10. Relative Antibiofilm Quantification
Relative biofilm quantification was performed using the same inoculum as for luminescence after 2 h and 30 min of incubation under the same conditions as described above. After the incubation period, the inoculum was removed, and the microplate was washed three times with phosphate buffer. Once the biofilm was dry, 250 µL of crystal violet (Sigma, St. Louis, MO, USA) solution (0.1%) was added to each well and removed after 5 min. After obtaining clear wash water, 250 µL of 20% ethanol/1% trichloroacetic acid was added to each well, and after color homogenization, the absorbance was read at 595 nm.
2.11. Statistical Analyses
All data were analyzed using the R 3.6.0 and RStudio 4.3.1 statistical software (http://www.R-project.org, accessed on 20 July 2022) and Rstudio (Rstudio Inc., Boston, MA, USA, version 1, April 1717). All data were expressed as mean values (±standard error) and submitted to two-way ANOVA with an LSD test when a significant difference was detected at p < 0.001.
3. Results
3.1. Selection of Essential Oils and Identification of Their Major Compounds
The twelve EOs were analyzed by GC(RI) and GC-MS, and the major compounds are listed in Table 1. The composition of EOs of Eucalyptus is dominated by 1,8-cineole at, respectively, 56.7% for E. globulus and 25.6% for E. polybractea. The main compound in the essential oils of M. communis and R. officinalis is α-pinene, comprising 43% and 42.5%, respectively, along with a notable amount of 1,8-cineole (28.8% and 9.8%). Citral (neral + geranial) is the main constituent of C. winterianus (73.2%) and L. citriodora (23.4%) along with limonene (23.1%) for the final oil. The compositions of the other EOs are characterized by various major compounds: P. asperum (citronellol 24.5%, geraniol 15.1%, and citronellyl formate 10.5%), L. hybrida (linalool 37.3% and linalyl acetate 34.4%), M. piperita (menthol 42.7% and menthone 25.2%), I. graveolens (bornyl acetate 53% and borneol 19.3%), P. lentiscus (α-pinene 25.9% and myrcene 17.5%), and O. vulgaris (carvacrol 62% and terpinolene 10.1%).

Table 1.
Major compounds of the eleven Corsican essential oils and Origanum vulgaris essential oil (internal control).
All these EOs are rich in monoterpenes, primarily oxygenated monoterpenes (alcohols, aldehydes, ketones, ether oxides, and esters) and also monoterpene hydrocarbons like myrcene, limonene, p-cymene, and α-pinene. Among the oxygenated monoterpenes, ether oxides are predominantly represented by 1;8-cineole; aldehydes by citral; ketones by menthone; esters by bornyl acetate; and alcohols by linalool, geraniol, menthol, citronellol, and carvacrol, with the latter phenolic compound being a direct result of p-cymene hydroxylation [29].
3.2. Chemical Composition of Myrtus communis EO and Its Chromatographic Fractions
Based on all previous results and the notable potential of M. communis EO to affect the bioluminescence, mobility, and biofilm formation of A. fischeri, this EO was selected for further investigation into QS inhibition.
The chemical composition of this essential oil is detailed in Table 2. Forty-one compounds were identified, representing 97.3% of the whole composition. The EO contained mainly monoterpenes, namely α-pinene (43.0%) followed by 1,8-cineole (28.8%), limonene (8.7%), linalool (3.8%), p-cymene (2.3%), α-terpineol (1.7%), and geranyl acetate (1.6%), with the other compounds amounting to less than 1% each.
Table 2.
Composition of Myrtus communis essential oil and the four fractions obtain after chromatography.
To investigate the anti-QS activities of M. communis EO, this sample was fractionated to separate molecules based on their polarity. It was submitted to column chromatography (CC) over silica gel using a gradient of solvent pentane/diethyl ether (100/0 to 0/100) and the four fractions (F1–F4) obtained were analyzed by GC(RI) and GS-SM.
The first fraction (F1), eluted with pentane, consists essentially of monoterpene hydrocarbons, mainly α-pinene (70.4%) and limonene (20.1%), which represent 43% and 8.7% of the composition of the whole essential oil, respectively. Fraction F2 (pentane/diethyl ether, 98/2) is composed of 94.2% oxygenated monoterpenes, with 80.8% of 1,8-cineole, the second major component of the EO. F1 and F2 have the highest identification percentages, at 99.2% and 98.3%, respectively. The next two fractions, F3 and F4, which are more complex, are identified at 61.1% and 63.3%.
The F3 fraction (pentane/diethyl ether, 90/10) is the most heterogeneous, containing 41.6% of oxygenated sesquiterpenes, 10.4% of phenylpropanoids (“Others” in Table 2), and 9.1% of oxygenated monoterpenes. The three major compounds of this fraction are caryophyllene oxide (33.5%), methyl eugenol (10.4%), and humulene epoxide II (8.1%), representing 0.3%, 0.2%, and 0.1% of M. communis EO, respectively.
F4, eluted with diethyl ether, is composed of 62.5% oxygenated monoterpenes, with small amounts of oxygenated sesquiterpenes (0.2%) and phenylpropanoids (0.6%). The major compounds in this fraction are linalool (27.5%) and α-terpineol (17.6%), two monoterpene alcohols, which account for 3.8% and 1.7% of M. communis EO, respectively.
3.3. Swimming Assay
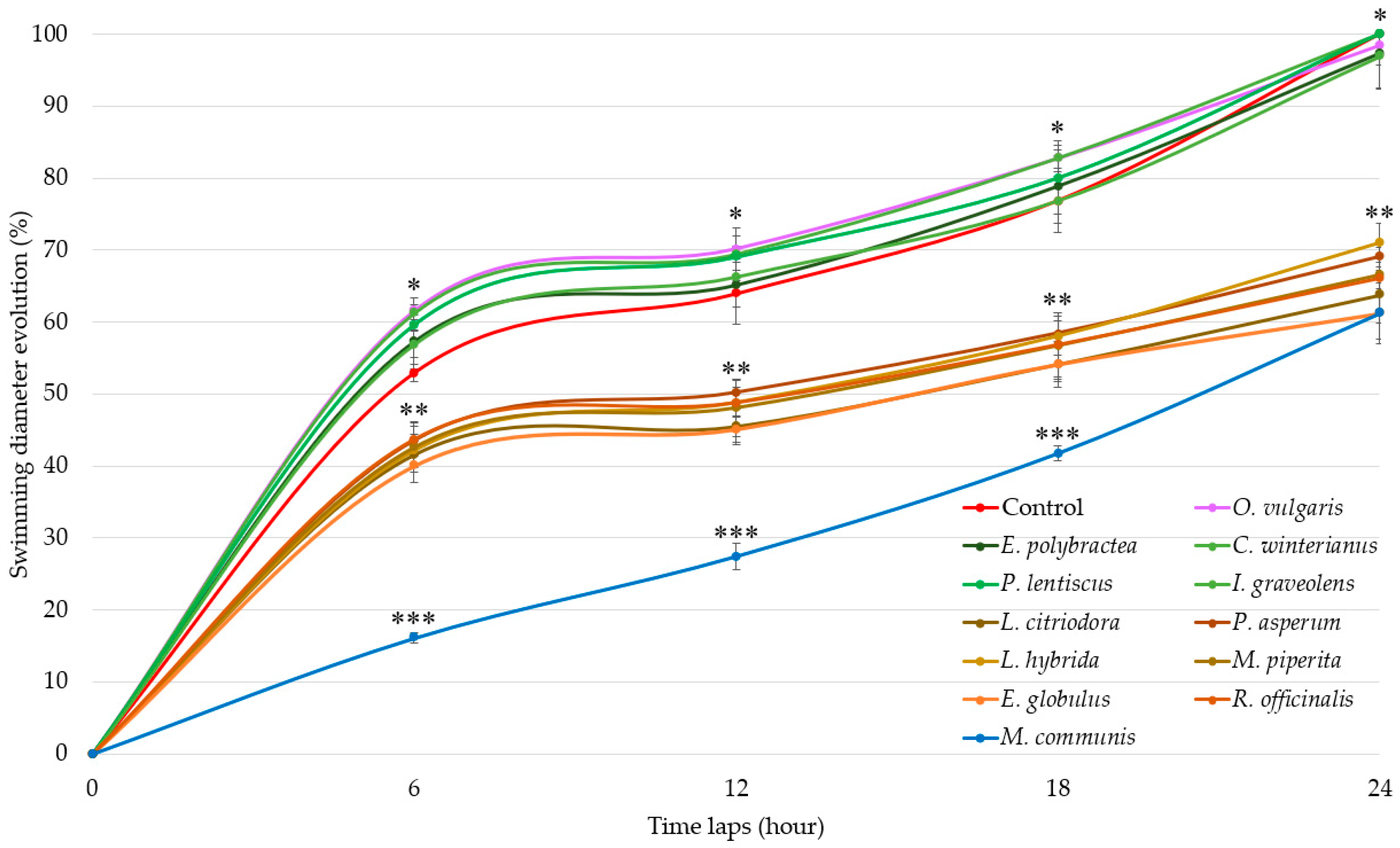
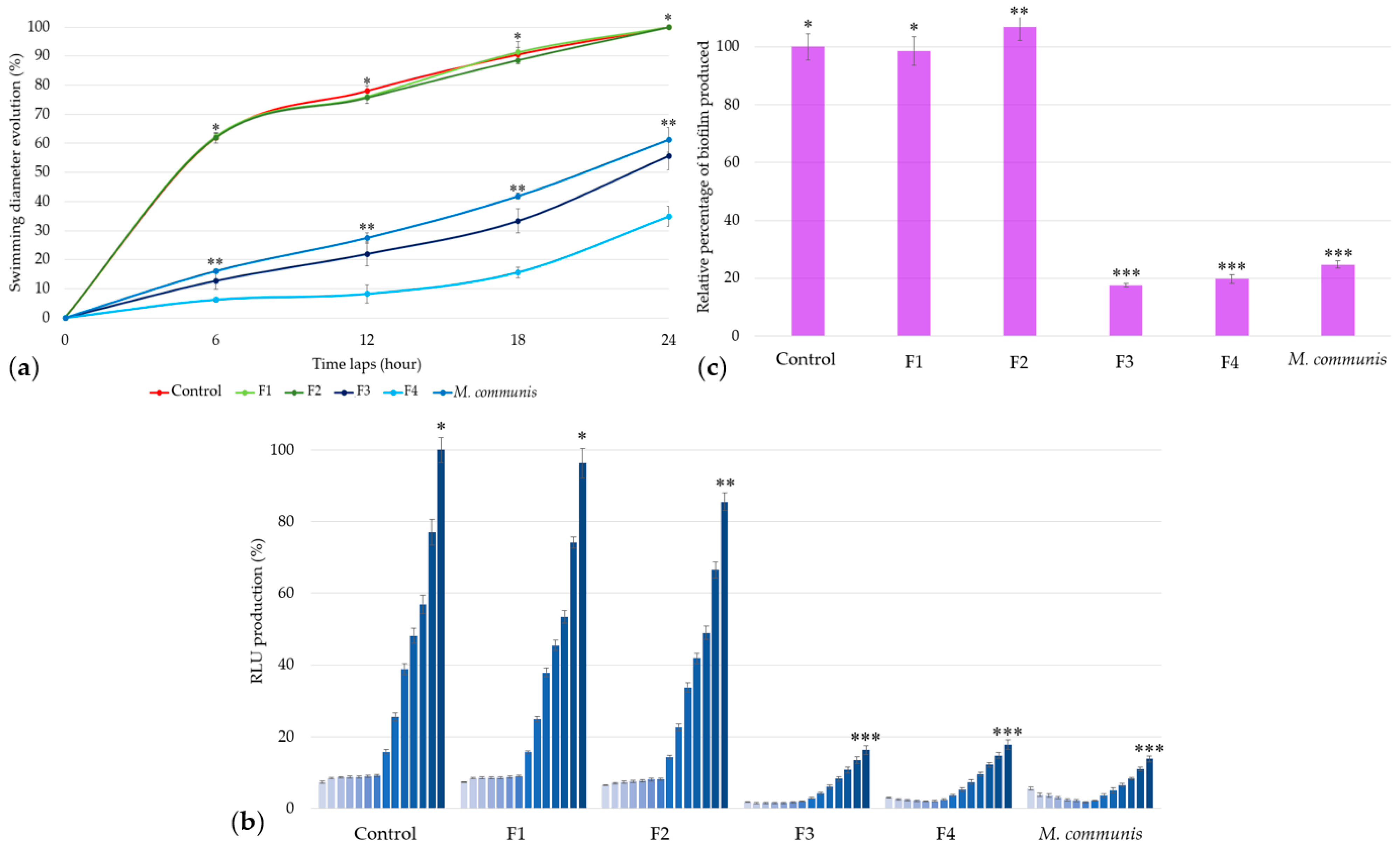
Swimming results (Figure 1) show three distinct groups of EOs activities.
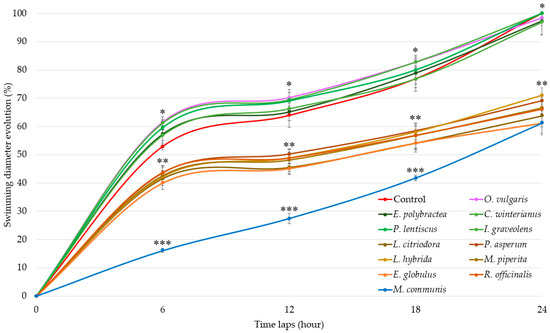
Figure 1.
Swimming diameter of Aliivibrio fischeri after treatment with or without (control) essential oils during four time laps. EO data are mean values (±SD) of at least four independent measurements (n = 3); different (*), (**) and (***) indicate significant differences during each time lap (p < 0.001).
EOs of G1 (E. polybractea, C. winteranius, I. graveolens, P. lentiscus, and O. vulgaris) did not influence A. fischeri motility as compared to the control. In contrast, all the other EOs lead to a significant decrease in swimming diameters. The EOs of G2 (E. globulus, R. officinalis, L. citriodora, P. asperum, L. hybrida, and M. piperita) decrease swimming diameter by around 60% at 6 h, 55% at 12 h, 45% at 18 h, and finally by 35% at the end of the 24 h measurement. The most significant delay was observed with M. communis EO (blue), which demonstrated stronger motility inhibition starting at 6 h with 36.86% (±0.68%), increasing to 42.94% (±1.80%) at 12 h, then rising further to 54.34% (±1.02%) at 18 h before decreasing to 38.82% (±4.24%) at 24 h, similarly to the G2-active EOs.
3.4. Bioluminescence Assay
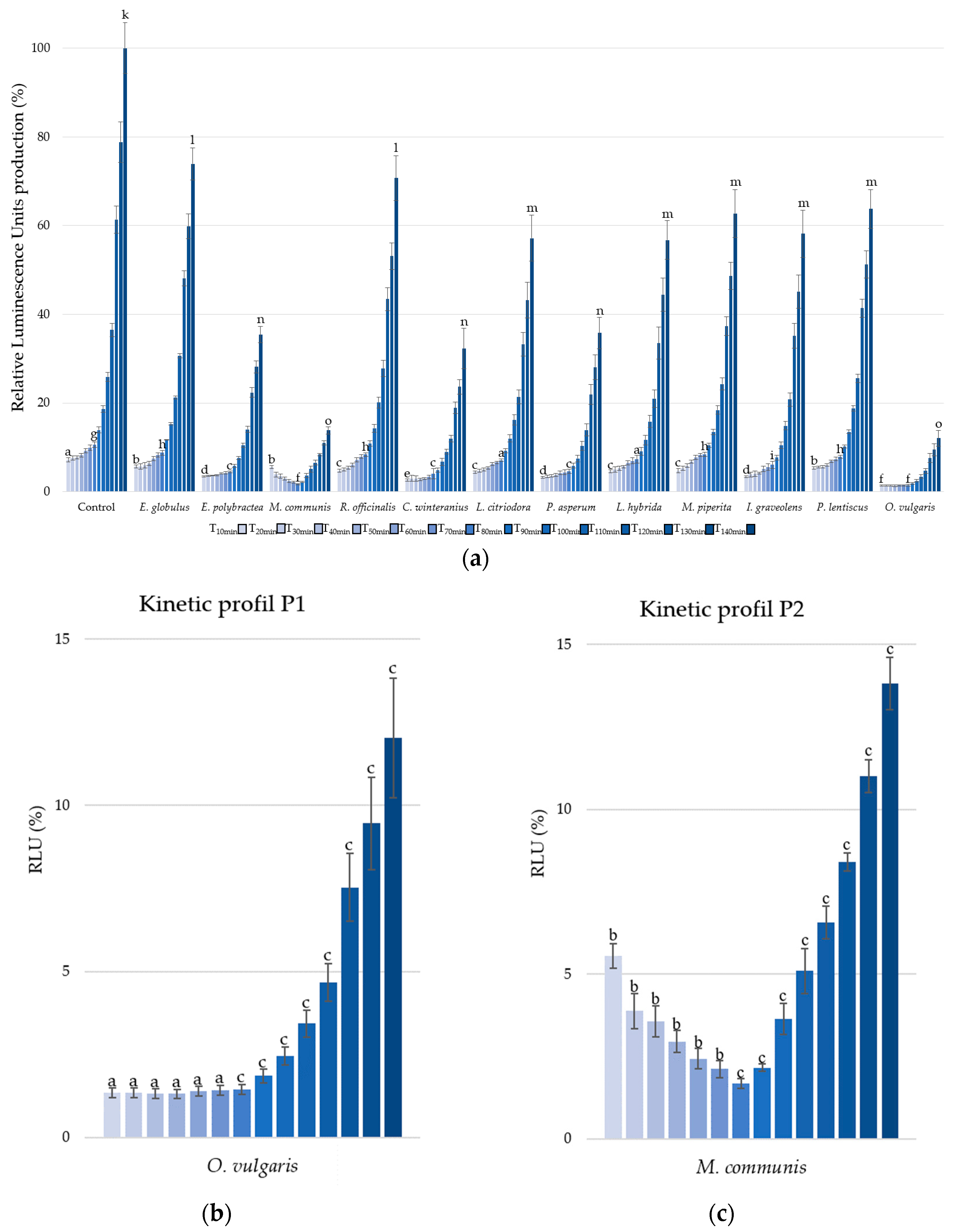
The second parameter, bioluminescence production, was measured under kinetic conditions from T10 minutes to T140 minutes, with 10 min intervals (Figure 2a).
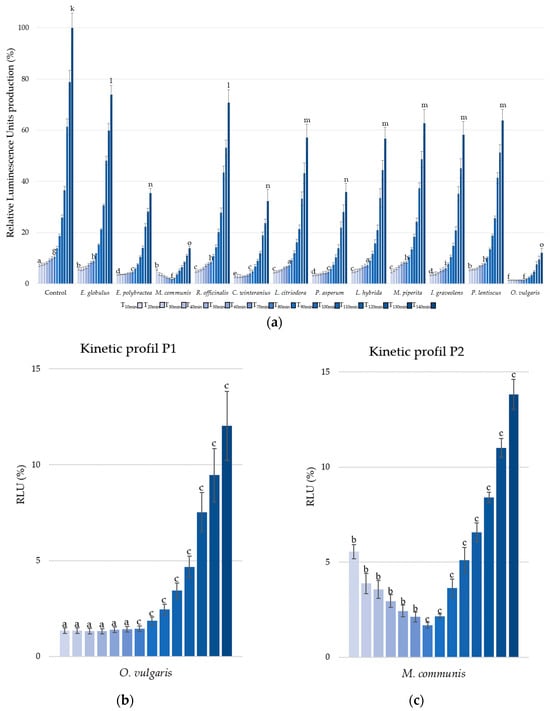
Figure 2.
Bioluminescence production of Aliivibrio fischeri after treatment with essential oils or without (control). Data are mean values (±SD) of at least four independent measurements (n ≥ 4); different lowercase letters indicate significant differences between each EO at three points during the kinetic cycles (1, 7, and 14) (p < 0.001) (a). Kinetic profiles in bioluminescence inhibition with O. vulgaris (b) and M. communis (c) EOs. Data are mean values (±SD) of at least four independent measurements (n ≥ 4) different lowercase letters indicate significant differences between the two EOs at each kinetic cycle (p < 0.001).
Only three other EOs present significative reduction in RLU, i.e., E. polybractea, C. winteranius, and P. asperum with respective luminescence reduction of 51.98% (±1.39%), 59.91% (±3.62%), and 55.34% (±3.33%) at T10 minutes. At the end of the kinetic cycle, E. polybractea, C. winteranius, and P. asperum EOs decrease, respectively, RLU by 64.59% (±1.85%) 67.71% (±4.62%), and 64.21% (±3.51%).
But the most significant activities were observed with EOs of O. vulgaris and M. communis, showing respective luminescence decreases of 86.28% (±0.16%) and 84.17% (±0.15%).
Furthermore, it is important to note that all the EOs and growth controls exhibited similar kinetic profiles, characterized by a stagnation in luminescence followed by an increase, with the sole exception of M. communis EO.
Indeed, to compare the kinetic profiles, O. vulgaris EO (Figure 2b) was chosen to represent P1 profile, while M. communis EO, the only P2 profile, was selected (Figure 2c). Two distinct trends are observed in the A. fischeri inhibition of bioluminescence production: the P1-Type, characterized by a stagnation in luminescence production (T1 to T5) before an increase, and the P2-Type, which shows a reduction (T1 to T5) in luminescence production followed by an increase.
3.5. Biofilm Assay
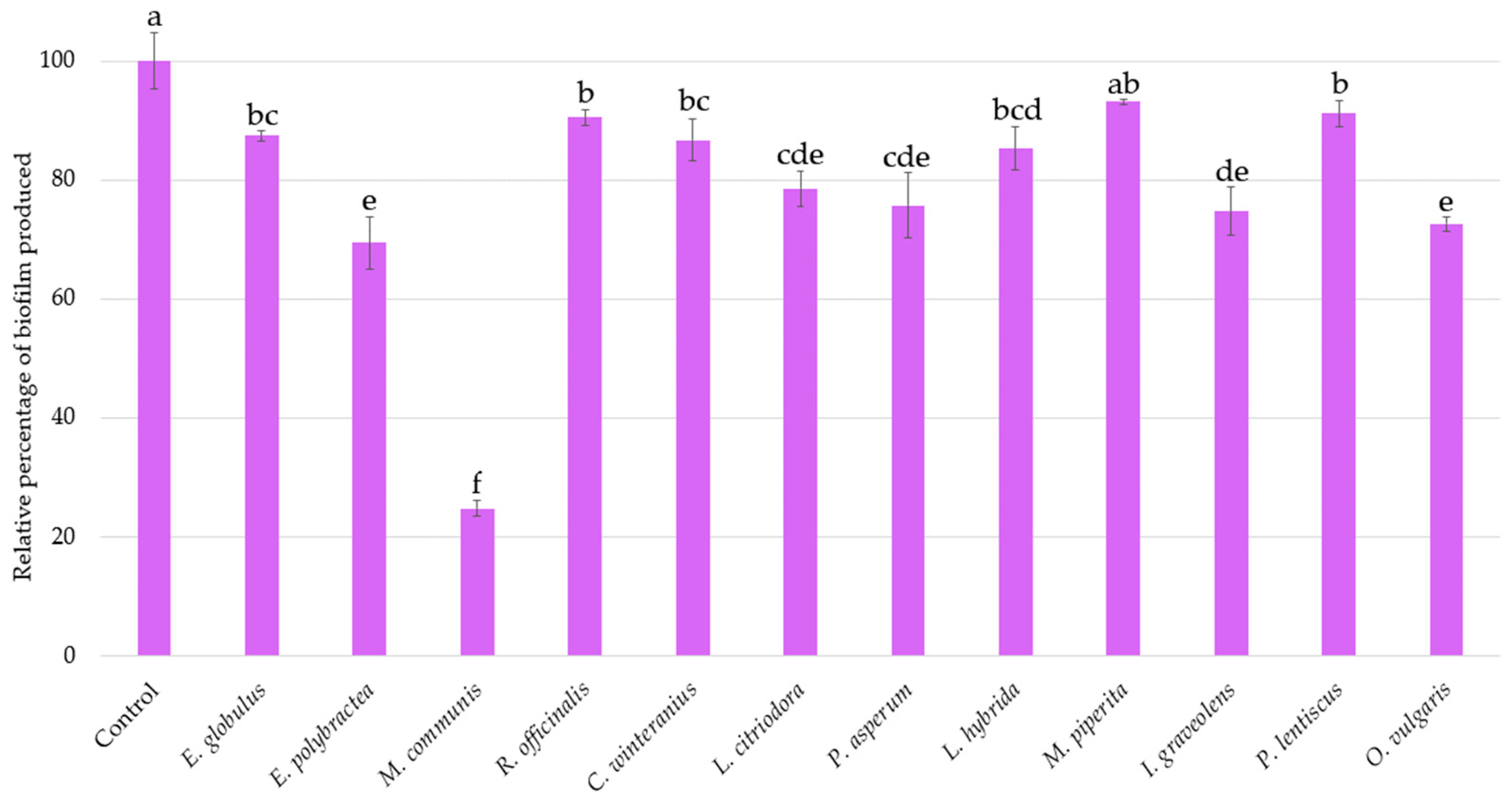
The same inoculum used to assess bioluminescence production was kept for investigating the biofilm production of A. fischeri after a 2 h 30 min treatment with each EO (Figure 3). An inhibition of 25% in biofilm formation is considered significant.
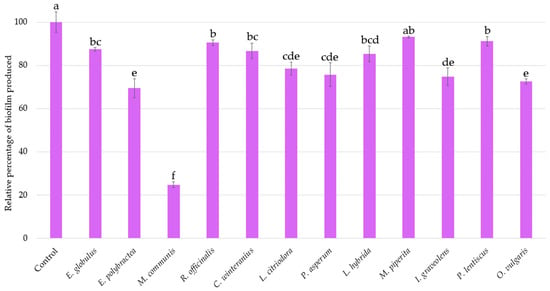
Figure 3.
Relative percentages of Aliivibrio fischeri biofilm production in the absence (control) or presence of EOs. Data are mean values (±SD) of at least four independent measurements (n ≥ 4) different lowercase letters indicate significant differences between each EO (p < 0.001).
The internal control O. vulgaris EO reduces biofilm formation by 27% (±1.25%). With similar activities E. polybractea, L. citriodora, P. asperum, and I. graveolens EOs decrease biofilm production by 20 to 30%. However, the effect of M. communis EO is the most important, with a reduction rate of 75.22% (±1.32%), further highlighting its strong biofilm-reducing activity.
3.6. Quorum-Quenching Activity of Myrtus communis EO and Its Chromatographic Fractions
The activities of the four fractions were assessed using the same parameters evaluated for M. communis EO: swimming, luminescence, and biofilm production. To select the anti-QS concentrations, the MIC of each fraction was determined: F1 and F2 were tested at 50 ppm, while F3 and F4 were used at 25 ppm because of their antibacterial activities at 50 ppm.
In comparison with results obtained for the bacterial positive control and M. communis EO, the F1 and F2 fractions―composed of α-pinene (70.4%) and limonene (20.1%) for the former, and 1,8-cineole (80.8%) for the latter—did not exhibit any activity against A. fischeri swimming (Figure 4a), bioluminescence production (Figure 4b), or biofilm formation (Figure 4c).
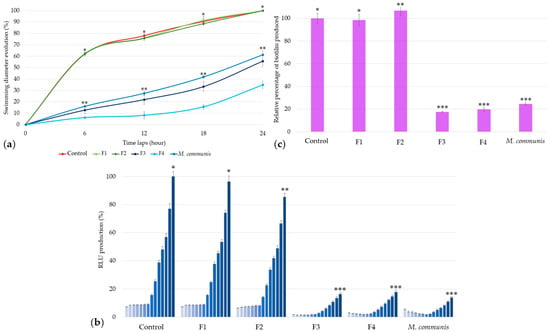
Figure 4.
Activity of M. communis EO and its chromatographic fractions on Aliivibrio fischeri swimming (a), bioluminescence (b), and biofilm production (c). Data are mean values (±SD) of at least four independent measurements (a, n = 3; b and c, n ≥ 4). Different (*), (**) and (***) indicate significant differences between each EO at each time lap (a), at the end of each kinetic cycle (b), or at 2 h 30 min (c) (p < 0.001).
In contrast to M. communis fractions F1 and F2 (G1), the use of 25 ppm of F3 and F4 (G2) against A. fischeri resulted in a significant reduction in swimming motility, respectively, 44.31% (±4.75%) and 65.10% (±3.40%) (Figure 4a).
Similarly, bioluminescence production was reduced by 83.68% (±1.23%) and 82.10% (±1.33%) with F3 and F4 at the end of the kinetic cycle. Both fractions show a P2-type profile, with F3 exhibiting a more intense activity at the beginning of the kinetic cycle (Figure 4b).
The relative quantification of biofilm production highlights the effectiveness of F3 and F4, with reductions of 82.44% (±0.66%) and 80.18% (±1.54%), respectively (Figure 4c).
These results show that M. communis EO, along with its F3 and F4 extracts, exhibits the strongest activity against Aliivibrio fischeri QS, including significant effects on swimming motility, bioluminescence, and biofilm production.
4. Discussion
In contrast to data from the literature, chemotype carvacrol Origanum vulgaris EO showed significant activity only in disrupting bioluminescence with no notable effect on biofilm formation or swimming [39,40,41]. Conversely, Corsican Myrtus communis EO decreased both swimming diameter and the production of bioluminescence as well as biofilm formation. The chemical analysis of M. communis EO yielded an identification rate of 97.3%. Among the 41 identified compounds, α-pinene, 1,8-cineole, and limonene were classified as major constituents. However, these main compounds are also present in other essential oils, such as E. globulus, L. citriodora, R. officinalis, and P. lentiscus, which do not show notable activity against quorum-sensing [42]. To further explore these findings and possibly identify the active molecules, M. communis EO was separated into four distinct chromatographic fractions (F1–F4).
Contrary to initial hypotheses, the overall activities of α-pinene, 1,8-cineole, and limonene were not confirmed. The results presented in Figure 4 show that only two fractions of chromatography of M. communis EO, F3, and F4, exhibited activity, while F1—which contained all the α-pinene and limonene from M. communis EO—did not, contrary to what was previously observed in other Gram-negative quorum-sensing studies [43,44]. Neither F1 nor F2, which contained all the 1,8-cineole, showed any activity, in contrast to other strains [45,46]. In contrast and according to the chemical fraction analysis, only the more polar fractions, F3 and F4, exhibited anti-QS activity comparable to that of the M. communis EO in all three studied parameters.
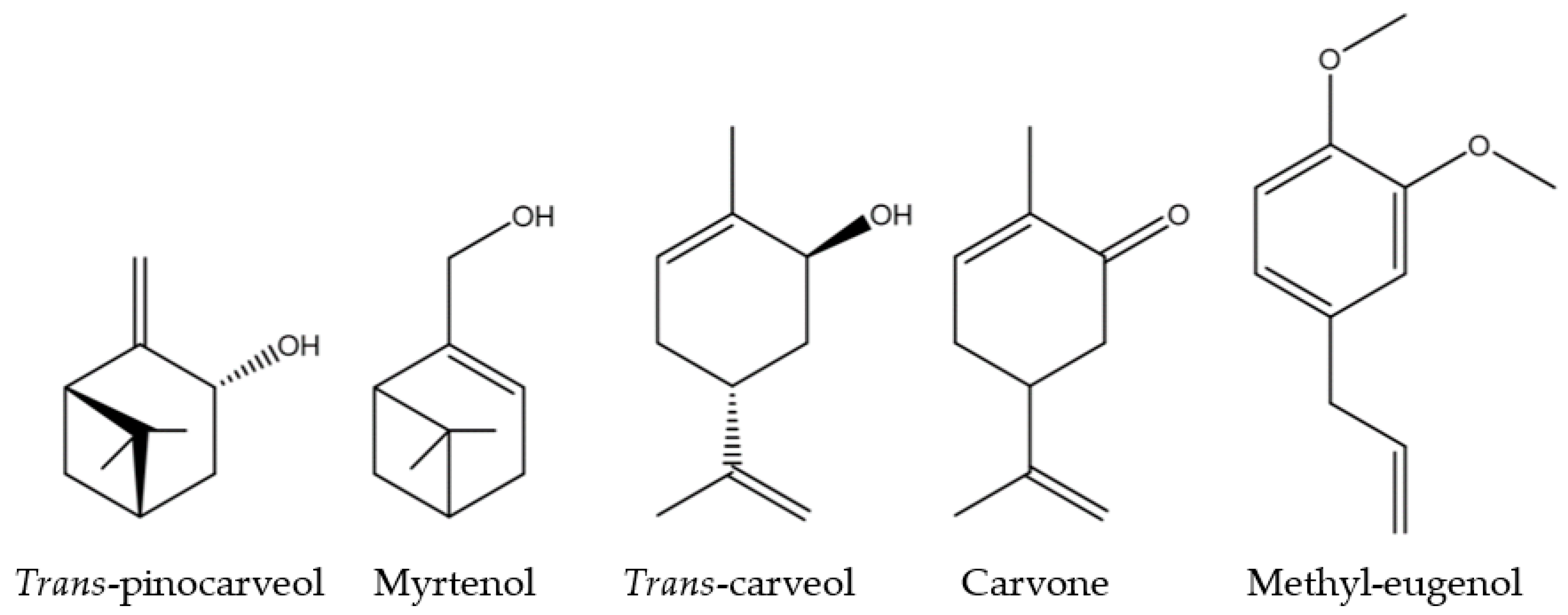
Both F3 and F4 fractions showed similar activity across the three parameters, mirroring the activity of M. communis EO. According to chemical analyses, five molecules, among the identified ones, were found to be common to both F3 and F4: trans-pinocarveol, myrtenol, trans-carveol, carvone, and methyl-eugenol. All of these molecules are oxygenated monoterpenes, except methyl-eugenol (a phenylpropanoid) (Figure 5).
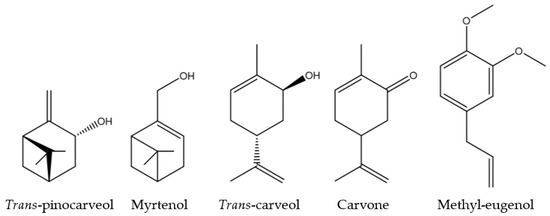
Figure 5.
Chemical structure of the five common F3 and F4 fractions molecules.
If the activities of M. communis EO can be linked to any of these molecules, it could be due to one of them acting alone, a combination of them, or their interaction with others. All the results suggest that a QS mechanism disruption is involved. Based on previous findings, hydroxyl and carbonyl functional groups seem to be important, as well as the presence of double bonds in cyclic molecules [47,48,49]. Trans-pinocarveol and myrtenol are unsaturated bicyclic monoterpenes with a hydroxyl group. Trans-carveol and carvone are unsaturated oxygenated monoterpenes with the p-menthane skeleton. They have two double bonds (C=C, one cyclic) and are differentiated by a hydroxyl group for trans-carveol, replaced by a carbonyl group for carvone. Finally, methyl-eugenol includes an aromatic ring (conjugated planar ring system) substituted by two methoxy groups and an allyl group.
Myrtenol has already demonstrated biofilm reduction ability against Staphylococcus aureus [50] and Klebsiella pneumoniae associated with antibiotics [51]. Concerning carveol, a study on limonene isomers highlighted that carveol exhibits stronger activity against Xanthomonas oryzae compared to limonene [52]. Carveol is a limonene derivative with a hydroxyl group on the C2, which is why the presence of the hydroxyl group is linked to its activity. Another structurally similar molecule to carveol is carvone, which has demonstrated QS disruption in Chromobacterium violaceum and Hafnia alvei, with a reduction in biofilm formation. Additionally, carvone impairs the motility of these two bacteria [53,54]. Indeed, carvone can reduce acyl-homoserine lactone production by disrupting AHL synthase and the QS transcriptional regulator.
As for methyl-eugenol, several studies have demonstrated its QS disruption abilities in Chromobacterium violaceum and Vibrio harveyi. This molecule may act as a competitive ligand for CviR, the QS system in Chromobacterium violaceum [55,56].
In Vibrio harveyi, a reduction in luminescence production has been observed with methyl-eugenol. Among the Gram-negative bacteria mentioned above, the QS systems of Chromobacterium violaceum and Halfnia alvei are regulated by N-hexanoyl-L-homoserine lactone (C6-HSL), which has a structure similar to N-(3-oxohexanoyl)-L-homoserine lactone (3-oxo-C6-HSL), the QS autoinducer for Vibrio harveyi and Aliivibrio fischeri [57]. This autoinducer plays a crucial role in bioluminescence production to the LuxR receptor, which initiates transcription. A reduction in 3-oxo-C6-HSL levels could be due to decreased production or inactivation via binding interactions which would directly disrupt biofilm formation and bioluminescence production in Aliivibrio fischeri. If A. fischeri bioluminescence is majority driven by LuxI/R, motility is regulated by AinS/R and LuxS/PQ systems [58,59]. This second AinS/R system is regulated by N-octanoyl-homoserine lactone (C8-HSL) also implied in bioluminescence at low cellular density [60].
Therefore, some of these five molecules, either alone or in combination, may interact with 3-oxo-C6-HSL and/or C8-HSL, potentially affecting other bacteria as well. For instance, QS systems of other Vibrio species, such as that of Vibrio anguillarum, a fish pathogen that can also infect humans, might be impacted [61]. The QS of Yersinia enterocolitica, a human pathogen, can also be disrupted because its most produced autoinducer is 3-oxo-C6-HSL [62]. Additionally, Agrobacterium tumefaciens and Erwinia carotovora, two plant pathogens, could experience reduced tumor production and the diminished activity of enzymes responsible for cell wall degradation due to the inhibition of C8-HSL or 3-oxo-C6-HSL [63,64].
5. Conclusions
To conclude, Myrtus communis Corsican essential oil is the most active among the twelve essential oils tested in the study. This EO was fractioned by column chromatography and the F3 and F4 oxygenated fractions demonstrated strong quorum-quenching activity at a sub-inhibitory concentration by disrupting biofilm formation, mobility, and bioluminescence. The EO of M. communis and its F3 and F4 fractions showed stronger activity than the carvacrol-rich O. vulgaris EO. Molecules, such as trans-pinocarveol, myrtenol, trans-carveol, carvone, and methyl eugenol, were identified in both active fractions only. These results highlight the importance of minority compounds which can interfere with QS-related traits. An in-depth study is necessary to elucidate their mechanisms of action and to consider the application of active fractions or compounds against other marine biofilm-forming bacteria with environmental and clinical relevance.
Author Contributions
Conceptualization, E.H., A.B., L.B., and E.G.; methodology E.H., A.B., M.P., and E.G.; validation, J.-P.P., A.B., M.P., L.B., and E.G.; formal analysis, E.H., J.-P.P., A.B., M.P., T.M., L.B., and E.G.; investigation, E.H., M.P., and T.M.; data curation, E.H., J.-P.P., A.B., M.P., T.M., L.B., and E.G.; writing—original draft preparation, E.H.; writing—review and editing, E.H., J.-P.P., A.B., M.P., T.M., L.B., and E.G.; visualization, E.H., J.-P.P., A.B., M.P., T.M., L.B., and E.G.; supervision, J.-P.P., A.B., L.B., and E.G.; project administration, L.B. and E.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article. Further inquiries can be directed to the corresponding author.
Acknowledgments
The authors are grateful to the Collectivité de Corse (E. Hardy doctoral contract). The authors gratefully acknowledge Philippe Biagini for his valuable discussions about this paper and Radia Lourkisti for her help with the statistical analyses.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Vinoj, G.; Vaseeharan, B.; Thomas, S.; Spiers, A.J.; Shanthi, S. Quorum-Quenching Activity of the AHL-Lactonase from Bacillus Licheniformis DAHB1 Inhibits Vibrio Biofilm Formation In Vitro and Reduces Shrimp Intestinal Colonisation and Mortality. Mar. Biotechnol. 2014, 16, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Defoirdt, T. Quorum-Sensing Systems as Targets for Antivirulence Therapy. Trends Microbiol. 2018, 26, 313–328. [Google Scholar] [CrossRef] [PubMed]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum Sensing in Bacteria: The LuxR-LuxI Family of Cell Density-Responsive Transcriptional Regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar] [CrossRef] [PubMed]
- Nealson, K.H.; Platt, T.; Hastings, J.W. Cellular Control of the Synthesis and Activity of the Bacterial Luminescent System. J. Bacteriol. 1970, 104, 313–322. [Google Scholar] [CrossRef]
- Spacapan, M.; Bez, C.; Venturi, V. Quorum Sensing Going Wild. iScience 2023, 26, 108000. [Google Scholar] [CrossRef]
- Fung, B.L.; Visick, K.L. LitR and Its Quorum-Sensing Regulators Modulate Biofilm Formation by Vibrio fischeri. J. Bacteriol. 2025, 207, e00476-24. [Google Scholar] [CrossRef]
- Septer, A.N.; Visick, K.L. Lighting the Way: How the Vibrio fischeri Model Microbe Reveals the Complexity of Earth’s “Simplest” Life Forms. J. Bacteriol. 2024, 206, e0003524. [Google Scholar] [CrossRef]
- Fung, B.L.; Esin, J.J.; Visick, K.L. Vibrio fischeri: A Model for Host-Associated Biofilm Formation. J. Bacteriol. 2024, 206, e00370-23. [Google Scholar] [CrossRef]
- Frans, I.; Michiels, C.W.; Bossier, P.; Willems, K.A.; Lievens, B.; Rediers, H. Vibrio anguillarum as a Fish Pathogen: Virulence Factors, Diagnosis and Prevention. J. Fish Dis. 2011, 34, 643–661. [Google Scholar] [CrossRef]
- Lorenzoni, G.; Tedde, G.; Mara, L.; Bazzoni, A.M.; Esposito, G.; Salza, S.; Piras, G.; Tedde, T.; Bazzardi, R.; Arras, I.; et al. Presence, Seasonal Distribution, and Biomolecular Characterization of Vibrio parahaemolyticus and Vibrio vulnificus in Shellfish Harvested and Marketed in Sardinia (Italy) between 2017 and 2018. J. Food Prot. 2021, 84, 1549–1554. [Google Scholar] [CrossRef]
- Leighton, R.E.; Correa Vélez, K.E.; Xiong, L.; Creech, A.G.; Amirichetty, K.P.; Anderson, G.K.; Cai, G.; Norman, R.S.; Decho, A.W. Vibrio parahaemolyticus and Vibrio vulnificus In Vitro Colonization on Plastics Influenced by Temperature and Strain Variability. Front. Microbiol. 2023, 13, 1099502. [Google Scholar] [CrossRef] [PubMed]
- Kovács, J.K.; Felső, P.; Makszin, L.; Pápai, Z.; Horváth, G.; Ábrahám, H.; Palkovics, T.; Böszörményi, A.; Emődy, L.; Schneider, G. Antimicrobial and Virulence-Modulating Effects of Clove Essential Oil on the Foodborne Pathogen Campylobacter jejuni. Appl. Environ. Microbiol. 2016, 82, 6158–6166. [Google Scholar] [CrossRef]
- Poli, J.-P.; Guinoiseau, E.; Luciani, A.; Yang, Y.; Battesti, M.-J.; Paolini, J.; Costa, J.; Quilichini, Y.; Berti, L.; Lorenzi, V. Key Role of Hydrogen Peroxide in Antimicrobial Activity of Spring, Honeydew maquis and Chestnut Grove Corsican Honeys on Pseudomonas aeruginosa DNA. Lett. Appl. Microbiol. 2018, 66, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Burt, S. Essential Oils: Their Antibacterial Properties and Potential Applications in Foods—A Review. Int. J. Food Microbiol. 2004, 94, 223–253. [Google Scholar] [CrossRef] [PubMed]
- Bavarsad, N.; Kouchak, M.; Mohamadipour, P.; Sadeghi-Nejad, B. Preparation and Physicochemical Characterization of Topical Chitosan-Based Film Containing Griseofulvin-Loaded Liposomes. J. Adv. Pharm. Technol. Res. 2016, 7, 91. [Google Scholar] [CrossRef] [PubMed]
- Basavegowda, N.; Baek, K.-H. Synergistic Antioxidant and Antibacterial Advantages of Essential Oils for Food Packaging Applications. Biomolecules 2021, 11, 1267. [Google Scholar] [CrossRef]
- Bouzabata, A.; Castola, V.; Bighelli, A.; Abed, L.; Casanova, J.; Tomi, F. Chemical Variability of Algerian Myrtus communis L. Chem. Biodivers. 2013, 10, 129–137. [Google Scholar] [CrossRef]
- Barbosa, L.; Filomeno, C.; Teixeira, R. Chemical Variability and Biological Activities of Eucalyptus Spp. Essential Oils. Molecules 2016, 21, 1671. [Google Scholar] [CrossRef]
- Da Cruz, E.D.N.S.; Peixoto, L.D.S.; Da Costa, J.S.; Mourão, R.H.V.; Do Nascimento, W.M.O.; Maia, J.G.S.; Setzer, W.N.; Da Silva, J.K.; Figueiredo, P.L.B. Seasonal Variability of a Caryophyllane Chemotype Essential Oil of Eugenia patrisii Vahl Occurring in the Brazilian Amazon. Molecules 2022, 27, 2417. [Google Scholar] [CrossRef]
- De Sousa, D.P.; Damasceno, R.O.S.; Amorati, R.; Elshabrawy, H.A.; De Castro, R.D.; Bezerra, D.P.; Nunes, V.R.V.; Gomes, R.C.; Lima, T.C. Essential Oils: Chemistry and Pharmacological Activities. Biomolecules 2023, 13, 1144. [Google Scholar] [CrossRef]
- Couic-Marinier, F.; Lobstein, A. Composition chimique des huiles essentielles. Actual. Pharm. 2013, 52, 22–25. [Google Scholar] [CrossRef]
- Wijesundara, N.M.; Lee, S.F.; Cheng, Z.; Davidson, R.; Langelaan, D.N.; Rupasinghe, H.P.V. Bactericidal Activity of Carvacrol against Streptococcus pyogenes Involves Alteration of Membrane Fluidity and Integrity through Interaction with Membrane Phospholipids. Pharmaceutics 2022, 14, 1992. [Google Scholar] [CrossRef] [PubMed]
- Herman, A.; Tambor, K.; Herman, A. Linalool Affects the Antimicrobial Efficacy of Essential Oils. Curr. Microbiol. 2016, 72, 165–172. [Google Scholar] [CrossRef]
- Wagle, B.R.; Upadhyay, A.; Upadhyaya, I.; Shrestha, S.; Arsi, K.; Liyanage, R.; Venkitanarayanan, K.; Donoghue, D.J.; Donoghue, A.M. Trans-Cinnamaldehyde, Eugenol and Carvacrol Reduce Campylobacter jejuni Biofilms and Modulate Expression of Select Genes and Proteins. Front. Microbiol. 2019, 10, 1837. [Google Scholar] [CrossRef]
- Lambert, R.J.W.; Skandamis, P.N.; Coote, P.J.; Nychas, G.-J.E. A Study of the Minimum Inhibitory Concentration and Mode of Action of Oregano Essential Oil, Thymol and Carvacrol. J. Appl. Microbiol. 2001, 91, 453–462. [Google Scholar] [CrossRef]
- Asfour, H. Anti-Quorum Sensing Natural Compounds. J. Microsc. Ultrastruct. 2018, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Aljaafari, M.N.; AlAli, A.O.; Baqais, L.; Alqubaisy, M.; AlAli, M.; Molouki, A.; Ong-Abdullah, J.; Abushelaibi, A.; Lai, K.-S.; Lim, S.-H.E. An Overview of the Potential Therapeutic Applications of Essential Oils. Molecules 2021, 26, 628. [Google Scholar] [CrossRef]
- Rattanachaikunsopon, P.; Phumkhachorn, P. Assessment of Factors Influencing Antimicrobial Activity of Carvacrol and Cymene against Vibrio cholerae in Food. J. Biosci. Bioeng. 2010, 110, 614–619. [Google Scholar] [CrossRef]
- Guinoiseau, E.; Luciani, A.; Rossi, P.G.; Quilichini, Y.; Ternengo, S.; Bradesi, P.; Berti, L. Cellular Effects Induced by Inula graveolens and Santolina corsica Essential Oils on Staphylococcus aureus. Eur. J. Clin. Microbiol. Infect. Dis. 2010, 29, 873–879. [Google Scholar] [CrossRef]
- Guinoiseau, E.; Lorenzi, V.; Luciani, A.; Tomi, F.; Casanova, J.; Berti, L. Susceptibility of the Multi-Drug Resistant Strain of Enterobacter aerogenes EA289 to the Terpene Alcohols from Cistus ladaniferus Essential Oil. Nat. Prod. Commun. 2011, 6, 1159–1162. [Google Scholar] [CrossRef]
- Poli, J.-P.; Guinoiseau, E.; De Rocca Serra, D.; Sutour, S.; Paoli, M.; Tomi, F.; Quilichini, Y.; Berti, L.; Lorenzi, V. Anti-Quorum Sensing Activity of 12 Essential Oils on Chromobacterium violaceum and Specific Action of Cis-Cis-p-Menthenolide from Corsican Mentha suaveolens ssp. Insularis. Molecules 2018, 23, 2125. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.-X.; Zeng, Y.-H.; Chen, H.-M.; Zhang, N.; Han, Y.; Long, H.; Xie, Z.-Y. Bioinformatic and Functional Characterization of Cyclic-Di-GMP Metabolic Proteins in Vibrio Alginolyticus Unveils Key Diguanylate Cyclases Controlling Multiple Biofilm-Associated Phenotypes. Front. Microbiol. 2023, 14, 1258415. [Google Scholar] [CrossRef] [PubMed]
- Miladinović, D.L.; Dimitrijević, M.V.; Mihajilov-Krstev, T.M.; Marković, M.S.; Ćirić, V.M. The Significance of Minor Components on the Antibacterial Activity of Essential Oil via Chemometrics. LWT 2021, 136, 110305. [Google Scholar] [CrossRef]
- Babushok, V.I.; Linstrom, P.J.; Zenkevich, I.G. Retention Indices for Frequently Reported Compounds of Plant Essential Oils. J. Phys. Chem. Ref. Data 2011, 40, 043101. [Google Scholar] [CrossRef]
- Chibane, E.M.; Ouknin, M.; Renucci, F.; Costa, J.; Majidi, L. Chemical Profile, Antioxidant and Antifungal Activity of Essential Oil from Cladanthus eriolepis. J. Essent. Oil Bear. Plants 2020, 23, 1296–1305. [Google Scholar] [CrossRef]
- Afifi, S.M.; El-Mahis, A.; Heiss, A.G.; Farag, M.A. Gas Chromatography–Mass Spectrometry-Based Classification of 12 Fennel (Foeniculum vulgare Miller) Varieties Based on Their Aroma Profiles and Estragole Levels as Analyzed Using Chemometric Tools. ACS Omega 2021, 6, 5775–5785. [Google Scholar] [CrossRef]
- National Institute of Standards and Technology. The NIST/EPA/NIH Mass Spectral Library; PC Version 2.1.2.19; Perkin-Elmer Corporation, NIST, U.S. Department of Commerce: Norwalk, CT, USA, 2014. [Google Scholar]
- Christensen, D.G.; Visick, K.L. Vibrio fischeri: Laboratory Cultivation, Storage, and Common Phenotypic Assays. CP Microbiol. 2020, 57, e103. [Google Scholar] [CrossRef]
- Čabarkapa, I.; Čolović, R.; Đuragić, O.; Popović, S.; Kokić, B.; Milanov, D.; Pezo, L. Anti-Biofilm Activities of Essential Oils Rich in Carvacrol and Thymol against Salmonella Enteritidis. Biofouling 2019, 35, 361–375. [Google Scholar] [CrossRef]
- Datta, S.; Singh, V.; Nag, S.; Roy, D.N. Carvacrol, a Monoterpenoid, Binds Quorum Sensing Proteins (LasI and LasR) and Swarming Motility Protein BswR of Pseudomonas aeruginosa, Resulting in Loss of Pathogenicity: An In Silico Approach. Can. J. Microbiol. 2025, 71, 1–15. [Google Scholar] [CrossRef]
- Lu, L.; Wang, J.; Qin, T.; Chen, K.; Xie, J.; Xi, B. Carvacrol Inhibits Quorum Sensing in Opportunistic Bacterium Aeromonas hydrophila. Microorganisms 2023, 11, 2027. [Google Scholar] [CrossRef]
- Abbas, A.; Anwar, F.; Alqahtani, S.M.; Ahmad, N.; Al-Mijalli, S.H.; Shahid, M.; Iqbal, M. Hydro-Distilled and Supercritical Fluid Extraction of Eucalyptus camaldulensis Essential Oil: Characterization of Bioactives Along with Antioxidant, Antimicrobial and Antibiofilm Activities. Dose-Response 2022, 20, 15593258221125477. [Google Scholar] [CrossRef] [PubMed]
- Qaralleh, H. Limonene as a Multi-Target Antibiofilm and Quorum Sensing Inhibitor Against Pseudomonas Aeruginosa. J. Basic Appl. Res. Biomed. 2024, 10, 80–88. [Google Scholar] [CrossRef]
- Wei, S.-Y.; Li, Y.-L.; Wang, L.; Chu, Z.-Y.; Qin, Y.-C.; Zeng, H. α-Pinene: Inhibitor of Acinetobacter Baumannii Biofilms and Potential Therapeutic Agent for Pneumonia. Int. Immunopharmacol. 2025, 151, 114287. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Baindara, P.; Sharma, P.; Jose, T.A.; V, K.; Manoharan, R.; Mandal, S.M. Anti-Biofilm Action of Cineole and Hypericum perforatum to Combat Pneumonia-Causing Drug-Resistant P. aeruginosa. Antibiotics 2024, 13, 689. [Google Scholar] [CrossRef]
- Karuppiah, V.; Thirunanasambandham, R.; Thangaraj, G. Anti-Quorum Sensing and Antibiofilm Potential of 1,8-Cineole Derived from Musa paradisiaca against Pseudomonas aeruginosa Strain PAO1. World J. Microbiol. Biotechnol. 2021, 37, 66. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef]
- Khwaza, V.; Aderibigbe, B.A. Antibacterial Activity of Selected Essential Oil Components and Their Derivatives: A Review. Antibiotics 2025, 14, 68. [Google Scholar] [CrossRef]
- Ultee, A.; Bennik, M.H.J.; Moezelaar, R. The Phenolic Hydroxyl Group of Carvacrol Is Essential for Action against the Food-Borne Pathogen Bacillus cereus. Appl. Environ. Microbiol. 2002, 68, 1561–1568. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, A.; Jayasree, T.; Valliammai, A.; Pandian, S.K. Myrtenol Attenuates MRSA Biofilm and Virulence by Suppressing sarA Expression Dynamism. Front. Microbiol. 2019, 10, 2027. [Google Scholar] [CrossRef]
- Maione, A.; La Pietra, A.; De Alteriis, E.; Mileo, A.; De Falco, M.; Guida, M.; Galdiero, E. Effect of Myrtenol and Its Synergistic Interactions with Antimicrobial Drugs in the Inhibition of Single and Mixed Biofilms of Candida Auris and Klebsiella Pneumoniae. Microorganisms 2022, 10, 1773. [Google Scholar] [CrossRef]
- Kim, H.; Kim, M.H.; Choi, U.-L.; Chung, M.-S.; Yun, C.-H.; Shim, Y.; Oh, J.; Lee, S.; Lee, G.W. Molecular and Phenotypic Investigation on Antibacterial Activities of Limonene Isomers and Its Oxidation Derivative against Xanthomonas oryzae Pv. Oryzae. J. Microbiol. Biotechnol. 2024, 34, 562–569. [Google Scholar] [CrossRef] [PubMed]
- Kanekar, S.; Fathima, F.; Rekha, P.-D. Carvone—A Quorum Sensing Inhibitor Blocks Biofilm Formation in Chromobacterium violaceum. Nat. Prod. Res. 2022, 36, 4540–4545. [Google Scholar] [CrossRef]
- Li, T.; Mei, Y.; He, B.; Sun, X.; Li, J. Reducing Quorum Sensing-Mediated Virulence Factor Expression and Biofilm Formation in Hafnia alvei by Using the Potential Quorum Sensing Inhibitor L-Carvone. Front. Microbiol. 2019, 9, 3324. [Google Scholar] [CrossRef] [PubMed]
- Sybiya Vasantha Packiavathy, I.A.; Agilandeswari, P.; Musthafa, K.S.; Karutha Pandian, S.; Veera Ravi, A. Antibiofilm and Quorum Sensing Inhibitory Potential of Cuminum cyminum and Its Secondary Metabolite Methyl Eugenol against Gram Negative Bacterial Pathogens. Food Res. Int. 2012, 45, 85–92. [Google Scholar] [CrossRef]
- Wang, W.; Lin, X.; Yang, H.; Huang, X.; Pan, L.; Wu, S.; Yang, C.; Zhang, L.; Li, Y. Anti-Quorum Sensing Evaluation of Methyleugenol, the Principal Bioactive Component, from the Melaleuca bracteata Leaf Oil. Front. Microbiol. 2022, 13, 970520. [Google Scholar] [CrossRef]
- Milton, D.L. Quorum Sensing in Vibrios: Complexity for Diversification. Int. J. Med. Microbiol. 2006, 296, 61–71. [Google Scholar] [CrossRef]
- Lupp, C.; Urbanowski, M.; Greenberg, E.P.; Ruby, E.G. The Vibrio fischeri Quorum-sensing Systems Ain and Lux. Sequentially Induce Luminescence Gene Expression and Are Important for Persistence in the Squid Host. Mol. Microbiol. 2003, 50, 319–331. [Google Scholar] [CrossRef]
- Dial, C.N.; Eichinger, S.J.; Foxall, R.; Corcoran, C.J.; Tischler, A.H.; Bolz, R.M.; Whistler, C.A.; Visick, K.L. Quorum Sensing and Cyclic Di-GMP Exert Control Over Motility of Vibrio fischeri KB2B1. Front. Microbiol. 2021, 12, 690459. [Google Scholar] [CrossRef] [PubMed]
- Lupp, C.; Ruby, E.G. Vibrio fischeri Uses Two Quorum-Sensing Systems for the Regulation of Early and Late Colonization Factors. J. Bacteriol. 2005, 187, 3620–3629. [Google Scholar] [CrossRef]
- Hu, J.-Y.; Zhang, X.-K.; Xin, C.-Q.; Zhang, L.; Kang, J.; Gong, P. Severe Infection by Vibrio anguillarum Following a Bite by a Marine Fish: A Case Report. Emerg. Microbes Infect. 2023, 12, 2204145. [Google Scholar] [CrossRef]
- Ng, Y.-K.; Grasso, M.; Wright, V.; Garcia, V.; Williams, P.; Atkinson, S. The Quorum Sensing System of Yersinia enterocolitica 8081 Regulates Swimming Motility, Host Cell Attachment, and Virulence Plasmid Maintenance. Genes 2018, 9, 307. [Google Scholar] [CrossRef] [PubMed]
- Kumar, L.; Patel, S.K.S.; Kharga, K.; Kumar, R.; Kumar, P.; Pandohee, J.; Kulshresha, S.; Harjai, K.; Chhibber, S. Molecular Mechanisms and Applications of N-Acyl Homoserine Lactone-Mediated Quorum Sensing in Bacteria. Molecules 2022, 27, 7584. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, H.; Xie, Y.; Guo, Y.; Cheng, Y.; Yao, W. Inhibitory Effects of Hexanal on Acylated Homoserine Lactones (AHLs) Production to Disrupt Biofilm Formation and Enzymes Activity in Erwinia carotovora and Pseudomonas fluorescens. J. Food Sci. Technol. 2023, 60, 372–381. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).